
Abstract
Pseudorabies virus (PRV) can spread along the peripheral nerves near the site of infection in the animals, and gradually migrates into the central nervous system, where it leads to the development of brain lesions. The aim of this study was to investigate the dynamics of microglia after PRV inoculation. A mouse model inoculated with PRV was established to study the interactions between PRV and microglia, microglial recruitment, and polarization effects. The mice were subcutaneously inoculated with different doses of PRV-Bartha K61 vaccine strain. The obtained results showed that mouse mortality rates increased with the applied doses of virus, and brain lesions, located in the brain tail and brain stem, were observed in each investigated group. Inflammatory cells were shown to infiltrate through the vasculature into perivascular cuff, and the number of microglia was increased as well. Mouse group treated with a medium infection dose demonstrated a high survival rate while developing serious brain lesions, and therefore, this dose was selected for further experiments. Immunohistochemistry, flow cytometry, and confocal laser scanning microscopy were used to analyze PRV-microglia interactions. After PRV inoculation, proliferating cell nuclear antigen (Pcna) and Iba1 double-positive cells were observed in the brain lesions, together with the activated microglia, suggesting that PRV can induce microglial proliferation and activation. Furthermore, 5-bromo-deoxy-uridine (BrdU) labeling demonstrated that microglial cells did not proliferate in situ and the proliferating cells originated from peripheral blood monocytes, mainly from the inflammatory monocytes (Ly6Chigh). In addition, microglia polarized into both M1 and M2 phenotypes by PRV infection. The results obtained in this study may help understand the development of pseudorabies infection and help improve the treatment, by recruiting and enhancing immune response.
Introduction
P
Microglia are the only resident phagocytic cells in the CNS, and normally, they are in a highly branched resting or ramified state. Microglial cells constitute 5–20% of total glial cells in rodents and exhibit several features that can distinguish them from other macrophage populations. Microglia can communicate with the surrounding neurons and other glial cells through the branches emerging from the cell body (31), and they are activated only to clear cellular debris or on brain damage, which leads to microglial cell proliferation. Microglia can rapidly respond to external stimuli and adopt an amoeboid-activated phenotype (30,39), and on activation, these cells express several molecules, such as Iba1 and CD45. The activated microglia, resting microglia, and bone marrow-derived microglia can be distinguished using flow cytometry analyses, since the resting microglia express low levels of CD45 (CD45low), followed by the activated microglia (CD45int), while the bone marrow-derived microglia express the highest level of CD45 (CD45high) (13). In different models of brain inflammation, microglia proliferate, which can be through rapid in situ proliferation or they can be derived from migratory precursor cells (6,13,15).
The bone marrow-derived cells are Gr1-positive, monocytes (Gr1+Ly6C+) and granulocytes (Gr1+Ly6G+). Mice have two subsets of monocytes, inflammatory monocytes, expressing CCR2+CX3CR1lowLy6Chigh, and resident monocytes, expressing CCR2−CX3CR1highLy6Clow (7,8,33). The functions of monocyte-derived macrophages and microglia-derived macrophages are different because of their distinct developmental origin and renewal mechanism (17,18,40). Microglia can polarize into both M1 and M2 phenotypes after stimuli, resulting in altered cytokine expression. M1 phenotype is associated with the activation of Thelper1 (Th1)-mediated immune response and these cells play proinflammatory functions, while M2 macrophages have regulatory functions during tissue repair, remodeling, and they promote Th2 immune response (2,20,35).
In the present study, our investigations, using different titers of PRV-Bartha K61 vaccine-inoculated mice, aimed to explore the dynamic changes of microglia during viral infection. We demonstrated that the viral spread into the CNS led to the development of brain lesions and induced changes in microglial numbers, morphology, phenotype, and function, activating the immune response.
Materials and Methods
Cell culture and viral strain
PK15 cells were used for the expansion of the virus. The cells were maintained in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum (FBS) in 25 cm2 flasks at 37°C and 5% CO2. PRV-Bartha K61 strain was generated from the live vaccine by dissolving the powder in 0.5 mL DMEM, which was then inoculated in PK15 cells. After 48 h, the flasks were frozen, and on thawing, the supernatants, containing viral particles, were collected by centrifugations. These samples were stored at −70°C until further use.
Tissue culture infectious dose (TCID)50 was determined. PK15 cells were seeded into 96-well plates at a density of 1 × 103 cells/well in 100 μL of DMEM, and incubated overnight at 37°C in an atmosphere containing 5% CO2. Afterward, the medium was removed and replaced with 100 μL of different dilutions of viral supernatant samples. After 2 h of incubation, 100 μL of DMEM with 2% FBS was added to each well. After 24 h of incubation, cytopathic effects were measured, and TCID50 was calculated.
Mouse model
Six-week-old male Balb/c mice were used. Mice were maintained under pathogen-free conditions with regular light/dark cycles, and sufficient food and water. Mice were anesthetized by diethyl ether before inoculation, and subcutaneous injection on the outside of the right leg containing 100 μL of viral supernatant (6 × 104 TCID50, 1 × 105 TCID50, and 2.7 × 105 TCID50) or DMEM was administered. Twelve mice were included in each group, in all experiments. All mice were killed by decapitation. All animal studies were approved by the Animal Care and Use Committee of Harbin Veterinary Research Institute, Chinese Academy of Agricultural Sciences.
Antibodies
The following antibodies were used in this study: rabbit anti-PRV antibody (1:25; USBio), rabbit anti-Iba1 polyclonal antibody (1:100; Wako), mouse anti-proliferating cell nuclear antigen (Pcna) IgG (1:100; Santa Cruz), rat anti-5-bromo-deoxy-uridine (BrdU) monoclonal antibody (1:100; Abcam), goat anti-rabbit IgG (whole molecule)-peroxidase antibody (1:200; Sigma-Aldrich), biotin-labeled goat anti-rat IgG(H+L) antibody (1:200; Abcam), streptavidin, horseradish peroxidase (HRP) conjugate (1:200; Millipore), goat anti-rat IgG(H+L) Texas Red conjugate (1:200; Vector Laboratories), goat anti-rabbit IgG (H+L) Texas Red conjugate (1:200; Vector Laboratories), Fluoroshield with DAPI (Sigma-Aldrich), CD11b microbeads (1:100; Miltenyi Biotec), FITC rat anti-mouse Ly-6C (1:200; BD Biosciences), antigen-presenting cell (APC) anti-BrdU (1:200; BioLegend), PE rat anti-mouse Ly6G (1:200; BD Biosciences), PerCP rat anti-mouse CD45 (1:200; BD Biosciences), and APC/Cy7 rat anti-mouse Gr1 (1:200; BD Biosciences).
BrdU labeling
BrdU can selectively incorporate into DNA at the S phase of the cell cycle, which can be used for the tracing of microglia. One group of animals received 75 mg/kg BrdU intraperitoneally at 132 and 138 h postinoculation, the second group received the same injection at 120 h after the inoculation, while the third group represented a control group. Mice from both groups were sacrificed at day 6 postinoculation.
Immunohistochemistry and immunofluorescence analyses
Fixed brains were paraffin embedded and cut into 4-μm-thick sections. All brains sections were deparaffinized in xylene and rehydrated with water using decreasing concentrations (100%, 95%, and 70%) of ethanol. Antigen retrieval was performed by immersing the slides in citrate acid (pH 7.4)/sodium citrate buffer solution (pH 8.0) at 121°C for 30 min, as given in the Materials and Methods section of the revised article, and when the sections cooled down to room temperature, they were incubated in the methyl alcohol containing 0.3% H2O2 for 30 min. Afterward, the sections were rinsed with tris buffered saline (TBS) three times and they were subsequently blocked with 8% skimmed milk. Following this, primary antibodies were added, and the sections were incubated at 4°C overnight. The following day, the treated sections were rinsed and incubated with the secondary antibody. For immunohistochemistry (IHC), if the secondary antibody used was not coupled with HRP, it was added separately, and the sections were incubated for 40 min at 37°C. The samples were stained with DAB and observed under a light microscope. For immunofluorescence analyses, fluorescence-labeled secondary antibodies were added and the samples were incubated for 1 h at room temperature in dark.
Flow cytometry preparation
The isolation of brain microglia was achieved by magnetic-activated cell sorting, using CD11b magnetic beads that bind microglia specifically. Using the Neural Tissue Dissociation kit, the brains were digested for 35 min at 37°C. Tissue debris was removed by passing the cell suspension through a 70-μm cell strainer. Following centrifugation, cells were resuspended in 30% Percoll and centrifuged at 400 g for 25 min. The supernatant containing myelin was discarded, and the pelleted cells were washed with phosphate-buffered saline. The final cell density was 2 × 105, cells were labeled with antibodies, fixed in 4% paraformaldehyde, and stored at 4°C until flow cytometry analyses.
Reverse transcription-polymerase chain reaction
Microglial cells were extracted from mouse brain at days 3 to 7 after PRV inoculation. Total RNA was extracted from the microglia using the RNA Extraction Kit (Takara, Shiga, Japan), and cDNA was synthesized from 5 μL of total RNA using the Reverse Transcription Kit (Takara) in 20 μL reaction volume. Polymerase chain reaction (PCR) was performed using 0.8 μM primers and 2 μL of cDNA. The sense and antisense primers for target genes are listed in Table 1. Amplification was performed in a Mastercycler ep realplex thermal cycler (Eppendorf) at 95°C for 5 min, followed by 30 cycles at 94°C for 30 sec, 58°C for 30 sec, 72°C for 1 min, and by a final elongation step at 72°C for 10 min. PCR analyses were carried out using Bio-Rad system software.
F, forward; R, reverse.
Statistical analyses
Each experiment was performed independently at least three times. Results are expressed as mean ± standard deviation. Statistical analyses were performed using GraphPad software. Acquired FACS data were analyzed using the flow cytometry software FlowJo. Quantification of cell populations of interests was performed based on flow cytometry analyses.
Results
Mouse model
The survival rate of infected mice was shown to depend on the selected virus strain, virus titer, and infection route (subcutaneous injection on the outside of the right leg). In our study, three different virus titers were used. First mouse deaths were observed at 6 days postinjection (dpi) in each investigated group, while at 7 dpi, a significant difference in mouse survival rates between the groups was observed. The survival rates of mice inoculated with 6 × 104 TCID50, 1 × 105 TCID50, and 2.7 × 105 TCID50 of PRV were shown to be 93.4%, 93.3%, and 85.7% at 6 dpi, and 93.4%, 66.67%, and 14.3% at 7 dpi, respectively (Fig. 1a). In contrast, no changes were observed in the normal mouse brain (Fig. 1b) and the brains of mice inoculated with DMEM (Fig. 1c). The infiltration of inflammatory cells into the perivascular cuff was observed in the infected brains of both PRV-inoculated groups, together with microglial proliferation, which occurred mainly at the brain tail and brain stem. The severity of pathological changes depended on the infection dose (Fig. 1d–f). The virus successfully infected the mouse brains, primarily the neurons, but microglial cells were infected as well (Fig. 1g). The group infected with 1 × 105 TCID50 was selected for further experiments.
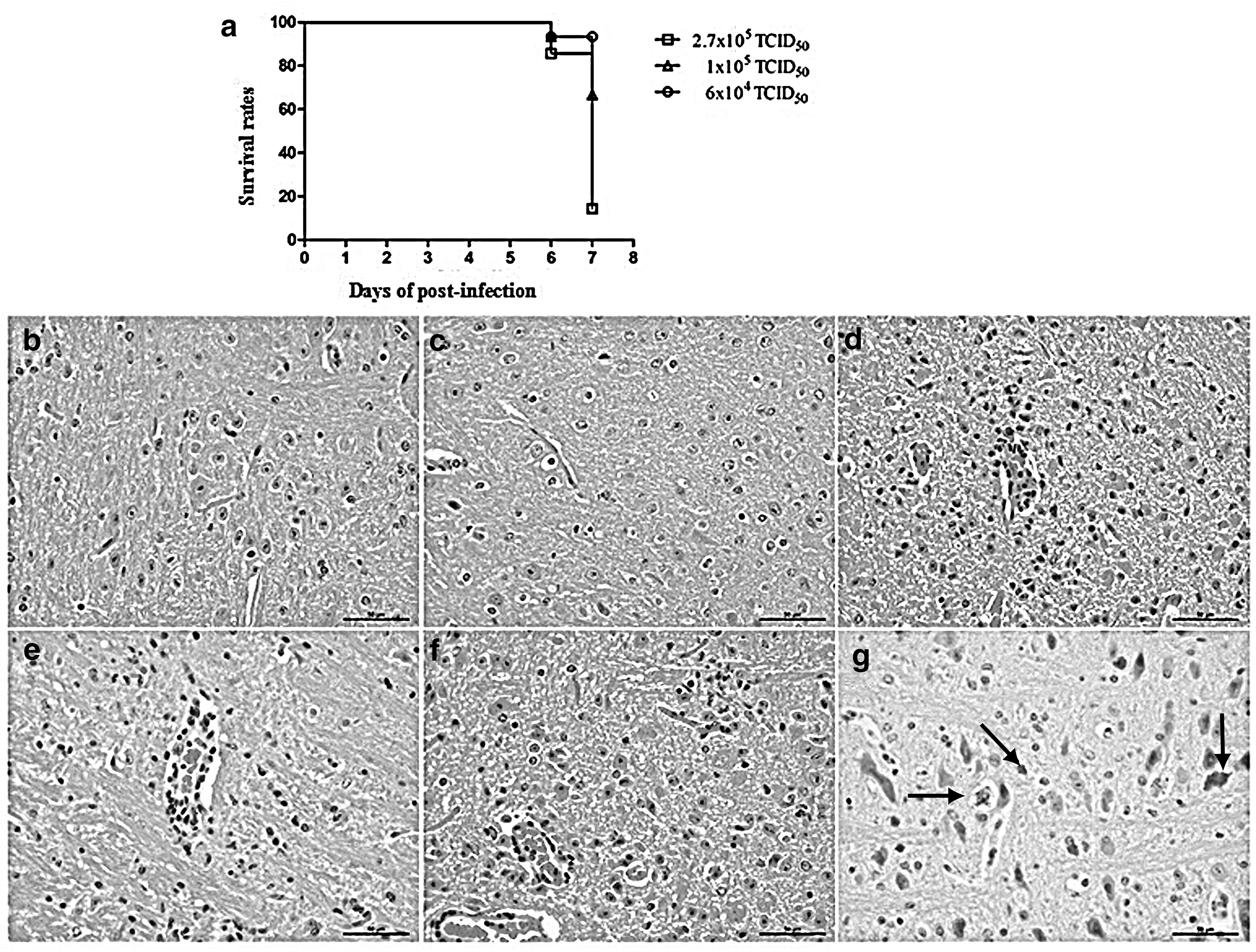
Validation of PRV infection mouse model. Survival rates were obtained using 12 mice per group.
Activation and proliferation of microglial cells
In infected mouse brains, the microglia were shown to be activated and to proliferate. Pcna antibody is used for the labeling of proliferating cells and Iba1 represents a specific microglial marker. IHC results showed that Pcna-positive cells were rare in brain samples obtained from the control mice at 6 dpi (Fig. 2a), while in the brains of PRV-inoculated mouse, a large number of Pcna-positive cells were observed in perivascular areas, and scattered around them (Fig. 2b). Double-positive Pcna and Iba1 cells were observed in the lesions of PRV-infected brain samples (Fig. 2c–f).
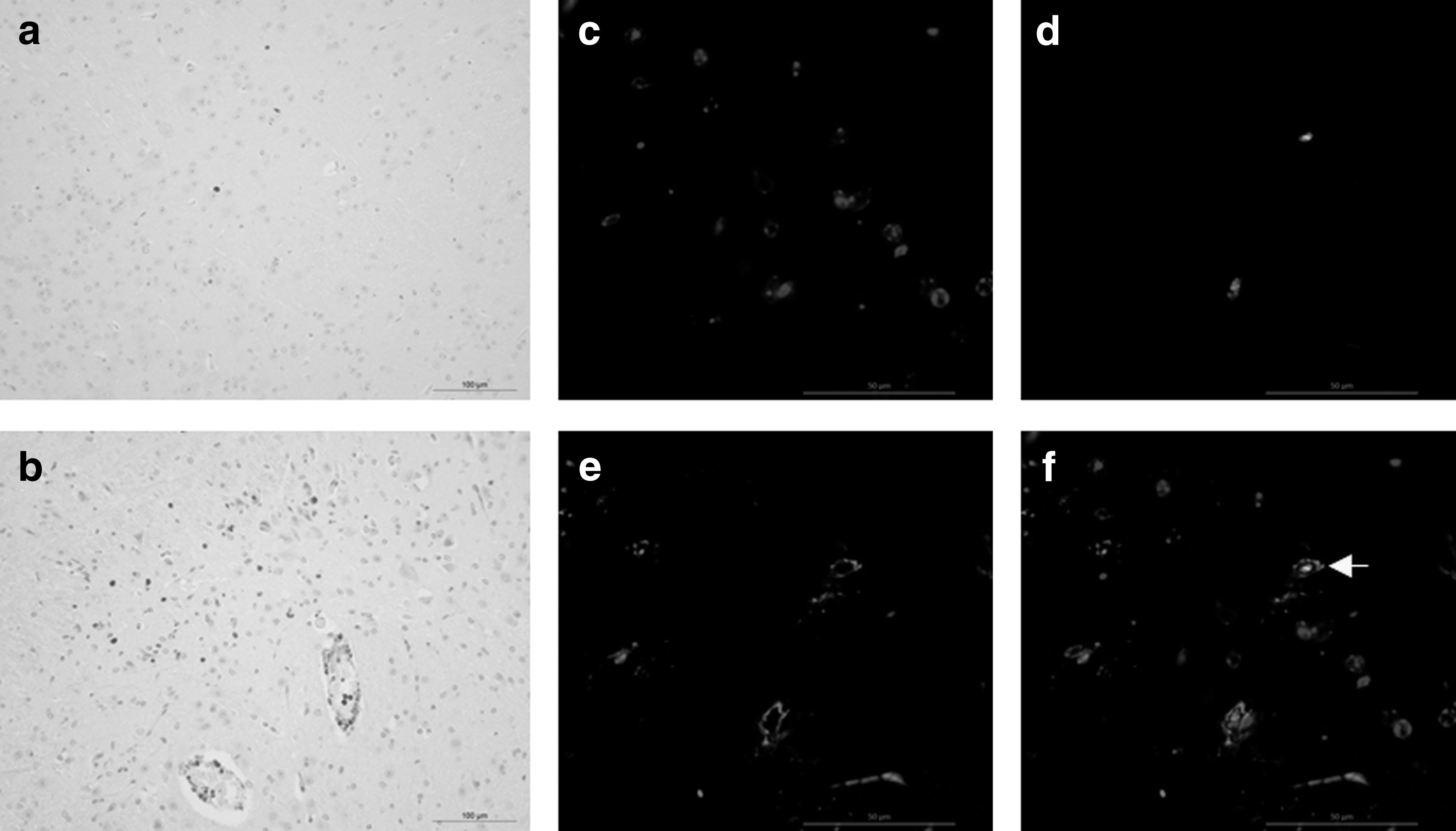
Microglia proliferation in PRV-infected brains. IHC was performed using the formalin-fixed tissue, which had undergone antigen retrieval in citrate buffer, pH 6.0, at 121°C for 10 min. DAB chromogen substrates were used for antigen visualization. Pcna-positive cells in brain tissues of mouse from group
Iba1 can be used for the labeling of both resting and activated microglia. Microglial cells were shown to be activated by PRV inoculation, since the morphology of these cells changed in PRV-infected mouse brains. Microglial cells in the brains of control animals were shown to have resting morphology with small-cell bodies and pronounced cellular processes (Fig. 3a), while the morphology of the activated cells in PRV-infected brains changed to round shape, together with the shrinking of cellular processes (Fig. 3b). Using CD11b microbeads to extract the microglia from mouse brains, we showed that the expression levels of CD45 were increased in the activated microglia. CD11b+CD45low expression is a marker of resting microglia, CD11b+CD45int expression, a marker of activated microglia, while macrophages show CD11b+CD45high expression. Our results showed that after PRV infection, the percentage of CD11b+CD45int cells increased in the infected brain (Fig. 3c) in comparison with the control brain (Fig. 3d). In addition, PRV and Iba1 double-positive cells were found in the infected brain, showing that the activated microglia support PRV replication (Fig. 4).
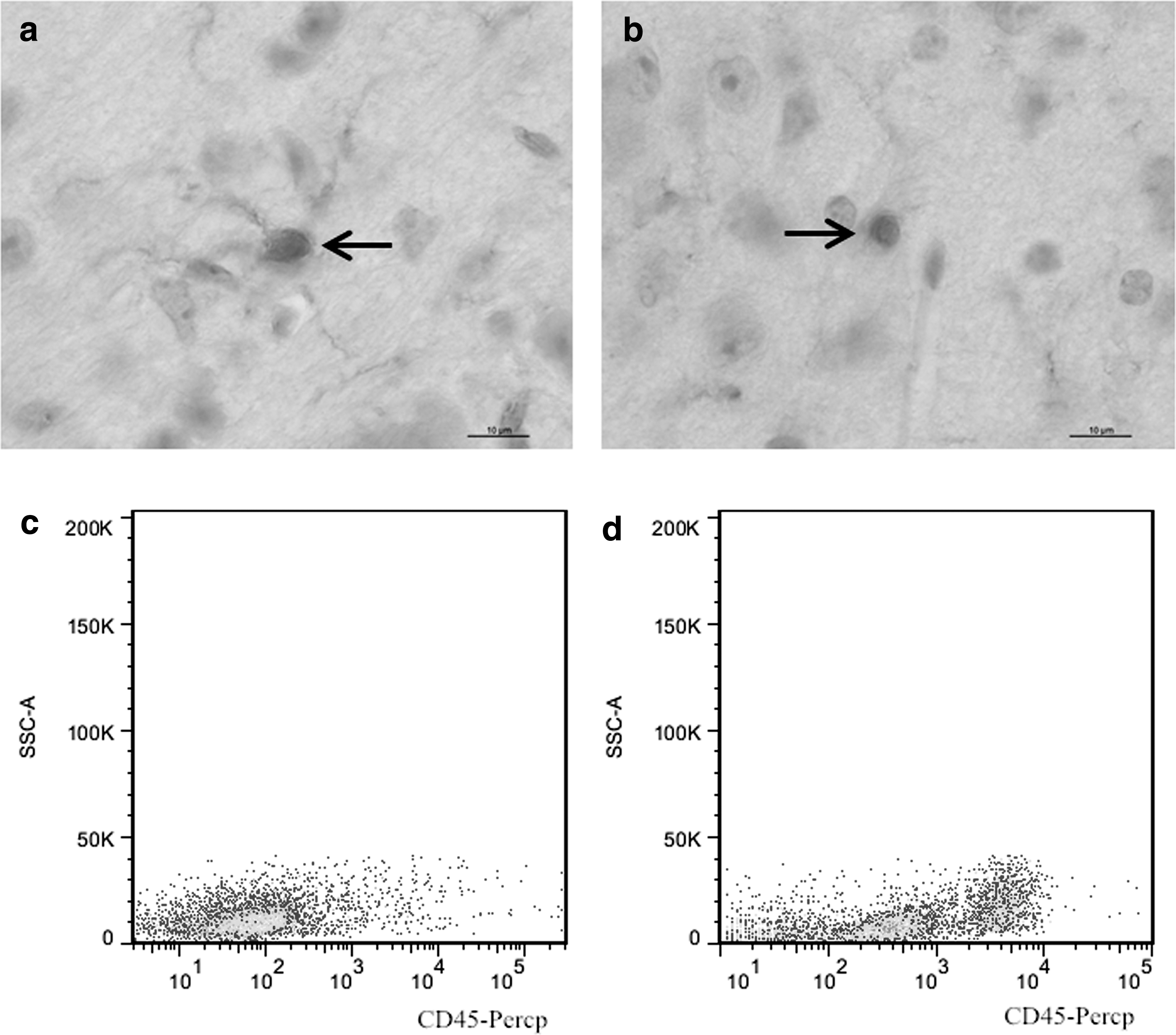
PRV infection induces the activation of microglia. Sections were incubated with an anti-Pcna antibody using IHC staining. The morphology of cells in brain sections of control group
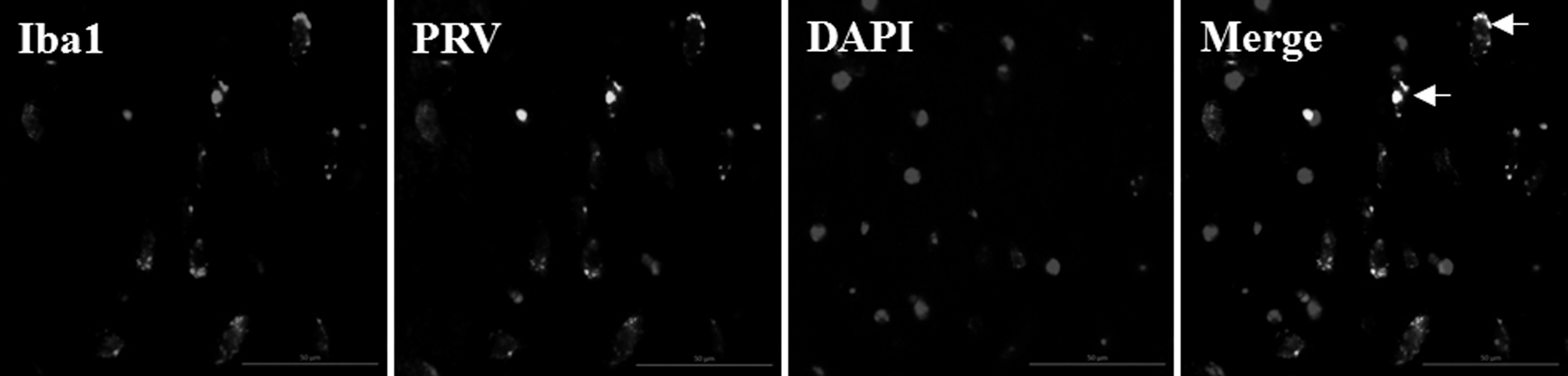
Phenotypes of single cell supporting viral replication in PRV-infected brains. Sections were incubated with an anti-Iba1 antibody, a microglia cell marker, and an antibody to label PRV using an indirect immunofluorescence assay. Multiple positive foci were visible in PRV-infected brains (arrow). Nuclei were stained with DAPI. OM, 50 μM.
Microglia did not proliferate in situ
One PRV-infected mouse group was injected with BrdU at 12 and 6 h before the analyses, while the other group received one BrdU dose, at 24 h before the mouse sacrifice, allowing the labeling of in situ microglial proliferation. Since the microglial precursors require 2–3 days to migrate to the brain lesions, these cells are not labeled with BrdU. BrdU-positive cells were observed only in the brains of mice that received multiple BrdU injections (Fig. 5a), while in the control brains and the brains obtained from mice injected with BrdU at 4 dpi, BrdU-positive cells were not observed (Fig. 5b, c). Similar results were obtained using flow cytometry (Fig. 5d–f).
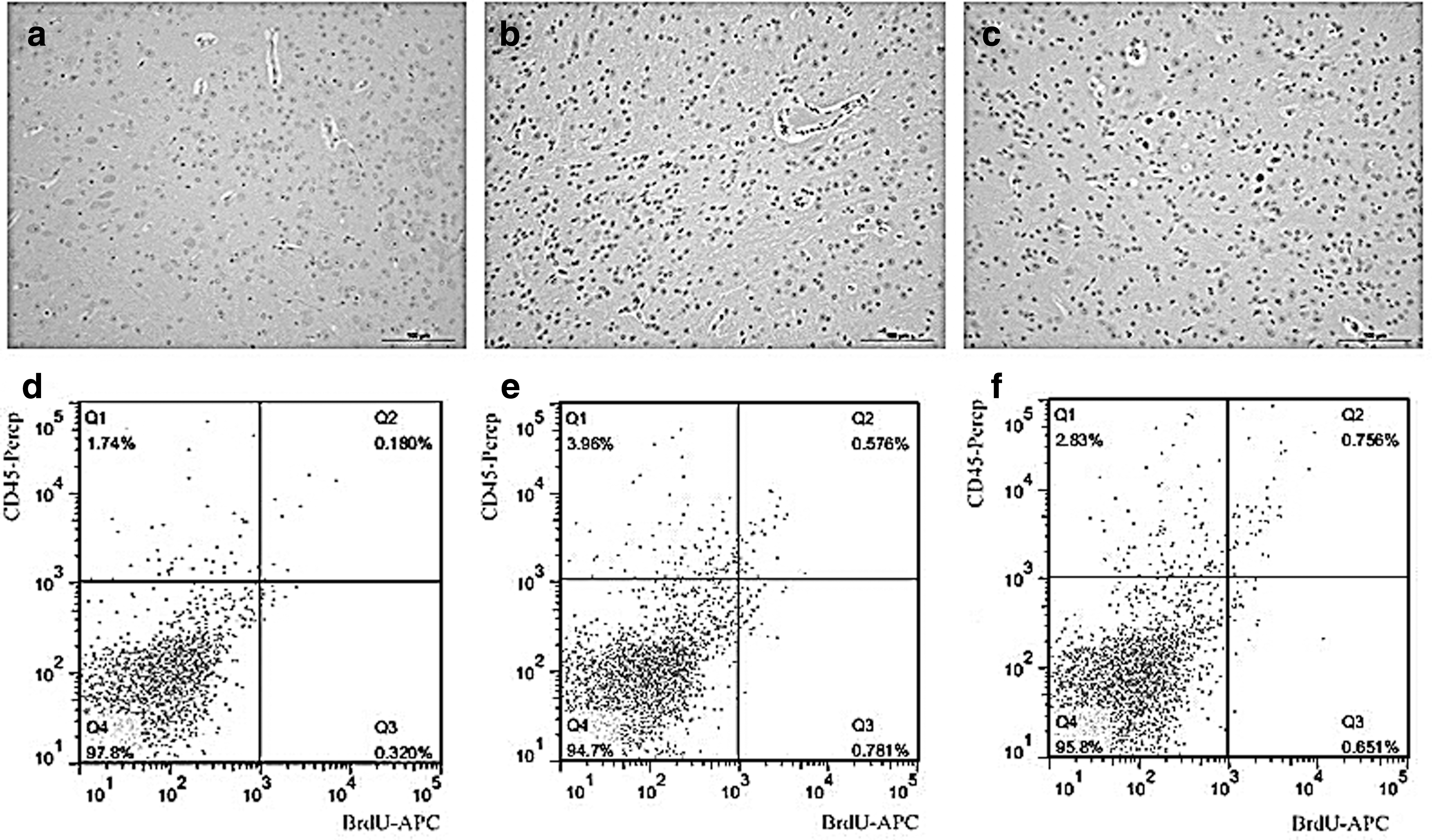
Microglia did not proliferate in situ. Sections were incubated with an anti-BrdU antibody using IHC staining.
Microglial cells originated from bone marrow-derived monocytes
Gr1 is expressed in the bone marrow-derived cells, and it may contain Ly6G or Ly6C antigens. Ly6G antigen is expressed by granulocytes, while Ly6C is expressed by monocytes and their precursors. We observed that the number of CD45high/Gr1+ expressing cells was 7.4% higher in PRV-infected brain samples than in the control samples (Fig. 6a, b). This demonstrated that the microglial cells observed in the brain lesions were bone marrow derived. In addition, the obtained results showed that there was no difference in the number of CD45high/Ly6Ghigh cells between the control and PRV-infected brains (Fig. 6c, d). The number of CD45high/Ly6Chigh cells increased about 24% in the infected brain compared with the control. Furthermore, the number of Ly6Chigh cells increased at 5 dpi, reaching significant difference in comparison with the control at 6 and 7 dpi (Fig. 6e–g).
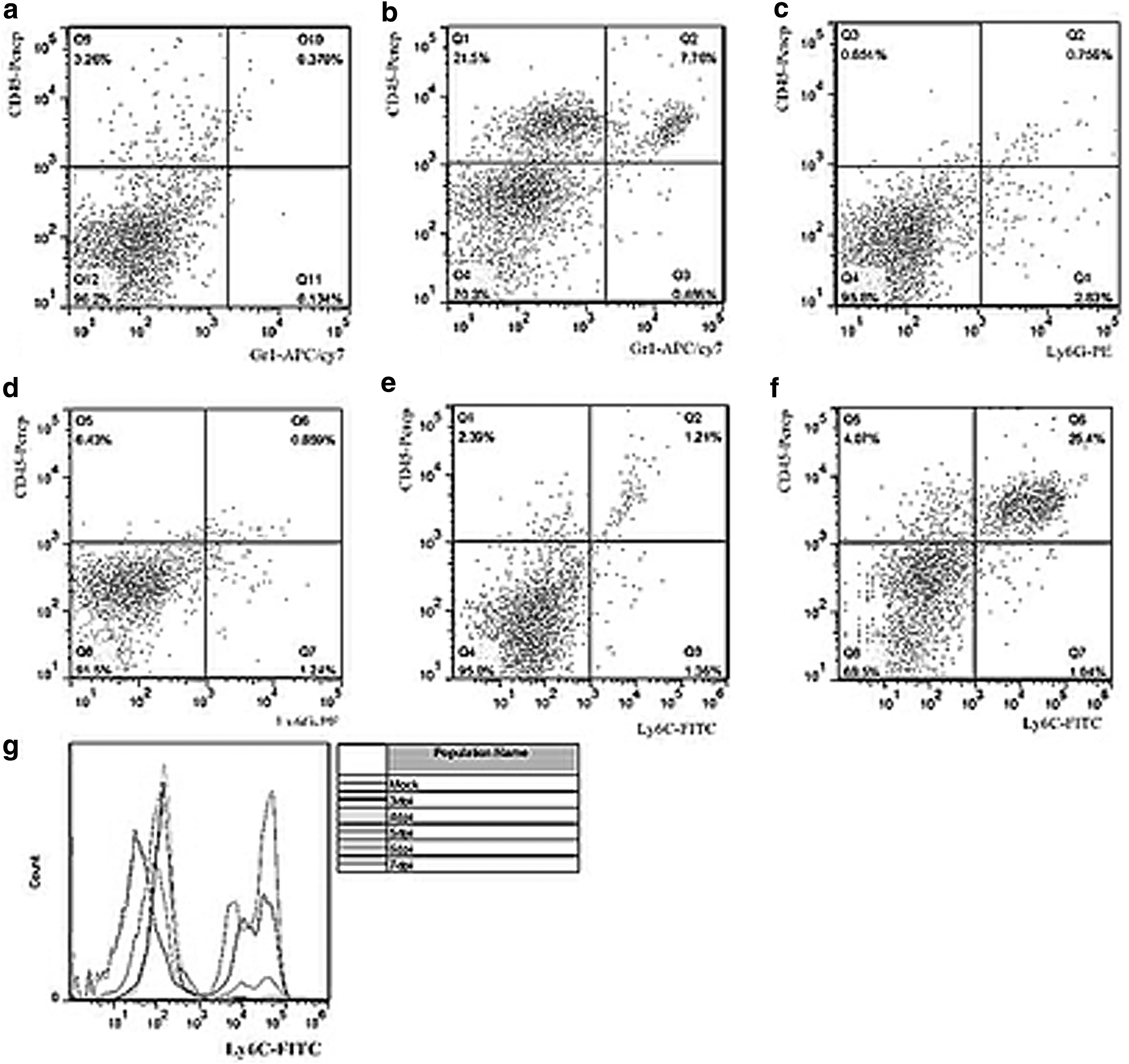
Monocyte precursors led to an increase in microglial cell numbers. Cell phenotypes in CD11b+ microglial cells were determined by flow cytometry. The microglia isolated from the control and PRV-infected mouse brains were labeled with CD45 and Gr1, and expressed in the bone marrow-derived cells
Microglial polarization
CD11b+ cells were isolated from the PRV-infected mouse brains. We analyzed the alterations in microglia-produced cytokines. The obtained results showed that the expression of both M1 and M2 markers gradually increased. The changes in M1 cytokine production were more pronounced than those of M2 cytokines. The expression of M1 cytokines TNF-α, iNOS, IL-6, and IL-12 was observed at 4 dpi, and the expression levels of these cytokines increased at 5 to 7 dpi, which was not observed in the control groups. The expression of M2 cytokines YM1 and IL-10 exhibited similar changes, but FIZZ1 expression levels did not change in comparison with the control (Fig. 7).
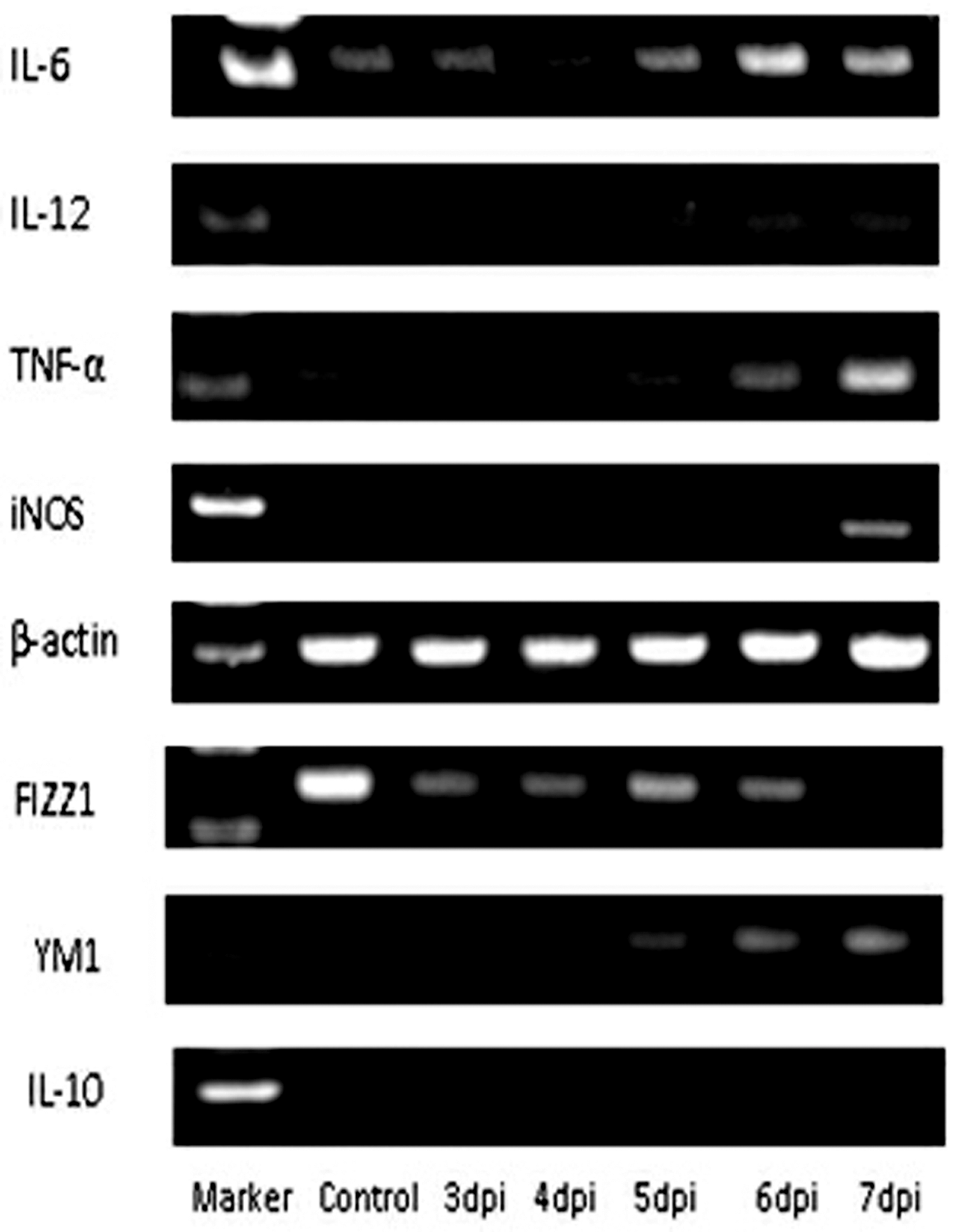
Microglial polarization occurred after PRV inoculation. The expression of M1 and M2 markers was analyzed by semiquantitative polymerase chain reaction at 3–7 days following PRV infection. The markers of M1-type macrophage include IL-6, IL-12, TNF-α, and iNOS, while the markers of M2-type macrophage include FIZZ1, YM1, and IL-10. β-actin was used as an internal gene.
Discussion
Although the natural hosts of PRV are adult swine, PRV can reportedly infect most livestock and wild animals. Clinical manifestations of the infection depend mostly on the virulence of the virus strain and the age of the affected pig. Common clinical symptoms of wild-type PRV infection include respiratory symptoms and growth retardation in growing pigs, reproductive failure in adult swine, and a fatal infection in suckling pigs (22,26). Several lines of evidence indicate that peripheral blood mononuclear cells, B lymphocytes, and T lymphocytes support PRV replication with different percentages of infection (23). Among the non-native hosts, rodents such as Balb/c mice have been widely used as animal models to study the pathogenesis of PRV infection in previous reports, including ours (36,37). Infection of the PRV virulent strain (PRV-HeN1) is lethal in neonatal piglets and in non-native hosts such as rodents with no identifiable symptoms of the CNS. In contrast, animals infected with an attenuated PRV vaccine strain (PRV-Bartha, used to protect swine from PRV for decades in China) exhibit longer survival time than wild-type PRV-infected animals with severe CNS symptoms. In this study, we used attenuated PRV strain, PRV-Bartha K61, because its lethality is lower than that of the virulent strain (1,19,37). The application of three different infection doses showed that the mortality of the treated mice depends on the applied infection dose. Since the survival rate of more than 50% and severe brain lesions were observed in the 1 × 105 TCID50-inoculated mouse group, we selected this dose for further experiments.
A previous study conducted to determine two distinct modes of neuroinvasion and lethality after murine flank inoculation with virulent (PRV-Becker) and attenuated strains (PRV-Bartha) of PRV revealed that mice infected with virulent (PRV-Becker) strains exhibit characteristic lesions and pruritus, die rapidly with no detectable CNS infection symptoms such as behavioral abnormalities, and have few viral antigens in the brain. In marked contrast, mice infected with an attenuated PRV vaccine strain (PRV-Bartha) have a longer survival time than wild-type PRV-infected animals, exhibit severe CNS abnormalities but absolutely no skin lesions, and have several viral antigens in the brain (3). These results suggest that PRV infection and transport in nerves in mice infected with PRV are associated with the virulence of the infecting strain; thus, virulent PRV (PRV-Becker) can enter the CNS by efferent and afferent routes, whereas the attenuated PRV vaccine strain (PRV-Bartha) enters exclusively by efferent routes. In this study, PRV infection and transport in cells from blood, routes by which the strains enter the brain, were determined. Using BrdU tracing technology, we showed that microglia did not proliferate in situ, suggesting that the blood precursors may regulate microglial numbers in brain lesions. Peripheral blood contains two monocyte subsets, inflammatory and circulating monocytes. Inflammatory monocytes can be distinguished from circulating monocytes by the differential expression of the Ly6C antigen, which forms a part of the Gr1 complex (12,13,35). In this study, the expression of Gr1 and Ly6C was shown to be upregulated in the infected brain compared with the control, but Ly6G expression did not change, suggesting that the microglia were derived from the inflammatory monocytes (9,14). Recruited monocytes can also contribute to the pathogenesis of infection and the development of chronic inflammatory diseases.
Microglial cells, belonging to the monocyte/macrophage lineage, are important brain tissue-resident professional phagocytes and APC that play a central role in inflammation and host defense (5). Macrophages display remarkable plasticity, polarized to both proinflammatory M1 and anti-inflammatory M2 phenotypes, and can change their physiology in response to environmental signals, for example, virus infection (10,21,25). The interactions between PRV and microglia were analyzed using phenotype-specific markers of mice macrophage polarization. The results showed that the expression of both M1 and M2 markers gradually increased, but the changes in M1 cytokine production were more pronounced than those of M2 cytokines. These suggested that PRV infection was shown to induce the proliferation and activation of microglia, mainly polarized into M1 phenotypes. Polarized M1-type macrophages have both beneficial and detrimental roles in several diseases. The protective role of M1 macrophages, which are related to the control of infection, is beneficial in many diseases caused by Listeria monocytogenes (32). However, an excessive or prolonged M1 activation is deleterious for the host during many diseases caused by Escherichia coli and Streptococcus sp. (24). However, further studies are needed to elucidate the role of microglial cell polarization in modulating PRV infection.
Footnotes
Acknowledgment
This study was supported by grants from the Chinese Academy of Agricultural Sciences fund for basic scientific research (2008041).
Author Disclosure Statement
The authors declare no conflicts of interest.