
Abstract
Direct-acting antivirals (DAAs) not only rapidly inhibited hepatitis C virus (HCV) replication but also modulated innate and adaptive immune response in chronic hepatitis C patients. However, the regulatory activity of DAAs to Toll-like receptor 2 (TLR2) stimulation on CD4+CD25+CD127dim/− regulatory T cells (Tregs) and T helper (Th) 17 cells was not completely understood. In the present study, a total of 23 patients with chronic HCV genotype 1b infection were enrolled, and blood samples were collected at baseline (treatment naive), end of therapy (EOT), and 12 weeks after EOT (SVR12) with daclatasvir plus asunaprevir therapy. TLR2 expression on Tregs and Th17 cells was measured by flow cytometry. Cellular proliferation, cytokine production, and suppressive activity were also tested in purified CD4+CD25+CD127dim/− Tregs in response to the stimulation of Pam3Csk4, an agonist of TLR2. Inhibition of HCV RNA by daclatasvir and asunaprevir did not affect either percentage of Tregs/Th17 cells or TLR2 expression on Tregs/Th17 cells. Pam3Csk4 stimulation also did not influence either cellular proliferation or Tregs/Th17 proportion at each time point. Stimulation with Pam3Csk4 only enhanced the suppressive function and interleukin (IL)-35 production by Tregs purified from baseline, but not those from EOT or SVR12. Similarly, Pam3Csk4 stimulation only elevated Th17 cell frequency of CD4+ T cells from baseline, but not those from EOT or SVR12. Moreover, daclatasvir and asunaprevir therapy did not promote TLR2-induced shift of Tregs toward Th17-like phenotype and function. These data suggested that daclatasvir plus asunaprevir therapy resulted in the decreased responsiveness of Tregs/Th17 cells to TLR2 stimulation in chronic hepatitis C patients, which might provide a novel mechanism underlying DAA-induced immunoregulation.
Introduction
H
Immune activation and T cell energy/exhaustion are strongly associated with disease progression and pathogenesis of HCV infection (20). However, few studies focused on the DAA therapy on the immune response in chronic hepatitis C patients. Burchill et al. demonstrated that DAAs induced rapid reversal of innate immune dysregulation with decline in genes associated with chronic interferon (IFN) stimulation and inflammation (3). Moreover, Najafi Fard et al. showed that DAA therapy enhanced total CD4+ and CD8+ T cell responses in chronic hepatitis C patients (17). These results suggested that IFN-free DAA regimen treatment could affect both innate and adaptive immune response in chronic hepatitis C patients.
Toll-like receptors (TLRs) are classical molecules in innate immunity and play critical roles in early host defense to cancers and viral infections (10). Signaling through TLR induces the expression of cytokines and chemokines, leading to the activation of adaptive immunity (14,18,25). TLR2 direct modulated CD4+CD25+ Tregs activity and promoted Th17 response in chronic hepatitis B patients (28,29). Our previous study also revealed that HCV infection shifted the balance of CD4+CD25+CD127dim/− regulatory T cells (Tregs) and T helper (Th) 17 cells through TLR2 stimulation, which provided a link between innate and adaptive immune response in HCV infection (13). Due to the immunoregulatory activity of DAAs, we hypothesized that inhibition of HCV replication by DAA treatment also alters TLR2 stimulation on Tregs/Th17 cell balance and functions. To test this possibility, we investigated the effect of TLR2 stimulation on inhibitory function of Tregs and Th17 differentiation in chronic hepatitis C patients with daclatasvir plus asunaprevir therapy.
Patients, Materials, and Methods
Subjects
The study protocol was approved by the Ethics Committee of the Affiliated Hospital to Changchun University of Chinese Medicine, and written informed consent was obtained from each subject. A total of 23 patients [14 males and 9 females; age, 37 (18–61) years; HCV RNA, 5.82 (2.21–8.01) log10copies/mL] with chronic HCV genotype 1b infection were enrolled in this study. All patients were followed up or hospitalized in the Affiliated Hospital to Changchun University of Chinese Medicine from May 2017 to June 2017. No patients were coinfected with human immunodeficiency virus (HIV) or other hepatitis viruses. The subjects who were afflicted with autoimmune disorder or cancer and were previously treated with interferon or other immunomodulatory reagents were also excluded. All patients were diagnosed exclusion of decompensated liver cirrhosis by ultrasound, Fibroscan test, or abdominal CT scan. All patients were treated with daclatasvir (60 mg, once daily) plus asunaprevir (100 mg, twice daily) for 24 weeks. Peripheral blood samples were collected at baseline, end of therapy (EOT), and 12 weeks post-therapy.
Peripheral blood mononuclear cell isolation
Peripheral blood mononuclear cells (PBMCs) were isolated by Ficoll-Hypaque (Sigma-Aldrich, St. Louis, MO) density gradient centrifugation. The isolated PBMCs were checked for viability and were frozen in liquid nitrogen at 107/mL until usage.
CD4+ T cells and CD4+CD25+CD127dim/− Tregs purification
CD4+ T cells and CD4+CD25+CD127dim/− Tregs were purified using Human CD4+ Cell Isolation Kit (Miltenyi, Bergisch Gladbach, Germany) and CD4+CD25+CD127dim/− Cell Isolation Kit II (Miltenyi) following the instruction of the manufacturer. The purity of enriched cells was more than 95% based on flow cytometry determination. Cells were cultured in RPMI1640 supplemented with 10% of heat-inactivated fetal bovine serum (FBS) at 37°C under a 5% CO2 environment.
Cell culture
A measure of 2.5 × 104 of purified CD4+CD25+CD127dim/− Tregs was cocultured with 105 of autologous CD4+CD25− T cells with anti-CD3/CD28 (eBioscience, San Diego, CA; final concentration 1 μg/mL) for 48 h. Before mixture, Tregs were stimulated with Pam3Csk4 (InvivoGen, San Diego, CA; final concentration 3 μg/mL) for 24 h. In certain experiments, PBMCs, CD4+CD25+CD127dim/− Tregs, or CD4+ T cells were stimulated with Pam3Csk4 (InvivoGen, final concentration 3 μg/mL) for 48 or 96 h. Cells and supernatants were harvested for further studies.
Flow cytometry
Cells were stimulated with Phorbol-12-myristate-13-acetate (50 ng/mL) and ionomycin (1 μg/mL), supplemented with Brefeldin A (10 μg/mL) for 6 h for Th17 cell measurement. Cells were transferred into FACS tubes, and anti-CD3-PerCP (BD Biosciences, San Jose, CA), anti-CD4-APC (eBioscience), anti-CD25-PE Cy7 (eBioscience), and anti-TLR2-PE (eBioscience) were added for a 30 min incubation at 4°C. Cells were then fixed by 100 μL of Fixation & Permeabilization Medium A (Caltag Laboratories, Invitrogen, Carlsbad, CA) for a 15 min incubation and then were resuspended in 100 μL of Fixation & Permeabilization Medium B (Caltag Laboratories, Invitrogen) containing anti-interleukin (IL)-17A-FITC (eBioscience) for a 20 min incubation at room temperature. In certain experiments, cells were only stained with anti-CCR4-PerCP (BD Biosciences) and anti-CCR6-PE (BD Biosciences). Samples were analyzed with a BD FACSAria II analyzer (BD Biosciences). Acquisitions were performed with CellQuest Pro software (BD Biosciences), and analyses were performed with FlowJo version 8.7.2 for Windows (Tree Star, Ashland, OR).
Cellular proliferation assay
Cellular proliferation was measured using Cell Counting Kit-8 (CCK-8, Alexis Biochemicals, San Diego, CA) in 96-well plates following the instruction of the manufacturer as previously described (13). Briefly, 20 μL of CCK-8 solution was added to each well for the last 4 h culture. Wells contained known number of viable PBMCs for creating a calibration curve to calculate tested cell numbers. The absorbance of samples was measured using Model 680 Microplate Reader (Bio-Rad, Hercules, CA) at 450 nm.
Enzyme-linked immunosorbent assay
Concentrations of IL-10, IL-35, IL-17, and IL-22 were measured using commercial Enzyme-linked Immunosorbent Assay Kits (CUSABIO, Wuhan, Hubei Province, China) following the instruction of the manufacturer.
Statistical analyses
All data were analyzed using SPSS19.0 for Windows (SPSS, Chicago, IL). Shapiro–Wilk test was used for normal distribution assay. Variables following normal distribution are presented as mean ± standard deviation, and statistical significance was determined by one-way ANOVA, SNK-q test, or paired t test. Variables following skewed distribution were presented as median [Q1, Q3], and statistical significance was determined by Kruskal–Wallis test or Wilcoxon matched pairs test. All tests were two tailed, and p < 0.05 was considered to indicate a significant difference.
Results
Virological and biochemical response to daclatasvir plus asunaprevir therapy
All of the 23 chronic hepatitis C patients completed daclatasvir plus asunaprevir therapy for 24 weeks, and no severe side effects were observed during and post therapy. All patients achieved virological response with undetectable HCV RNA at the EOT and achieved SVR at 12 weeks after EOT (SVR12). Nineteen of 23 patients were afflicted with elevated alanine aminotransferase (ALT) at baseline, whereas 17 patients achieved biochemical response at EOT and SVR12 with ALT normalization.
Comparable proportion of Tregs/Th17 cells and TLR2 expression in response to daclatasvir plus asunaprevir therapy
Peripheral bloods were collected at baseline, EOT, and SVR12. PBMCs were isolated for analysis of Tregs/Th17 cells and TLR2 expression using flow cytometry. Representative flow dots of CD4+CD25+CD127dim/− Tregs and IL-17A-producing CD4+ T cells (Th17 cells) are shown in Figure 1A and B, respectively. The level of TLR2 expression was also measured in both Tregs and Th17 cells, and typical flow histogram is also shown in Figure 1A and B, respectively. Daclatasvir plus asunaprevir therapy did not lead to the changes in proportion of either Tregs (Baseline: 4.90% ± 1.40%; EOT: 4.56% ± 0.97%; SVR12: 4.91% ± 0.88%; one-way ANOVA, p = 0.486; Fig. 1C) or Th17 cells (Baseline: 1.41% ± 0.17%; EOT: 1.41% ± 0.17%; SVR12: 1.39% ± 0.16%; one-way ANOVA, p = 0.846; Fig. 1D). There were also no significant differences among mean fluorescence intensity values corresponding to TLR2 in either Tregs (Baseline: 120.1 ± 7.97; EOT: 119.8 ± 6.93; SVR12: 120.5 ± 11.00; one-way ANOVA, p = 0.963; Fig. 1E) or Th17 cells (Baseline: 81.65 ± 11.83; EOT: 79.37 ± 10.89; SVR12: 78.78 ± 10.41; one-way ANOVA, p = 0.651; Fig. 1F). There were no significant differences of either Tregs/Th17 frequency or TLR2 expression between patients with elevated ALT and those with normal ALT at baseline (all p > 0.05).
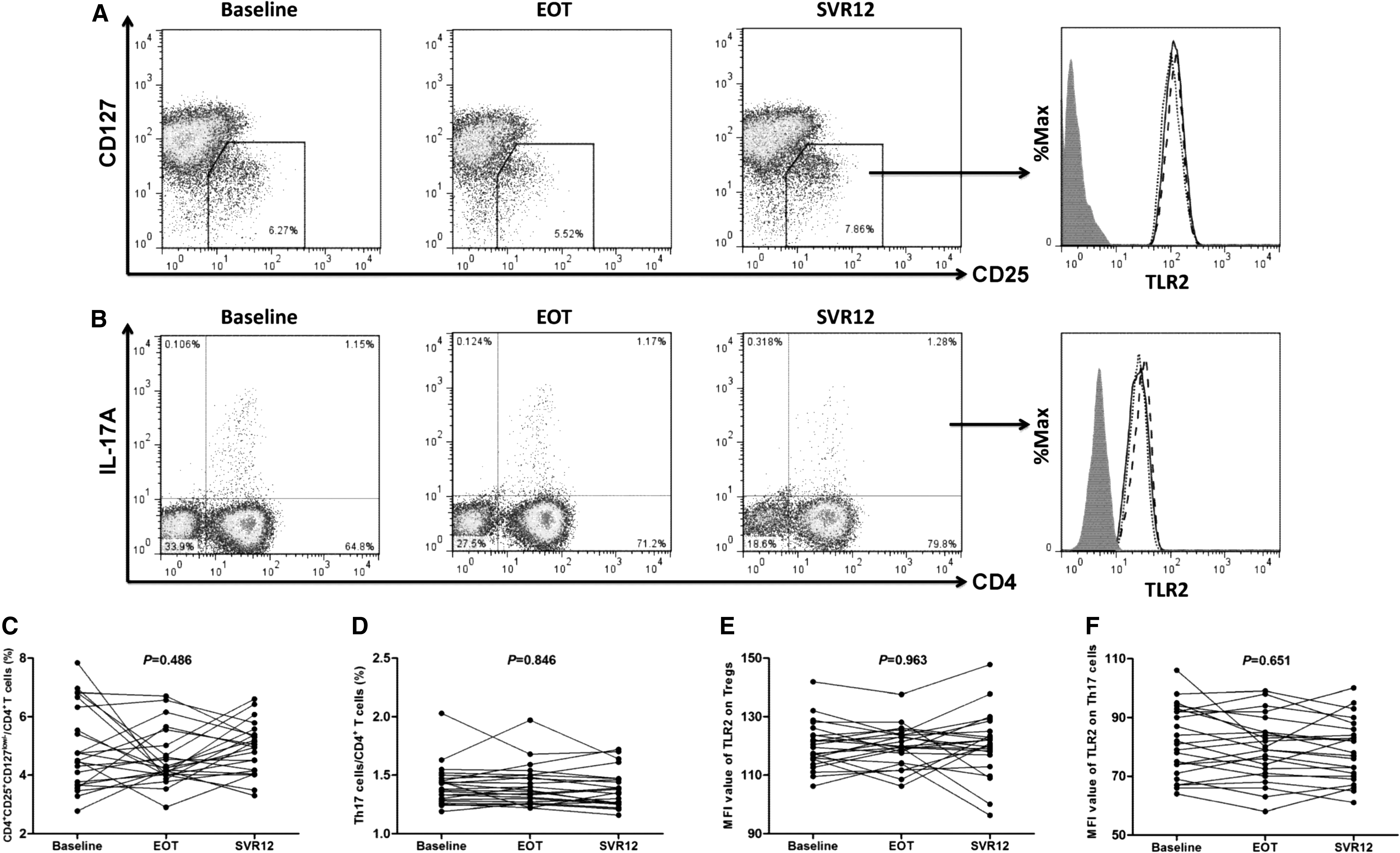
Comparison of CD4+CD25+CD127dim/− Tregs/Th17 cell proportion and TLR2 expression in response to Daclatasvir plus asunaprevir therapy. PBMCs were isolated from peripheral bloods which were collected at baseline, EOT, and SVR12 in all enrolled 23 chronic hepatitis C patients. Representative flow dots of
TLR2 stimulation did not affect the proliferation and Tregs/Th17 cell proportion
A measure of 106 of PBMCs was cultured with or without PamCsk4 stimulation in the presence of anti-CD3/CD28 for 48 h. CCK-8 results revealed that Pam3Csk4 stimulation did not influence the cellular proliferation of PBMCs isolated from baseline [(2.89 ± 0.73) × 106 vs. (2.94 ± 0.78) × 106; Paired t test, p = 0.556, Fig. 2A], EOT [(2.80 ± 0.73) × 106 vs. (2.91 ± 0.76) × 106; Paired t test, p = 0.147, Fig. 2A], and SVR12 [(2.62 ± 0.71) × 106 vs. (2.71 ± 0.80) × 106; Paired t test, p = 0.219, Fig. 2A]. Pam3Csk4 stimulation also did not affect the frequencies of CD4+CD25+CD127dim/− Tregs and Th17 cells within cultured PBMCs from baseline, EOT, or SVR12 (Paired t tests, all p > 0.05, Fig. 2B, C). There were no significant differences of either cellular proliferation or Tregs/Th17 frequency between patients with elevated ALT and those with normal ALT at baseline (all p > 0.05).

The influence of TLR2 stimulation on cellular proliferation and CD4+CD25+CD127dim/− Tregs/Th17 cells. 106 of PBMCs, which isolated from all enrolled 23 chronic hepatitis C patients, were cultured with or without Pam3Csk4 stimulation in the presence of anti-CD3/CD28 for 48 h.
TLR2 stimulation enhanced the suppressive function of Tregs from baseline, but not Tregs from EOT and SVR12
We sorted highly pure CD4+CD25+CD127dim/− Tregs and CD4+CD25− T cell subsets from ten chronic hepatitis C patients at baseline, EOT, and SVR12. Tregs were stimulated with or without Pam3Csk4 for 24 h. Tregs were washed twice, and 2.5 × 104 of Tregs was cocultured with 105 of autologous CD4+CD25− T cells with anti-CD3/CD28 for another 48 h. Cells and supernatants were harvested for proliferation and cytokine production analyses. There were no significant differences in the suppressive functions of Tregs among baseline, EOT, and SVR12 either before or post Pam3Csk4 stimulation (one-way ANOVA, p > 0.05, Fig. 3A). The presence of Pam3Csk4 remarkably enhanced the suppressive activity of Tregs from baseline, which presented as downregulation of cellular proliferation in coculture system [(2.74 ± 0.09) × 105 vs. (2.65 ± 0.14) × 105; Paired t test, p = 0.0071, Fig. 3A]. However, this enhancement effect failed to achieve statistical differences of Tregs from either EOT [(2.73 ± 0.06) × 105 vs. (2.66 ± 0.12) × 105; Paired t test, p = 0.094, Fig. 3A] or SVR12 [(2.68 ± 0.11) × 105 vs. (2.65 ± 0.07) × 105; Paired t test, p = 0.176, Fig. 3A]. Moreover, we also measured IL-10, IL-35, IL-17, and IL-22 production in the supernatants of cocultured cells. Pam3Csk4 stimulation notably increased the production of IL-10 in all groups (Baseline: 48.00 [34.97, 90.64] pg/mL vs. 63.70 [42.92, 128.1] pg/mL, Wilcoxon matched pairs test, p = 0.014; EOT: 55.50 [46.92, 102.4] pg/mL vs. 62.40 [43.92, 136.8] pg/mL, Wilcoxon matched pairs test, p = 0.047; SVR12: 52.50 [38.02, 104.2] pg/mL vs. 89.20 [58.69, 121.9] pg/mL, Wilcoxon matched pairs test, p = 0.032, Fig. 3B). Furthermore, treatment with Pam3Csk4 only robustly enhanced IL-35 secretion in baseline cocultured cells (54.63 ± 16.79 pg/mL vs. 94.15 ± 20.75 pg/mL, Paired t test, p = 0.0002, Fig. 3C), but not in EOT (47.86 ± 9.05 pg/mL vs. 46.19 ± 19.58 pg/mL, Paired t test, p = 0.768, Fig. 3C) or SVR12 (50.70 ± 30.07 pg/mL vs. 49.64 ± 14.70 pg/mL, Paired t test, p = 0.761, Fig. 3C) cocultured cells. More importantly, IL-35 production was significantly reduced at EOT and SVR12 in comparison with baseline after Pam3Csk4 stimulation (SNK-q tests, p < 0.0001, Fig. 3C). However, there were no significant differences of IL-17 and IL-22 expression in all groups in response to Pam3Csk4 stimulation (Paired t tests, all p > 0.05, Fig. 3D, E). Interestingly, we found that four patients showed increasing IL-17 production, while six patients revealed elevated IL-22 secretion. All those four patients with increasing IL-17 also produced higher level of IL-22 by CD4+ T cells.
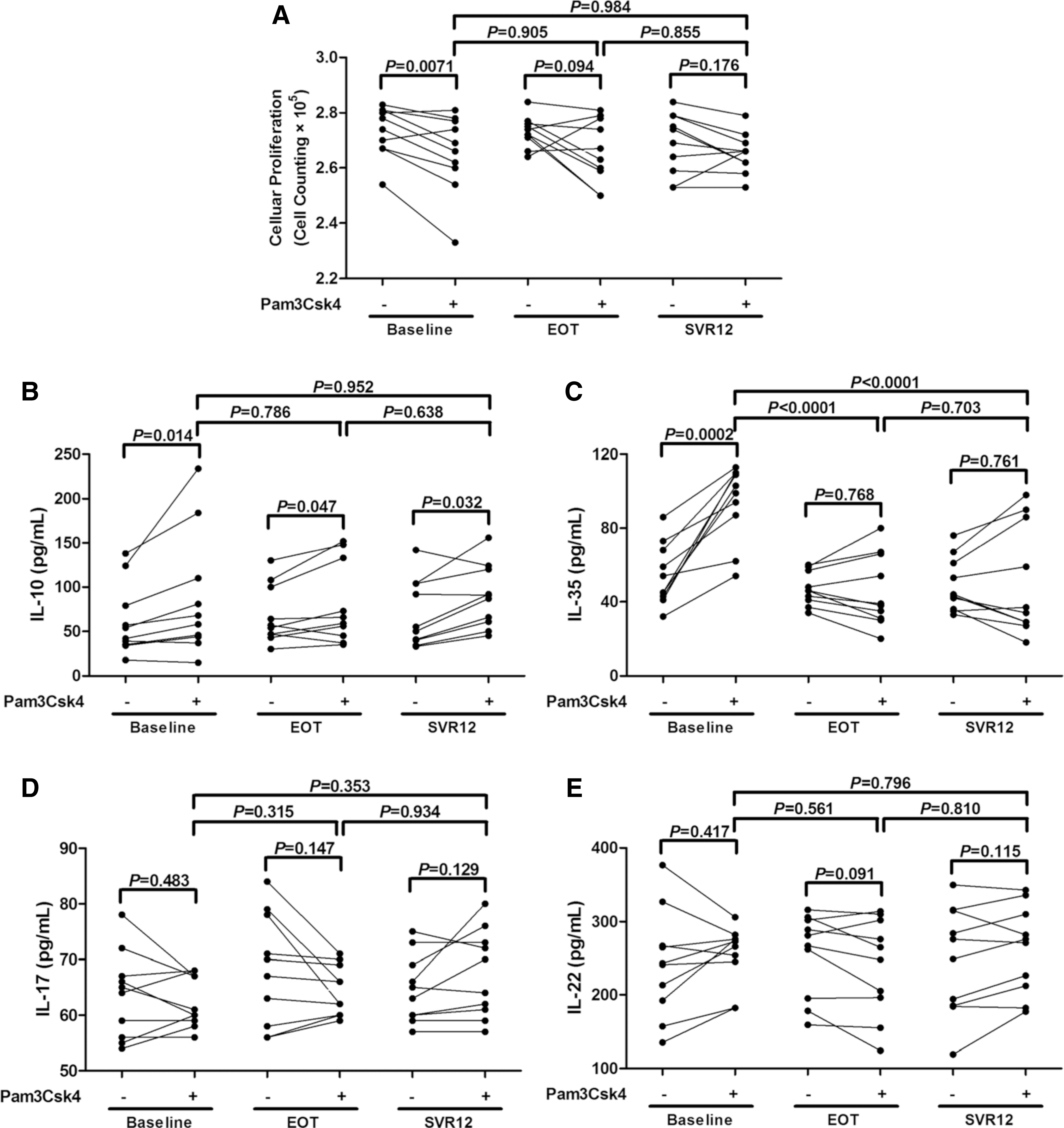
The influence of TLR2 stimulation on the suppressive capacity of CD4+CD25+CD127dim/− Tregs. CD4+CD25+CD127dim/− Tregs were purified from 10 chronic hepatitis C patients and were stimulated with or without Pam3Csk4 for 24 h and washed twice. 2.5 × 104 of Tregs was cocultured with 105 of autologous CD4+CD25− T cells with anti-CD3/CD28 for another 48 h.
TLR2 stimulation did not promote Th17 differentiation of Tregs, but only elevated Th17 cell frequency of CD4+ T cells from baseline
Our previous study revealed that TLR2 stimulation did not enhance a Th17 shift from CD4+CD25+CD127dim/− Tregs in treatment-naive chronic hepatitis C patients (13). Herein, we analyzed the effect of TLR2 stimulation on the differentiation of CD4+CD25+CD127dim/− Tregs in DAA-experienced chronic hepatitis C patients. A measure of 2.5 × 104 of purified CD4+CD25+CD127dim/− Tregs from the above ten patients was stimulated with or without Pam3Csk4 in a RPMI1640 supplemented with 10% of FBS and anti-CD3/CD28 for 96 h. Tregs did not elevate CCR4 and CCR6 expression in response to TLR2 stimulation in all three groups (Paired t tests, all p > 0.05, Fig. 4A, B). Moreover, the production of IL-17 and IL-22 by Tregs in the cultured supernatants was also investigated. There were no remarkable differences in IL-17 and IL-22 expression in response to TLR2 stimulation (Paired t tests, all p > 0.05, Fig. 4C, D).
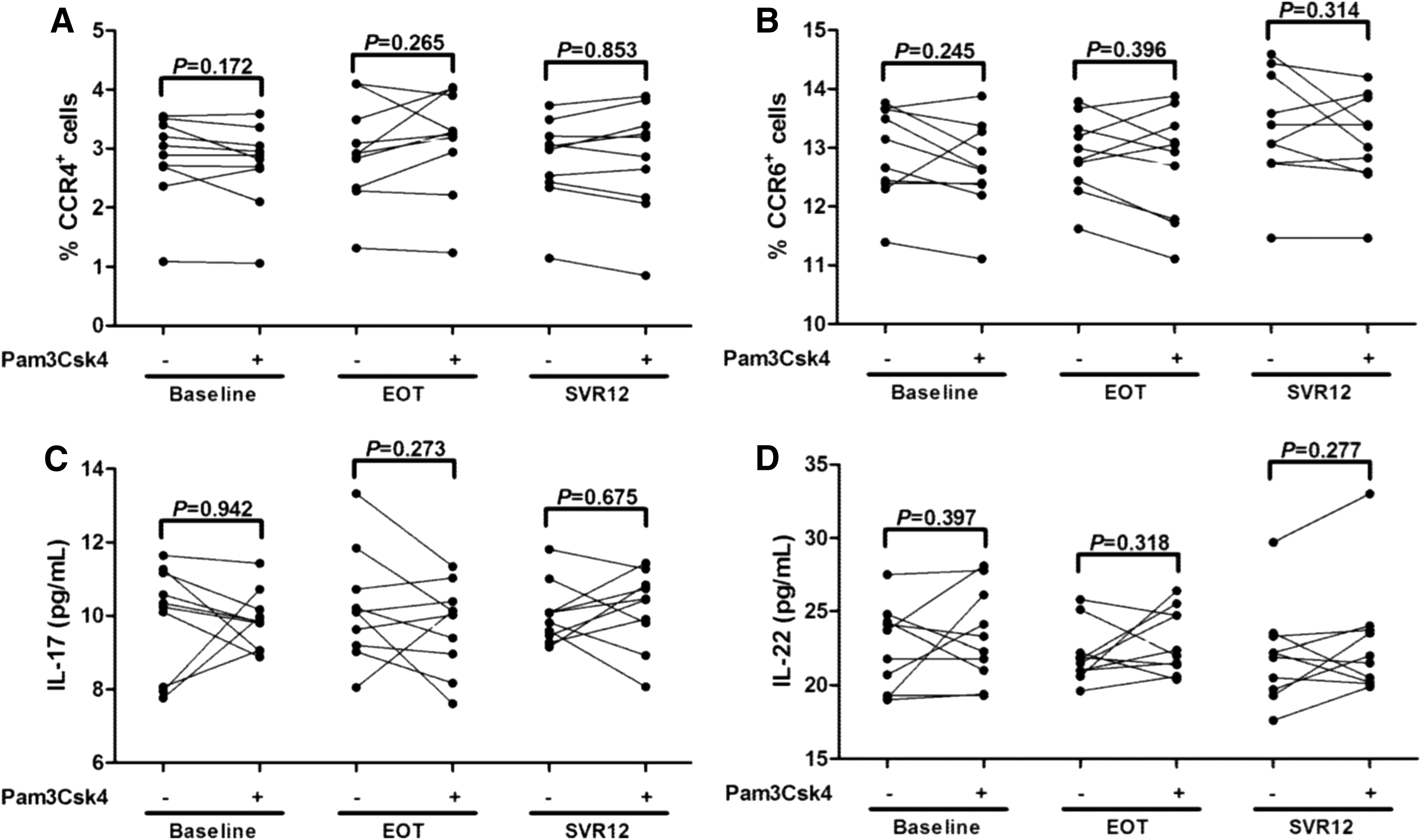
The influence of TLR2 stimulation on the CCR4/CCR6 expression and IL-17/IL-22 production by CD4+CD25+CD127dim/− Tregs. CD4+CD25+CD127dim/− Tregs were purified from ten chronic hepatitis C patients, which were used in Figure 3. 2.5 × 104 of purified Tregs were stimulated with or without Pam3Csk4 for 96 h. Cells were harvested and stained for the expression of
Furthermore, 105 of purified CD4+ T cells from the other 13 chronic HCV-infected patients were cultured in the presence or absence of Pam3Csk4 for 96 h. Cells and supernatants were harvested for further experiments. The percentage of Th17 cells was notably increased in response to TLR2 stimulation in CD4+ T cells at baseline (4.30 ± 1.23% vs. 5.05 ± 1.69 pg/mL, Paired t test, p = 0.039, Fig. 5A). There were also elevated trends of Th17 cell frequency in response to TLR2 stimulation at EOT and SVR12, but these differences just failed to achieve significances (Paired t tests, p = 0.065 and p = 0.071, respectively, Fig. 5A). IL-17 and IL-22 production in the supernatants revealed similar trends with the changes of Th17 proportion in response to TLR2 stimulation. Both IL-17 and IL-22 secretion was significantly elevated in response to TLR2 stimulation at baseline (IL-17: 30.85 ± 8.81 pg/mL vs. 59.48 ± 8.94 pg/mL, Paired t test, p < 0.0001, Figure 5B; IL-22: 290.5 ± 71.42 pg/mL vs. 324.4 ± 71.58 pg/mL, Paired t test, p = 0.0059, Fig. 5C) and EOT (IL-17: 32.08 ± 9.26 pg/mL vs. 39.23 ± 16.24 pg/mL, Paired t test, p = 0.043, Fig. 5B; IL-22: 283.9 ± 70.14 pg/mL vs. 315.3 ± 91.16 pg/mL, Paired t test, p = 0.029, Fig. 5C). Furthermore, IL-17 production was notably decreased at EOT and SVR12 in comparison with baseline after Pam3Csk4 stimulation (SNK-q tests, p = 0.0008 and p < 0.0001, Fig. 5B). However, there were no remarkable differences in IL-17 and IL-22 between cells with or without TLR2 stimulation at SVR12 (Paired t tests, all p > 0.05, Fig. 5B, C).
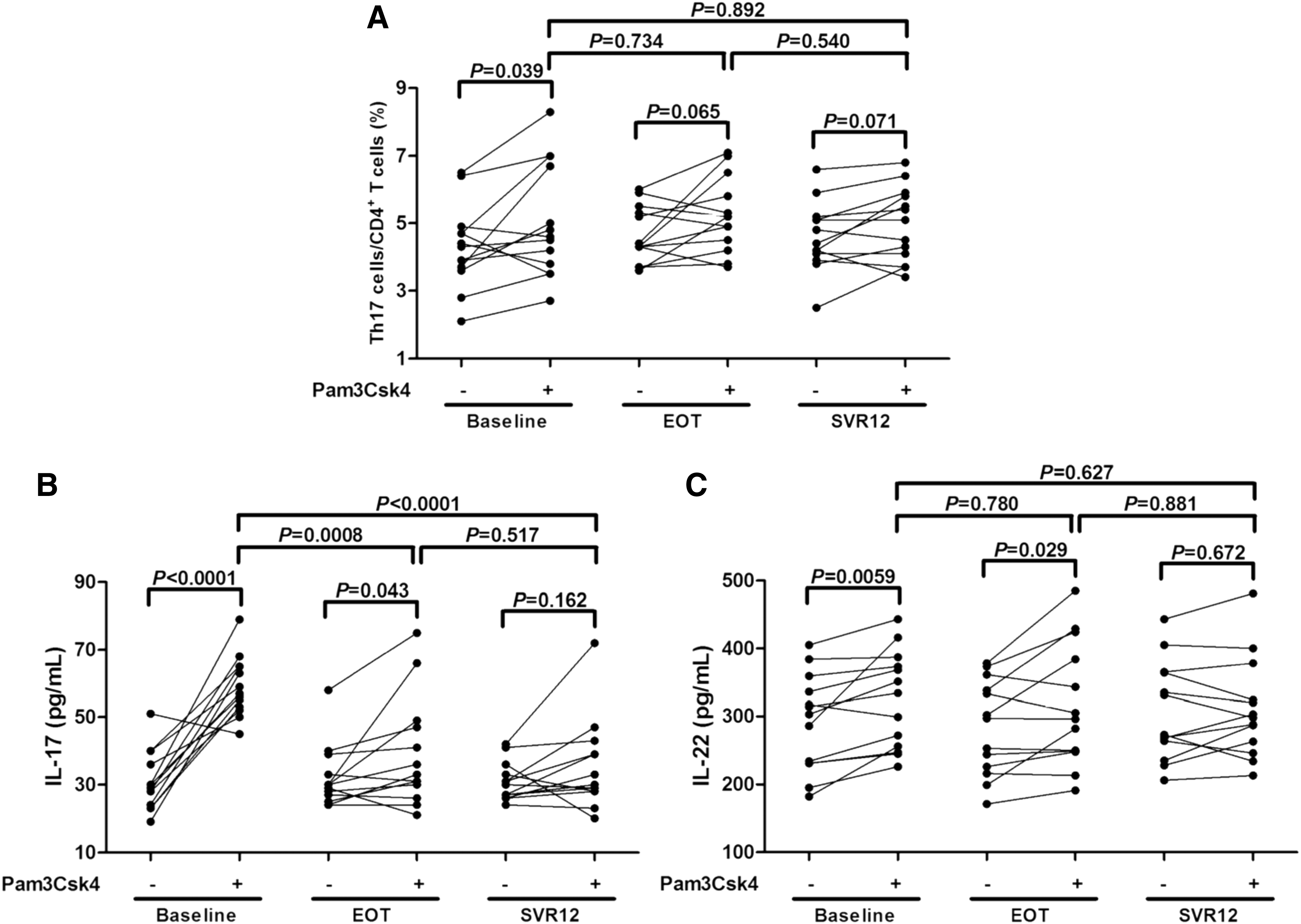
The influence of TLR2 stimulation on Th17 cell proportion and IL-17/IL-22 production in CD4+ T cells. CD4+ T cells were purified from other 13 chronic hepatitis C patients. 105 of purified CD4+ T cells were stimulated with or without Pam3Csk4 for 96 h. Cells and supernatants were harvested for further experiments.
Discussion
In this study, treatment with daclatasvir plus asunaprevir continuously inhibited HCV replication, resulting in SVR in all enrolled patients. Although DAA therapy did not induce the downregulation of either Tregs/Th17 percentage or TLR2 expression in chronic hepatitis C patients, the responsiveness of Tregs and Th17 cells to TLR2 stimulation was significantly reduced in response to successful DAA treatment. The current results indicated a potential novel mechanism of DAA-induced immunoregulation in chronic hepatitis C patients.
Chronic HCV infection led to the imbalance between Tregs and Th17 cells, which mainly presented as the enhancement of both percentage and suppressive activity of Tregs (7). This imbalance could be modulated by various molecules and cytokines (such as TLR2 and IL-35) in chronic hepatitis C patients (12,13). Conventional IFN-based treatment on Tregs/Th17 cells remained highly controversial in chronic HCV infection (19). Hao et al. reported that inhibition of HCV RNA by peg-IFN-α2 plus ribavirin therapy was closely associated with decreased Tregs frequency (7), whereas a persistence of Tregs in blood stream and liver in patients who achieved SVR to IFN-based therapy was shown by Boonstra and colleagues (4,23). Sofosbuvir plus daclatasvir increased the numbers of Tregs, Th17 cells, T follicular helper cells, and memory B cells in HCV-associated cryoglobulinemia vasculitis patients (21). In contrast, Langhans et al. indicated that sofosbuvir based antiviral therapy did not reduce the proportion and activation status of Tregs even in the long term after HCV elimination (11). Consistent with the results from Langhans et al. (11), we found that daclatasvir plus asunaprevir did not normalize either Tregs frequency or increased TLR2 expression on Tregs in chronic hepatitis C patients. More importantly, we also first reported that neither Th17 percentage nor TLR2 expression on Th17 cells was affected in response to daclatasvir plus asunaprevir therapy. These results suggested that sustained inhibition of HCV replication by daclatasvir plus asunaprevir might not be paralleled with normalization of innate or adaptive immune response in chronic hepatitis C.
Sofosbuvir plus IFN therapy induced a transient elevation of coexpressing activation markers on Tregs (11), revealing a reprogramming of Tregs to effector T cells (8). Furthermore, DAA treatment also altered the inflammatory cytokine expression profile in chronic hepatitis C and HCV-induced carcinogenesis (5). Thus, the microenvironment, which was important for Tregs function and stability (1), was changed in response to DAA therapy. Schmiedel et al. revealed that praziquantel therapy decreased schistosoma-associated Tregs; however, their inhibitory function remained unaltered with comparable immunosuppressive cytokine production before and after treatment (22). In the present study, we investigated the changes of responsiveness of Tregs/Th17 cells to TLR2 stimulation following DAA treatment. DAA therapy did not affect the inhibitory capacity of Tregs, which presented as comparable cellular proliferation and IL-10/IL-35 production in coculture system among baseline, EOT, and SVR12. Consistent with our previous results, treatment with Pam3Csk4 enhanced the suppressive activity in treatment-naive chronic hepatitis C patients. However, TLR2 stimulation only slightly downregulated proliferation of cocultured cells without statistical differences, which was accompanied by comparable IL-35 production, but elevated IL-10 secretion in DAA-experienced patients. It was well accepted that mechanism of Tregs suppressive function included direct cell-to-cell contact and cytokine production (15). Since we used identical cell numbers in each coculture system and Pam3Csk4 stimulation did not affect cellular proliferation, the DAA-induced decreased responsiveness of Tregs to TLR2 stimulation might be partly due to reduced IL-35 secretion. Furthermore, similar findings were observed in TLR2 stimulation on Th17 cells. Pam3Csk4 stimulation induced elevated Th17 cells and IL-17/IL-22 production in cultured CD4+ T cells isolated from treatment-naive chronic hepatitis C patients. DAA treatment reduced responsiveness of Th17 cells to TLR2, although IL-17 and IL-22 production was not significantly decreased. Moreover, reduction production of IL-35 and IL-17 at EOT and SVR12 after Pam3Csk4 stimulation indicated decreased responsiveness of Treg and Th17 cells to TLR2 stimulation. Further experiments were needed for elucidation of the mechanism underlying the DAA-induced responsiveness of Tregs/Th17 cells to TLR2 stimulation.
Our previous study also showed that effective anti-HCV treatment by peg-IFN-α2 plus ribavirin elevated TLR2-induced shift of Tregs toward Th17-like phenotype and function without loss of suppressive capacity in chronic hepatitis C (13). We then further investigated Th17-like differentiation from Tregs in response to TLR2 stimulation. Unfortunately, we did not observe either increased CCR4/CCR6 expression or elevated IL-17/IL-22 production by Tregs, indicating that effective DAA treatment did not enhance TLR2-mediated Th17 differentiation in chronic hepatitis C. Thus, decreased responsiveness of Tregs and Th17 cells might not only be due to the inhibition of HCV RNA replication but also be associated with the DAA-induced alteration in microenvironment.
There were several limitations of this study. We did not analyze the responsiveness of liver-resident Tregs/Th17 cells to TLR2 stimulation in response to DAA therapy due to the ethical reasons. Moreover, we only enrolled chronic HCV genotype 1b infected patients who received daclatasvir plus asunaprevir therapy because these DAA-based regimens were first approved by China Food and Drug Administration in April 2017. Other DAAs, including sofosbuvir and daclatasvir, were approved in September 2017. Thus, it was impossible to analyze the SVR of chronic hepatitis C patients with sofosbuvir-based therapy. Further experiments would be performed for expanded DAA regimens on Tregs/Th17 cell function.
Conclusion
In summary, DAA therapy can persistently inhibit viral replication in almost all treated chronic hepatitis C patients. We revealed that DAA did not normalize the increased percentage of Tregs/Th17 cells or affect TLR2 expression on these cells. However, there was a significant reduction in the responsiveness of Tregs and Th17 cells to TLR2 stimulation. The current data suggested that DAAs might influence the interaction between innate and adaptive immune system, providing a novel mechanism underlying DAA-induced immunoregulation.
Footnotes
Author Disclosure Statement
No competing financial interests exist.