
Abstract
microRNAs have been reported to play crucial roles in various biological processes, including cell proliferation, apoptosis, tumor genesis, and viral infections. miR-26b has been found to be involved in the pathogenesis of multiple tumors, however, little is known about the role it plays in innate immune responses. In this study, we report that miR-26b is able to induce type-I interferon (IFN) expression, which was supported by both quantitative real time polymerase chain reaction and luciferase reporter assays. Conversely, production of IFN was reduced upon inhibition of miR-26b. Sequentially, ectopic expression of miR-26b led to upregulated expression of STAT1 and IFN-stimulated genes (ISGs). Furthermore, overexpression of miR-26b repressed the replication of vesicular stomatitis virus (VSV) and Sendai virus (SeV). In turn, IFN was able to induce the expression of miR-26b in a time-dependent manner. In all, we found that miR-26b could inhibit VSV replication through upregulation of type-I IFNs and ISGs and could in turn be upregulated by IFNs.
Introduction
M
Type-I interferon (IFN-α/β) is known for its antiviral activity, which plays critical roles in regulating the immune system against various viral infections (1,13). By binding to its receptors (IFN-AR1 and IFN-AR2), IFN-α/β can induce the tyrosine phosphorylation of the JAK family (JAK1 and Tyk2), followed by the activation of signal transducers and activators of transcription (STAT1 and STAT2) (1,15). The activated STATs then translocate to the nucleus, inducing the expression of interferon-stimulated genes (ISGs), such as IFIT1 and MX1 (13,23), which execute a series of biological functions, including antiviral activity.
In this report, we discovered that miR-26b significantly enhanced the activity of IFN signaling. Expression of IFNs, STAT1, and ISGs all increased upon ectopic expression of miR-26b mimics. As a result, miR-26b greatly inhibited the replication of vesicular stomatitis virus (VSV) and Sendai virus (SeV), two widely used viral models to assess the antiviral activity of IFNs (4,12). In turn, IFN stimulated the expression of miR-26b in a dose-dependent manner. This positive feedback may enhance the antiviral activity of miR-26b. As miRNAs are small molecules of no antigenic ability, miR-26b may become a potential effective antiviral product.
Materials and Methods
Cell culture and transfection
Human HEK293T cells were cultured in DMEM (HyClone) supplemented with 10% FBS (GIBCO, Life Technologies), glutamine, and antibiotics (100 U/mL penicillin, and 100 μg/mL streptomycin), at 37°C and 5% CO2. The miR-26b mimic, inhibitor, and the nonspecific controls, were all purchased from GenePharma. The sequence of miR-26b mimic and inhibitor are as following: 5′-UUCAAGUAAUUCAGGAUAGGU CUAUCCUGAAUUACUUGAAUU-3′, 5′-ACCUAUCCUGAAUUACUUGAA-3′. A total of 100 nM mimic or inhibitor of miR-26b was used. In dose-dependent assays, 0, 25, 50, or 100 nM mimic of miR-26b mimic was used. miR-26b mimic or inhibitor was transfected into 293T, A549, or 2fTGH cells using Lipofectamine 3000 (Invitrogen) according to the manufacturer's instruction. Cells were transfected for 48 h and then lysed for protein or RNA.
Western blots
Cells were lysed in NP-40 lysis buffer (50 mM Tris-HCl, pH 7.4, 150 mM NaCl, 1% NP-40, 1 mM EDTA, 10 mM N-ethylmaleimide, 50 μg/mL PMSF) supplemented with protease inhibitor mixture (Sigma) before being subjected to SDS-PAGE. Samples transferred to polyvinylidene fluoride (PVDF) membranes (Millipore) were immunostained with the following primary antibodies: anti-STAT1 (CST, 9172S) and anti-VSVG (Santa Cruz, sc-66180). The PVDF membrane was then subjected to secondary antibodies: HRP-conjugated goat anti-rabbit or goat anti-mouse (Bioworld), followed by detection with ECL-Prime (Thermo Scientific).
Analysis of mRNA expression
Total RNAs were extracted from Human HEK293T cells using TRIzol reagent (Invitrogen). Two micrograms of RNA was used for cDNA synthesis with random primers (Thermo) and the specific miR-26 RT primer with the sequence as following: 5′-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGGAGCCAAG-3′. mRNA level was determined by quantitative real time polymerase chain reaction (qRT-PCR) with specific primers. Human GAPDH gene was used as an internal control. SYBR Green Supermix (BIO-RAD) was used and samples were examined on an ABI Step One Plus Sequence Detection System. The primer sequences are as following: miR-26b (5′-ACACTCCAGCTGGGCCTGTTCTCCATTA-3′ and 5′-CTCAACTGGTGTCGTGGAGTCGGCA-3′), IFN-α (5′-TGGGAACAGAGCCTCCTAGA-3′ and 5′-CAGGCACAAGGGCTGTATTT-3′), IFN-β (5′-CATTACCTGAAGGCCAAGGA-3′ and 5′-CAGCATCTGCTGGTTGAAGA-3′), IFIT1 (5′-GCCTATCGCCAAGATTTAGATGA-3′ and 5′-TTCTGGATTTAACCGGACAGC-3′), and MX1 (5′-CGGACTGCGCCAAGATGTT-3′ and 5′-TTGACTCGCATCAGGAAGCG-3′).
Luciferase reporter assay
Luciferase reporter assays were performed in human HEK293T cells. Cells were plated in a 24-well plate 20–24 h before transfection. P125-LUC or ISRE-LUC plasmid was then cotransfected with miR-26 mimic or inhibitor or the negative controls on the following day. Forty-eight hours after transfection, cells were lysed and the relative luciferase activity was measured according to the manufacturer's instructions.
Statistical analysis
All data were representative for three independent experiments and were presented as mean ± standard deviations. Statistical significance was determined by the two-tailed Student's t-test. p < 0.05 was deemed statistically significant.
Results
miR-26b induces the expression of type-I IFNs
The role of miR-26b in multiple tumors is widely studied by many groups. However, researches regarding the function of miR-26b in regulating IFN signaling and viral infections are relatively few. To explore how miR-26b regulates the expression of type-I IFNs, we transfected mimics or inhibitor of miR-26b in HEK293T cells and detected the expression levels of IFNs. As shown in Figure 1, the mRNA level of IFN-α significantly increased along with the ectopic expression of miR-26b mimics in HEK293T. To confirm the effects of miR-26b on IFNs expression, we used different doses of miR-26b. Results showed that IFN-β expression was induced by miR-26b in a dose-dependent manner (Fig. 1C). Similar results were achieved in 2fTGH cells (Supplementary Fig. S1; Supplementary Data are available online at
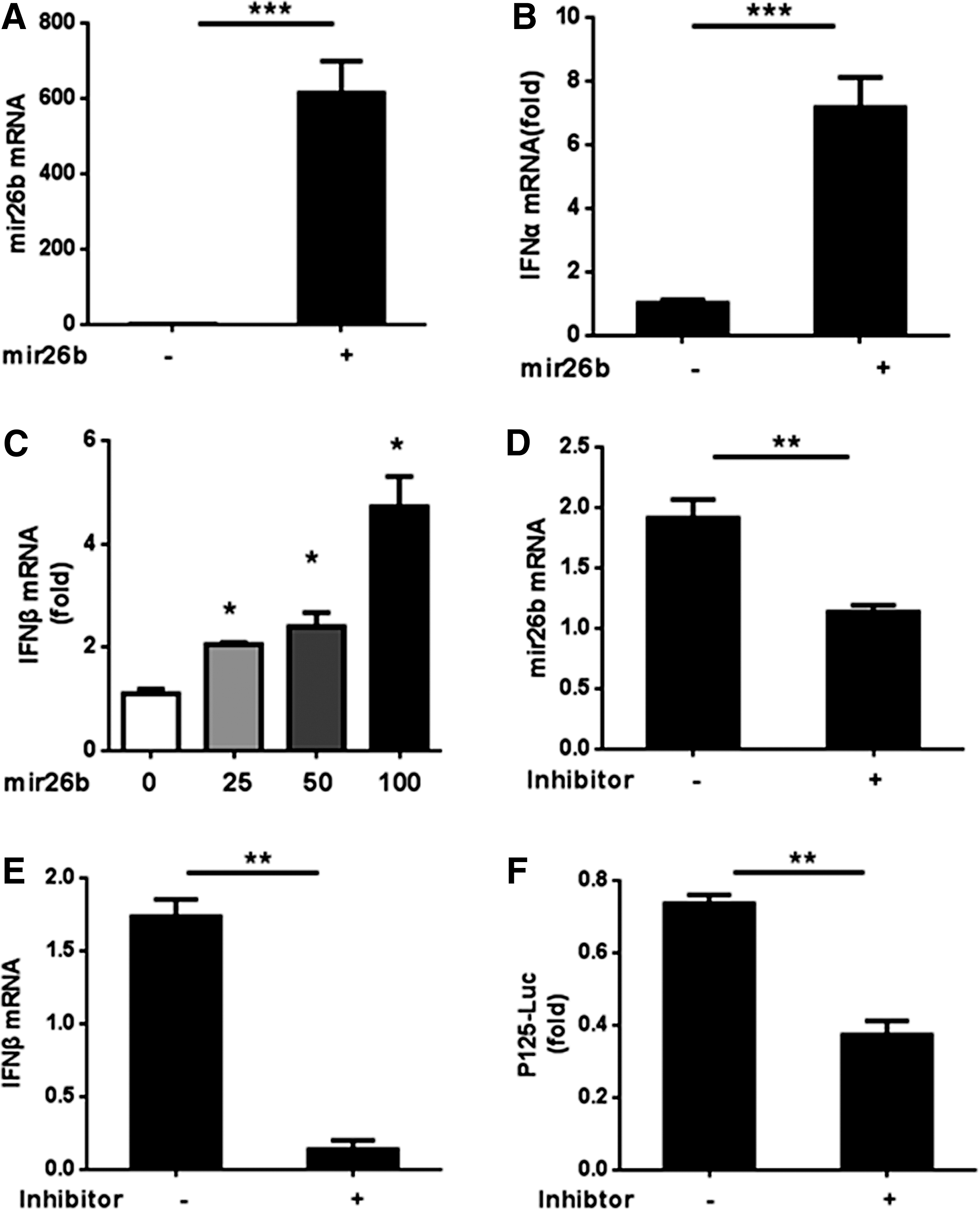
miR-26b induces the expression of type-I IFNs.
miR-26b upregulates the expression of ISGs
It is well known that the JAK-STAT signaling can be induced by type-I IFNs (1). As we had confirmed that miR-26b could upregulate the expression of type-I IFNs, we supposed that miR-26b might be able to induce the expression of STAT1 and ISGs (IFN-induced genes). In an attempt to verify this hypothesis, we first examined the mRNA and protein level of STAT1 after transfecting mimics of miR-26b into HEK293T cells. Both the mRNA and protein levels were upregulated by mimics of miR-26b (Fig. 2A–C).Then, we sought to verify the effects of miR-26b on the expression of ISGs. On the one hand, we transfected the ISRE-LUC reporter into HEK293T cells together with the inhibitor or mimics of miR-26b or the relative controls. The analysis of the luciferase reporter assays demonstrated that inhibition of miR-26b led to a significant decrease in the luciferase activity of ISRE-LUC; whereas on the contrary, introduction of mimics of miR-26b in the HEK293T cells resulted in an increase of the luciferase activity (Fig. 2D, E). On the other hand, we detected the mRNA expression of the ISGs, including IFIT1 and MX1, after transfecting the mimics or inhibitor of miR-26b for 48 h. mRNA level of both IFIT1 and MX1 increased along with the overexpression of miR-26b, whereas decreased following the inhibition of it (Fig. 2F–I). Taken together, these results demonstrated that miR-26b could upregulate the expression of STAT1 and ISGs in HEK293T cells.
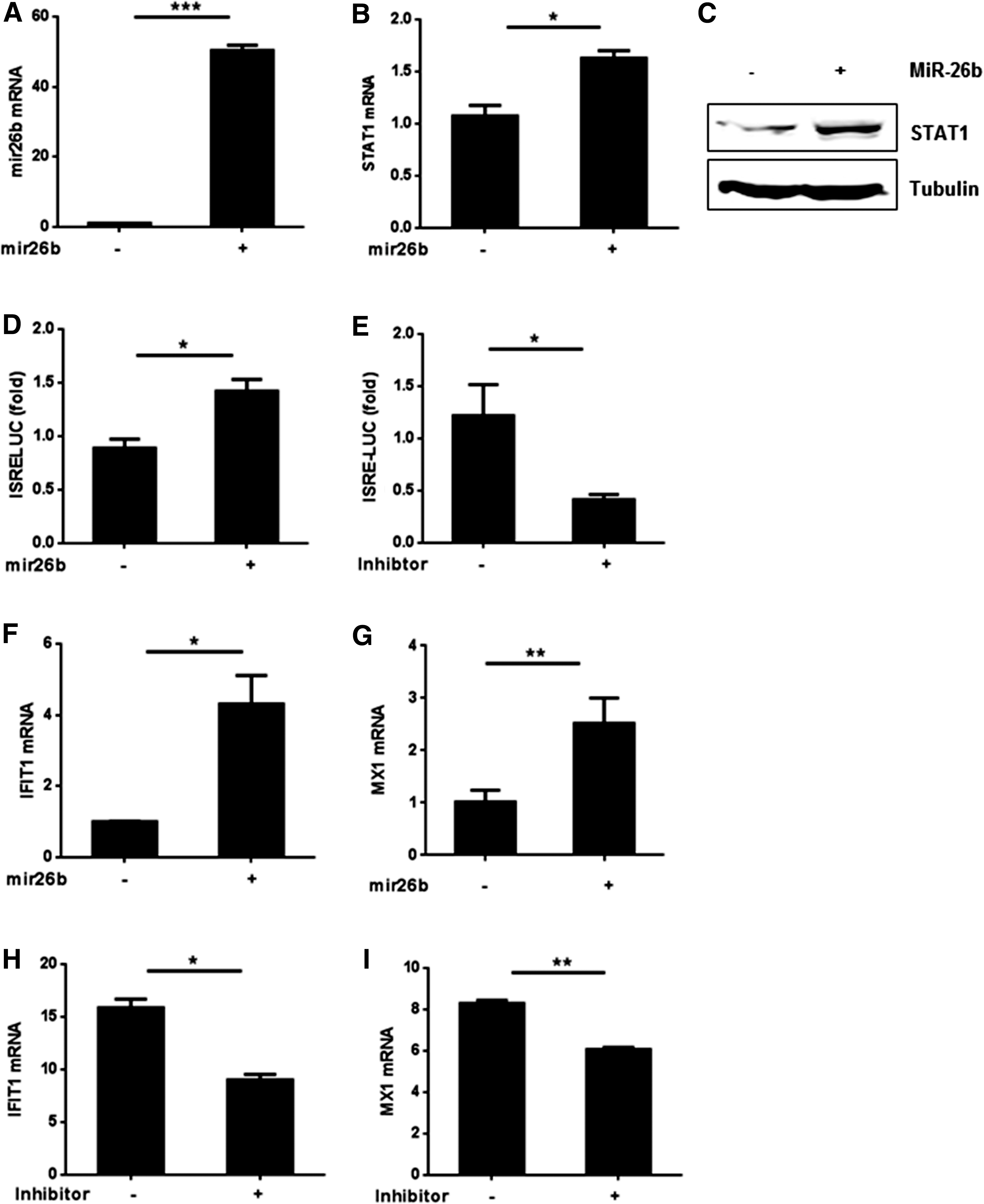
miR-26b upregulates the expression of IFN-stimulated genes.
miR-26b represses the replication of SeV and VSV
VSV and SeV are widely used viral models to evaluate the antiviral ability of IFNs. Therefore, we used these two viruses to examine whether miR-26b could inhibit viral replication through inducing the expression of type-I IFNs and ISGs. VSVG is one of the major components of the VSV virus and is often used as a marker protein for VSV. Overexpression of miR-26b mimics reduced the expression of VSVG both in the mRNA and protein levels (Fig. 3A–C). On the contrary, transfecting miR-26b inhibitor into HEK293T cells infected with VSV led to a rise in the transcription level of VSVG (Fig. 3D, E). Similarly, overexpression of miR-26b inhibitor in HEK293T cells led to a ∼1.6-fold upregulation of SeV mRNA replication (Fig. 3F). Besides, miR-26b also repressed VSV or SeV replication in 2fTGH or A549 cells that showed this effect was not limited to one cell line (Supplementary Fig. S2). Taken together, our results indicated that miR-26b could play a role in repressing virus replication.

miR-26b represses the replication of SeV and VSV.
miR-26b expression is upregulated by IFN in turn
We next asked whether IFNs could regulate the expression of miR-26b. IFN-α was added to the medium of HEK293T cells for 0, 4, 7, or 10 h, and then the relative expression level of miR-26b was analyzed. As shown in Figure 4A, mRNA level of miR-26b increased along with the addition of IFNs. Moreover, the longer the stimulus lasts, the more obvious the increase. Similar results were achieved when IFN-α was replaced by IFN-β. Therefore, we got the conclusion that type-I IFNs could in turn upregulate the expression of miR-26b in a time-dependent manner.
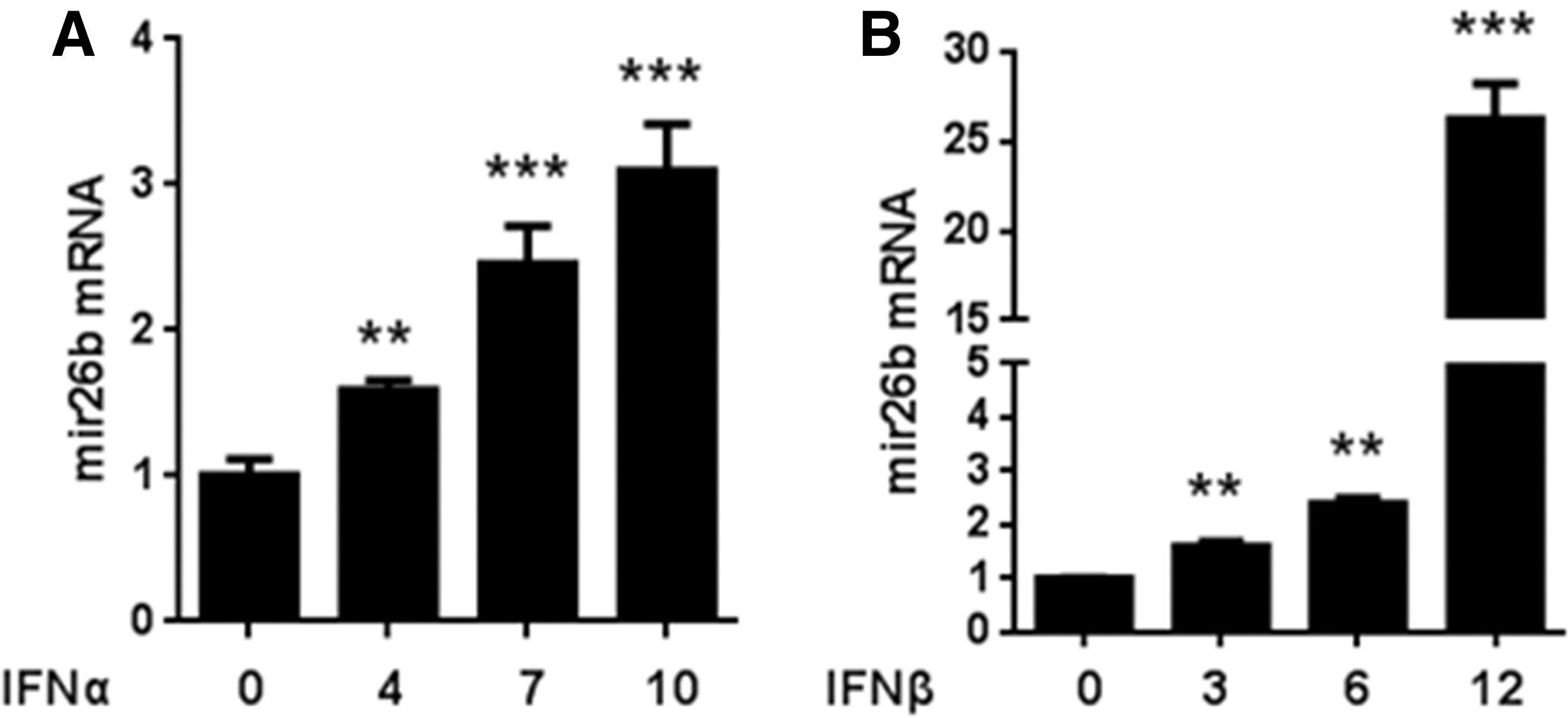
miR-26b expression is upregulated by IFN in turn.
Discussion
The miR-26 family, including miR-26b, has been revealed to play important roles in tumor progression and cardiac diseases (6,8). While in recent years, a few researches also demonstrated the role of miR-26b in inflammation through regulating the NF-κB signaling or IL-6 (2). However, none of them was focused on the regulation of IFNs. In this report, we, for the first time, demonstrated that miR-26b could repress the replication of viruses through inducing the expression of type-I IFNs.
First of all, we demonstrated that expression of type-I IFNs, including IFN-α and IFN-β in HEK293T cells could be upregulated by miR-26b mimics, whereas downregulated by miR-26b inhibitor (Fig. 1). However, we failed in finding a direct target of miR-26b, which could act as an intermediate role between miR-26 and IFNs. STAT1 is a predicted target of miR-26b. However, in our report, we found that as an ISG, STAT1 was upregulated by miR-26b mimics. Other ISGs, including IFIT1 and MX1, which could be induced by type-I IFNs, could also be upregulated by miR-26b mimics (Fig. 2). Furthermore, we found that replication of two viral models, VSV and SeV, could both be repressed by miR-26b mimics (Fig. 3). This was probably owing to the antiviral effect of IFNs induced by miR-26b. Intriguingly, we also discovered that miR-26b could in turn be upregulated by type-I IFNs in a time-dependent manner (Fig. 4). This makes a positive feedback relationship between miR-26b and type-I IFNs and may enhance the antiviral activity of miR-26b.
In summary, miR-26b inhibits virus replication through upregulating expression of type-I IFNs and ISGs and can in turn be upregulated by IFNs. Although finding the direct target of miR-26b, which may act as a regulator of IFNs, remains an unsolved problem. Our report provides evidence that miR-26b may play a significant role in the defense of viral infections.
Footnotes
Acknowledgments
This research was supported by grants from the National Natural Science Foundation of China (81602472, 31370873, and 31570865), the Program of 1000 Young Talents, and Jiangsu Provincial Distinguished Young Scholars (BK20130004). The authors thank the program for Changjiang scholars and Innovative Research Team in the University of Ministry of Education of China (PCSIRT-IRT1075), the Jiangsu Provincial Innovative Research Team, and a project funded by the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD).
Author Disclosure Statement
No competing financial interests exist.