
Abstract
During respiratory viral infection, conventional dendritic cells (cDCs) take up antigen and migrate to the draining lymph nodes to present viral antigen and activate cytotoxic T lymphocytes; however, regulation of cDC activation and migration may be age dependent. In this study, we used a mouse model of paramyxoviral infection (Sendai virus) and demonstrated that cDCs, which have migrated from lungs to the draining lymph nodes, are delayed in expressing activation markers in neonatal mice compared with adults. Neonatal lung cDCs expressed reduced levels of MHC Class II (major histocompatibility complex II) and CCR7 (chemokine receptor type 7) on postinfection days 3 and 5, respectively. The level of the CCR7 ligand CCL19 was significantly reduced in neonatal lungs during the course of viral infection. Interestingly, the arachidonic acid metabolite prostaglandin D2 (PGD2) was present at significantly higher levels in neonatal bronchoalveolar lavage fluid compared with adults. This was associated with increased expression of lipocalin PGD2 synthase mRNA levels in neonatal lungs and in isolated neonatal tracheal epithelial cells. Although thymic stromal lymphopoietin (TSLP) expression has been associated with increased PGD2 production, we found that TSLP levels were reduced in neonatal lungs. Importantly, blocking PGD2 function using a prostaglandin D2 receptor 1 (DP1) antagonist restored cDC activation in neonates. Together, these data suggest that cDC activation in neonates is delayed by a PGD2 mechanism and associated decreased chemokine signals.
Introduction
C
We previously demonstrated that neonatal mice have reduced inflammation during the innate immune response to the paramyxovirus Sendai virus (SeV). During infection, adult mice exhibited body weight loss that correlated with the level of inflammation in the lungs (typified by a neutrophilic influx and increased expression of interferon γ [IFN-γ], interleukin 1 [IL-1β], IL-6, and tumor necrosis factor [TNF-α]). In contrast, SeV-infected neonatal mice did not lose weight during infection. They exhibited a growth pattern similar to uninfected neonates; this correlated with an absence of inflammatory cell influx and reduced expression of IFN-γ, IL-1β, IL-6, and TNF-α in the lungs. Despite the absence of inflammation, the neonatal mice were able to inhibit viral replication and eliminate virus in a manner similar to adult mice (1). This observation that neonates were fully capable of viral clearance was in contrast to studies demonstrating fewer cDCs with a reduced capacity for antigen presentation and IFN-γ production in neonatal spleen, thymus, and lymph nodes compared with adults (3). Thus, we wanted to determine the contributions of cDCs to the neonatal response to SeV infection.
In this study, we demonstrated for the first time that the activation of CD103+ cDCs was delayed in neonatal mice. A delay was observed in the expression of activation markers MHC Class II and CCR7 on neonatal CD103+ cDCs. In addition, the expression of the CCR7 ligand CCL19 was reduced in neonatal lungs. Interestingly, the level of prostaglandin D2 (PGD2) was significantly increased in neonatal bronchoalveolar lavage fluid, and this was associated with increased expression of lipocalin-type PGD2 synthase in isolated airway epithelial cells. Treatment of neonatal mice with PGD2 DP1 receptor antagonist BWA868C at the time of infection was able to prevent the delay of CD103+ cDC activation in neonates, suggesting a potential regulatory role for PGD2 in the neonatal lung during viral infection.
Materials and Methods
Mice and viral infection
Wild-type C57BL/6 mice were purchased from Jackson Laboratories. Mice were housed in pathogen-free environment in a biohazard facility in microisolator cages. All procedures were reviewed and approved by the Saint Louis University Animal Studies Committee. Adult 6- to 8-week-old C57BL/6 mice were anesthetized and infected intranasally with 500 pfu/g body weight of SeV (Strain 52; American Type Culture Collection) in 30 μL phosphate-buffered saline (PBS). Similarly, 2-day-old neonatal mice were infected intranasally with 500 pfu/g body weight of SeV in 6 μL of PBS as described (1). Mice were monitored daily for body weight loss and general health. To track cells migrating from the lungs to the lymph nodes, mice were inoculated intranasally with Cell Proliferation Dye eFluor® 450 (eBioscience, Inc., San Diego, CA) at the time of viral infection. To block PGD2 activity, neonatal mice were inoculated with PBS containing virus and 10 nmol of the DP1 receptor antagonist BWA868C (Cayman Chemical, Ann Arbor, MI) (29).
Enzyme-linked immunosorbent assay
Lungs were homogenized in 1% Nonidet P-40, 0.05 M Tris, pH 8.0, 250 mM NaCl, 1 mM ethylenediaminetetraacetic acid (EDTA) containing protease inhibitor cocktail (Sigma Aldrich, St. Louis, MO). A bicinchoninic acid assay (BCA; Thermo Scientific, Rockford, IL) was used to normalize protein concentrations. Thymic stromal lymphopoietin (TSLP) was measured using a Mouse TSLP enzyme-linked immunosorbent assay (ELISA) Max kit according to manufacturer's recommendations (Biolegend®, San Diego, CA).
Immunohistochemistry
Lungs were inflated with 10% neutral buffered formalin and fixed for 72 h. Five micrometers paraffin sections were incubated with rabbit anti-mouse TSLP antibody (Santa Cruz Biotechnology, Santa Cruz, CA) for 1 h at 25°C. Primary antibody binding was detected with biotinylated goat anti rabbit immunoglobulin G (Santa Cruz Biotechnology) for 30 min at 25°C and the VECTASTAIN ABC-AP Kit (Vector Laboratories). Sections were stained with an alkaline phosphatase red substrate and counterstained with hematoxylin.
PGD2 ELISA
Bronchoalveolar lavage (BAL) was performed by tracheal cannulation as described (23). Total protein concentration in BAL fluids was determined with the Pierce BCA protein assay kit (Thermo Scientific). Lipids were extracted and analyzed with a PGD2 MOX-ELISA kit according to manufacturer's instructions (Cayman Chemical). Prostaglandin levels were calculated and normalized to the total protein concentrations in the BAL fluid.
Flow cytometry
To obtain a single cell suspension, lungs were minced and placed in Dulbecco's modified Eagle medium (DMEM) containing hyaluronidase (5 U/mL), collagenase I (250 U/mL), and DNAse (50 U/mL; Worthington Biomedical Corporation) and incubated for 45 min in 37°C at 5% CO2, followed by incubation with EDTA (1 mM) for 15 min. Lung tissues were gently mashed between frosted slides and connective tissue was removed using cell strainers. Ammonium-chloride-potassium lysis buffer (Thermo Fisher Scientific, Waltham, MA) was used to lyse the erythrocytes followed by centrifugation at 1,200 rpm at 4°C for 7 min. Cells were incubated with fixable viability dye (eFluor) and Fc block, and then stained with: CD45-Per Cp/Cy5.5 (Biolegend), Lin cocktail-FITC (Biolegend), Siglec-F-Alexa Fluor 647 (BD Biosciences, San Jose, CA), CD103-Pacific Blue (Biolegend), CD11c-Alexa Fluor 700 (Biolegend), MHC Class II-IA-IE-PE (Biolegend), and CCR7-PE-Cy7 (eBioscience, Inc.). Viable and singlet cells were used for analysis and CD45+/Lin−/Siglec-F−/CD103+/CD11c+ cDCs were analyzed for MHC Class II and CCR7 expression (7,19). SeV-specific CD8+ T cells were stained with H-2Kb SeV Tetramer-FAPGNYPAL-PE (MBL International, Inc., Woburn, MA). Data were analyzed using FlowJo software (Tree Star, Inc., Ashland, OR).
Airway epithelial cell isolation
Airway epithelial cells were isolated as described. In brief, tracheas were dissected and incubated for 18 h at 4°C in DMEM containing 1.5 mg/mL pronase (23). The collection tube was gently inverted to loosen tracheal epithelial cells and to rinse the tracheas. Fetal bovine serum was added to achieve a final concentration of 5%. The epithelial cells were collected by centrifugation at 1,200 rpm at 4°C in a table top Sorvall RT600.
RNA isolation and quantitative PCR analysis
RNA was extracted using Illustra RNAspin Mini RNA Extraction Kit (GE Healthcare, Buckinghamshire, United Kingdom). cDNA was generated using High-Capacity Reverse Transcription Kit (Applied Biosystems, Carlsbad, CA). Quantitative real-time polymerase chain reaction (qPCR) was performed using the MJ Chromo 4 detection system (Bio-Rad Laboratories, Hercules, CA). SYBR Green master mix (Sigma Aldrich) was used for qPCR analysis. Disassociation curve analysis was performed to assess the specificity of amplification for each sample, and PCR product size was verified by agarose gel electrophoresis. Percent expression was normalized by GAPDH and analyzed using the comparative threshold cycle (Ct) method (18). CCL19: forward, 5′-GGG GTG CTA ATG ATG CGG AA-3′, reverse, 5′-CCT TAG TGT GGT GAA CAC AAC A-3′; CCL21: forward 5′-GTC CGA GGC TAT AGG AAG CA-3′, reverse, 5′-TTA GAG GTT CCC CGG TTC TT-3′; CXCL13: forward, 5′-CTC TCC AGG CCA CGG TAT T-3′, reverse, 5′-TAA CCA TTT GGC ACG AGG AT-3′; GAPDH: forward, 5′-AGA TTG TCA GCA ATG CAT CC-3′; reverse, 5′-ACA GTC TTC TGG GTG GCA GT-3′; Hpgds (hematopoietic PGD2 synthase): forward, 5′-GGA AGA GCC GAA ATT ATT CGC T-3′; reverse, 5′-ACC ACT GCA TCA GCT TGA CAT–3′; Ptgdr (DP1 receptor): forward, 5′–ATG AAC GAG TCC TAT CGC TGT-3′; reverse, 5′-ACA CGA GCA CAT AAA AGA CCG-3′; Ptgdr2 (DP2 receptor): forward, 5′-AGA TGG TCC AGC TTC CAA ACC-3′; reverse, 5′-ACA GGA TGA GTC CGT TTT CCA-3′; Ptgds (PGD2 synthase): forward, 5′-CAG GAA AAA CCA GTG TGA GAC CAA-3′; reverse, 5′-AGA GGG TGG CCA TGC GGA AG-3′. The primer sequences were obtained from PrimerBank (25 –28).
Statistical analysis
A two-tailed t-test was used to analyze mean values between groups. All values represent mean ± standard deviation. p-Values <0.05 were considered statistically significant.
Results
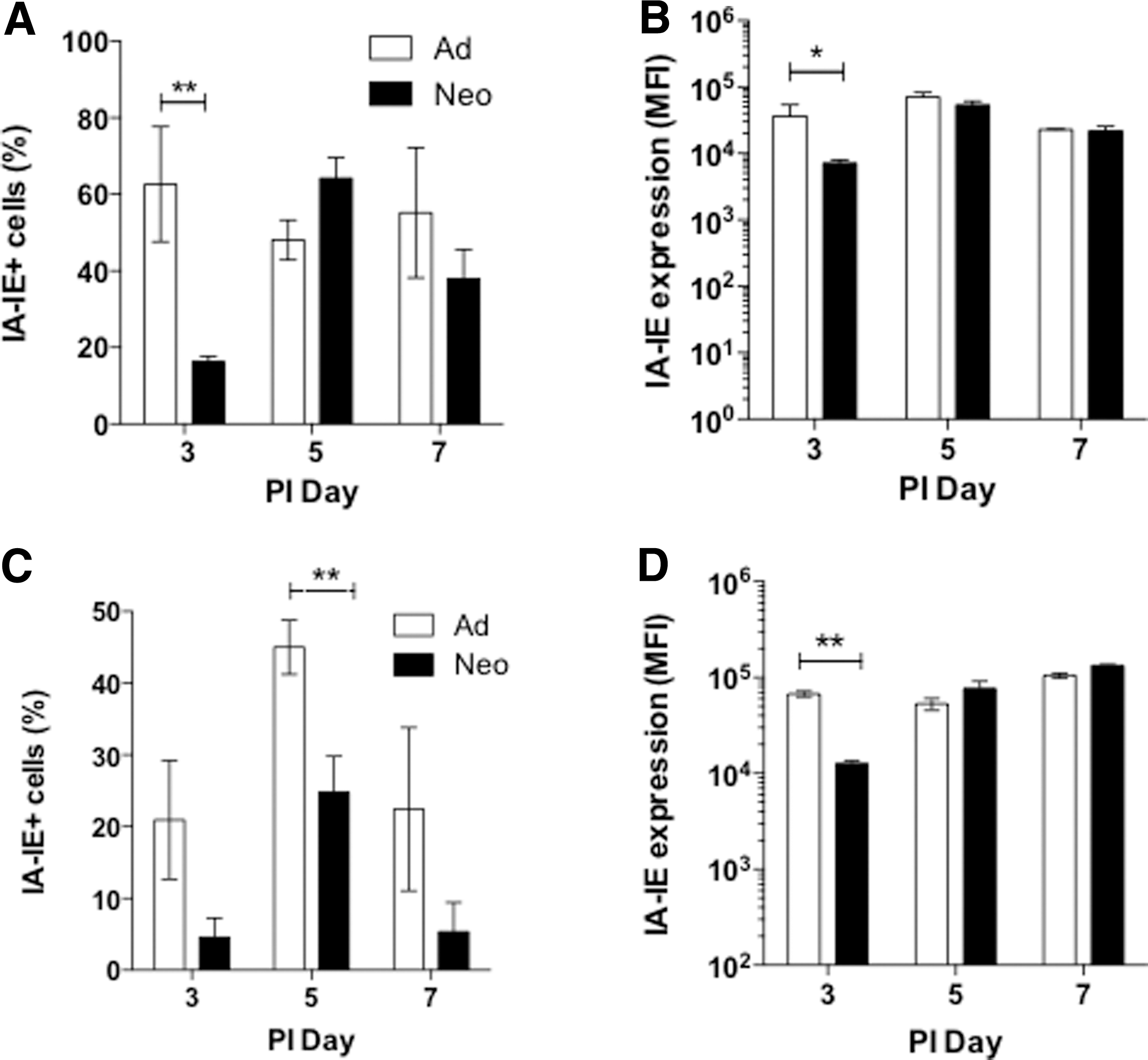
Adult and neonatal C57BL/6 mice were infected with SeV (500 pfu/g body weight) and cDC activation and migration were assessed in adult and neonatal lungs and lymph nodes. cDCs were defined as viable singlet cells that were CD45+/Lin−/Siglec−F−/CD103+/CD11c+ cells. A representative gating strategy of adult lymph nodes is given in Figure 1. In neonatal lungs, we detected a significantly reduced frequency of MHC Class II+ cDCs on postinfection day 3 compared with adults (Fig. 2A), and these cells also had reduced expression of MHC Class II compared with adult cells (Fig. 2B). In the neonatal lymph nodes, fewer MHC Class II+ cDCs were present on days 3 and 5 postinfection (Fig. 2C). In addition, there was significantly reduced MHC Class II expression on neonatal cDCs in lymph nodes on day 3 postinfection; however, by day 5 the expression level of MHC Class II on cDCs was similar in both adults and neonates (Fig. 2D).
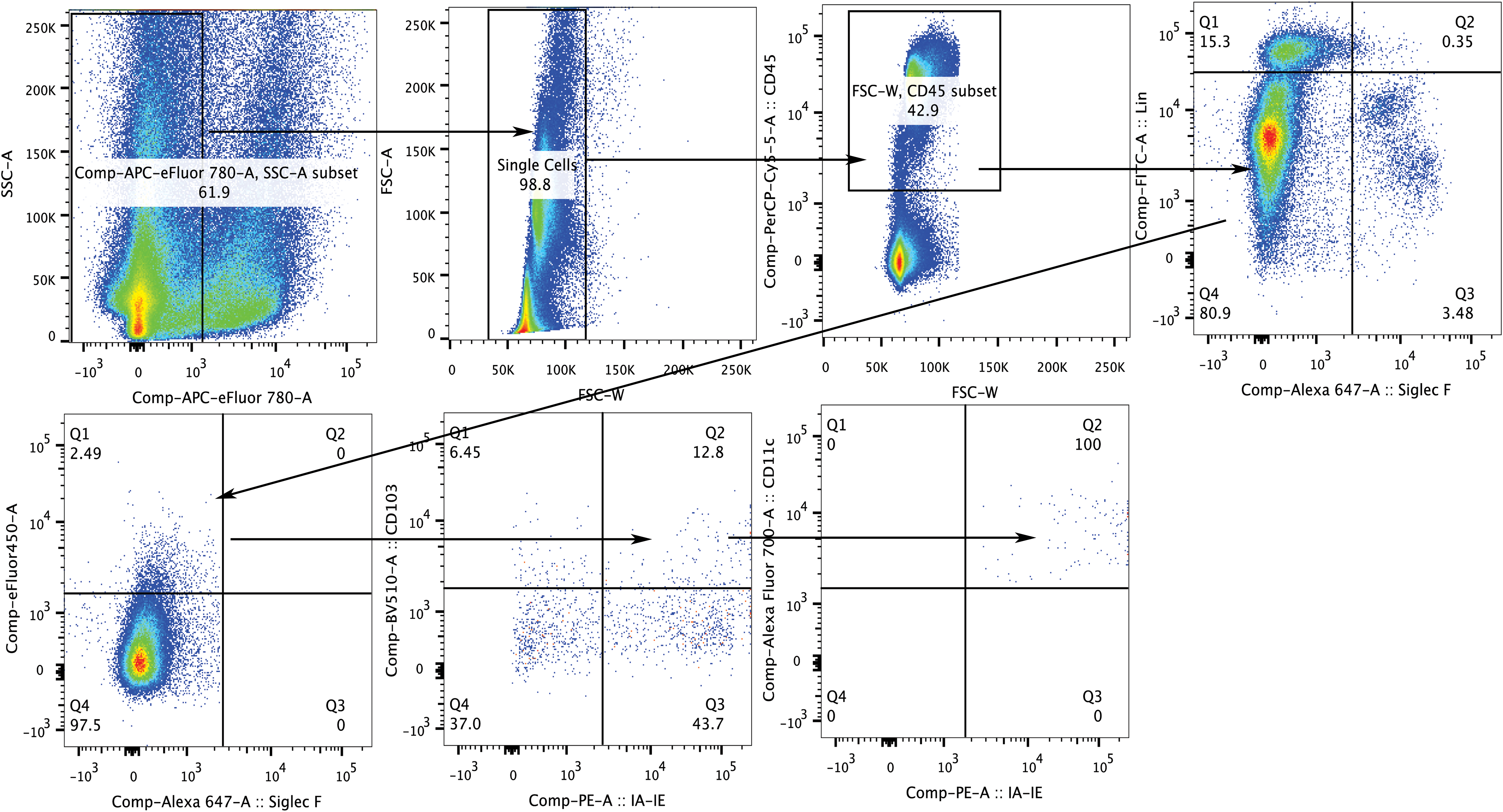
Representative gating strategy for cDC populations showing adult lymph nodes on postinfection day 3. Viable CD45+ single cell populations positive for intranasal cell proliferation dye were selected, followed by selection of lineage− and SiglecF− cells. The CD11c+/CD103+ cell population was assessed for MHC Class II expression. Arrows denote the gates that were analyzed in each step. cDCs, conventional dendritic cells; MHC Class II, major histocompatibility complex II. Color images available online at
Analysis of MHC Class II+ cDCs in lungs and lymph nodes in neonates and adults during the course of infection. Adults (white bars) and neonates (black bars) were infected intranasally with SeV (500 pfu/g body weight) and tissues were isolated on the indicated postinfection days. Percentage of MHC Class II+ cDCs (CD45+/CD103+/CD11c+) in lungs
Total cell numbers per gram of body weight were similar in both the lungs and lymph nodes of adult and neonatal mice (Fig. 3A). To track cDC migration from the lung to the lymph nodes, the Cell Proliferation Dye eFluor 450 was administered intranasally at the time of viral inoculation. Figure 3B shows that the average frequency of MHC Class II+/CD103+ cDCs of eFluor 450-labeled cells in neonatal draining lymph nodes was lower than in adult lymph nodes on day 3 postinfection. The figure shows the results from two independent experiments, and each data point represents the mean of one experiment. For each experiment, lymph nodes from two adult mice or six neonatal mice were pooled for analysis. In one experiment, eFluor 450-labeled cDCs were undetectable in the pooled neonatal lymph nodes.

Migration of MHC Class II+/CD103+eFluor+ cDCs to draining lymph nodes and CD8+ T lymphocyte analysis. Adult and neonatal C57BL/6 mice were inoculated intranasally with saline containing SeV (500 pfu/g body weight) and eFluor® 450 dye.
CD8+ T lymphocytes are critical for clearance of SeV. The frequency of CD8+ T lymphocytes was significantly reduced in neonatal lungs, but significantly increased in neonatal lymph nodes on postinfection day 11 (Fig. 3C). Analysis of total CD8+ cells per gram body weight showed a similar pattern in the lung and lymph nodes (Fig. 3E). To determine whether the delay in cDC activation affected SeV-specific CD8+ cytotoxic T lymphocytes, flow cytometry with the SeV-specific tetramer (FAPGNYPAL) was performed. Figure 3D and F show that the frequency and total numbers of SeV-Tet+/CD8+ cytotoxic T cells in the lungs and lymph nodes of adults and neonates were similar.
Dendritic cell migration is coordinated by chemokine gradients and chemokine receptor expression. Activated cDCs upregulate expression of CCR7 that facilitates their migration to the draining lymph nodes (14). Neonatal lung cDCs exhibited significantly reduced surface expression of CCR7 on postinfection day 5 compared with adults (Fig. 4A). There were no significant differences in the surface expression of CCR7 between adult and neonatal cDCs at any time point in the draining lymph nodes (Fig. 4B). The CCR7 ligands, CCL19 and CCL21, coordinate the migration of cDCs to the lymph nodes. During SeV infection, CCL19 expression increased in both adult and neonatal lungs, although the neonates expressed significantly lower levels of CCL19 compared with adults at all time points (Fig. 4C). In contrast, neonates showed significantly higher expression of CCL21 in postinfection days 3 and 7 lungs (Fig. 4D).
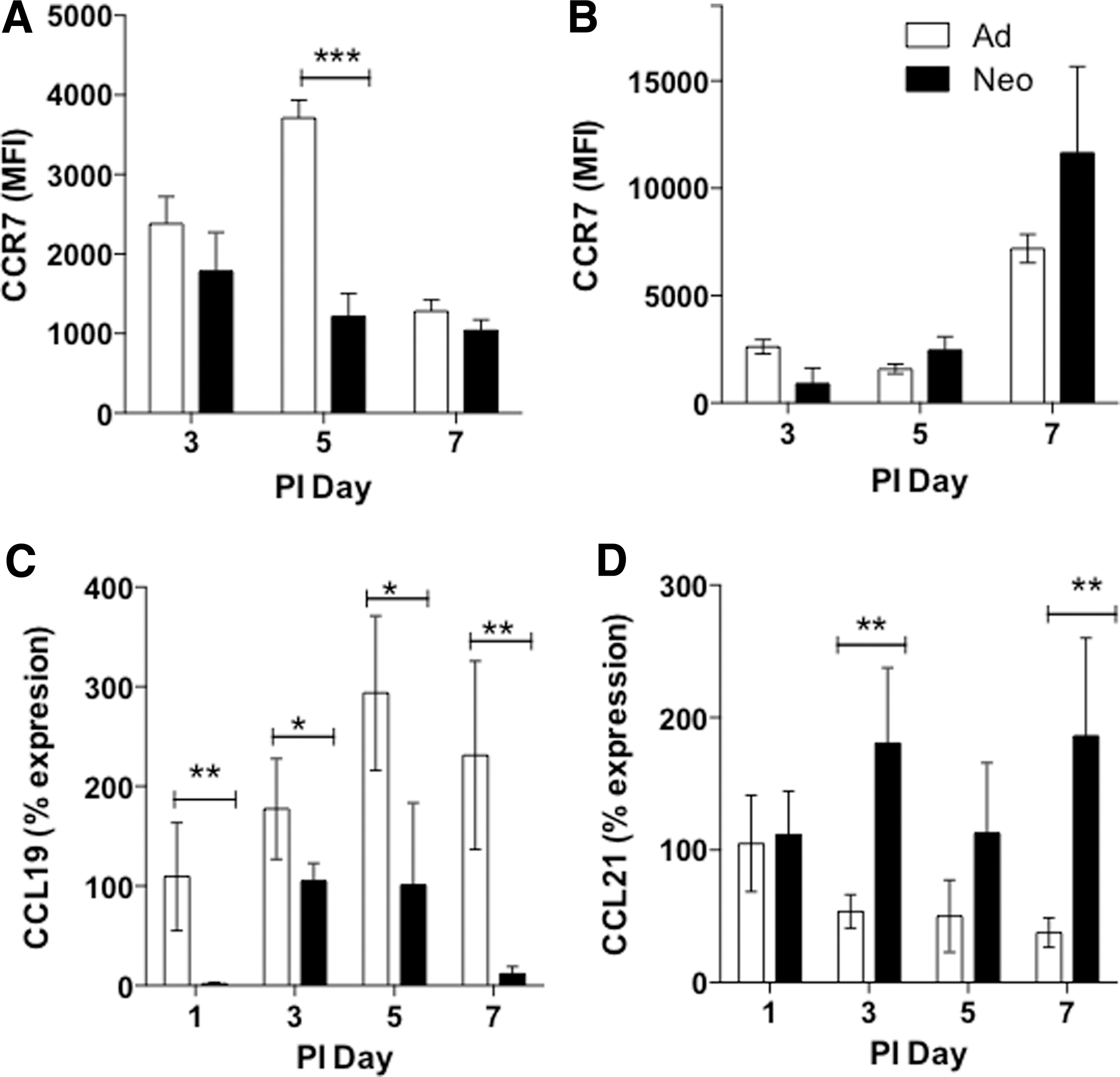
Chemokine receptor and ligand expression is altered in neonatal lungs during paramyxoviral infection. Adults (white bars) and neonates (black bars) were infected with SeV as described. Flow cytometry for CCR7 expression on CD45+/CD103+/CD11c+ cDCs in the lungs
Previous studies in geriatric mice showed that the arachidonic acid metabolite PGD2 delayed cDC migration during respiratory viral infections (29). Consequently, we assessed PGD2 levels in adult and neonatal BAL fluids during the course of SeV infection. Neonatal mice showed significantly higher levels of PGD2 compared with adults on postinfection days 2, 3, and 5 (Fig. 5A). We next tested the two enzymes that catalyze the conversion of PGH2 to PGD2: ptgds (lipocalin-type PGD2 synthase) and hpgds (hematopoietic PGD2 synthase) (13). Neonatal mice showed higher expression of ptgds mRNA in whole lungs compared with adults on days 0 and 1 (Fig. 5B). Because SeV primarily infects tracheal epithelial cells we also examined ptgds mRNA expression in primary isolated tracheal epithelial cells. The results showed a more pronounced increase in ptgds in the neonatal epithelial cells compared with adult cells (Fig. 5C). In contrast, there was no significant difference in expression of hematopoietic PGD2 synthase in lungs (Fig. 5D) or isolated tracheal epithelial cells after viral infection (Fig. 5E).

PGD2 levels and lipocalin PGD2 synthase expression are increased in neonatal lungs during infection. Adults (white bars) and neonates (black bars) were infected with SeV (500 pfu/g body weight).
In vitro studies demonstrated that TSLP can stimulate PGD2 production in mast cells (2); however, we found that TSLP protein levels were significantly lower in neonatal mice despite high PGD2 levels (Fig. 6A). In uninfected adult lungs, protein levels of TSLP averaged 24 pg/mL. The adult levels of TSLP decreased slightly after infection, but these differences were not significant. In contrast, there was a significantly lower level of TSLP protein expression in neonatal lungs compared with adults at all time points tested. For the most part, TSLP levels were below the level of detection with immunostaining. Figure 6C shows the presence of TSLP in uninfected adult lungs where the expression was primarily localized to the airway epithelial cells, although some staining was also visible in a few cells in the lung parenchyma. TSLP protein was not detectable in adult lungs after SeV infection (data not given). There was no TSLP protein detected in uninfected (Fig. 6D) or infected neonatal lungs (data not given).
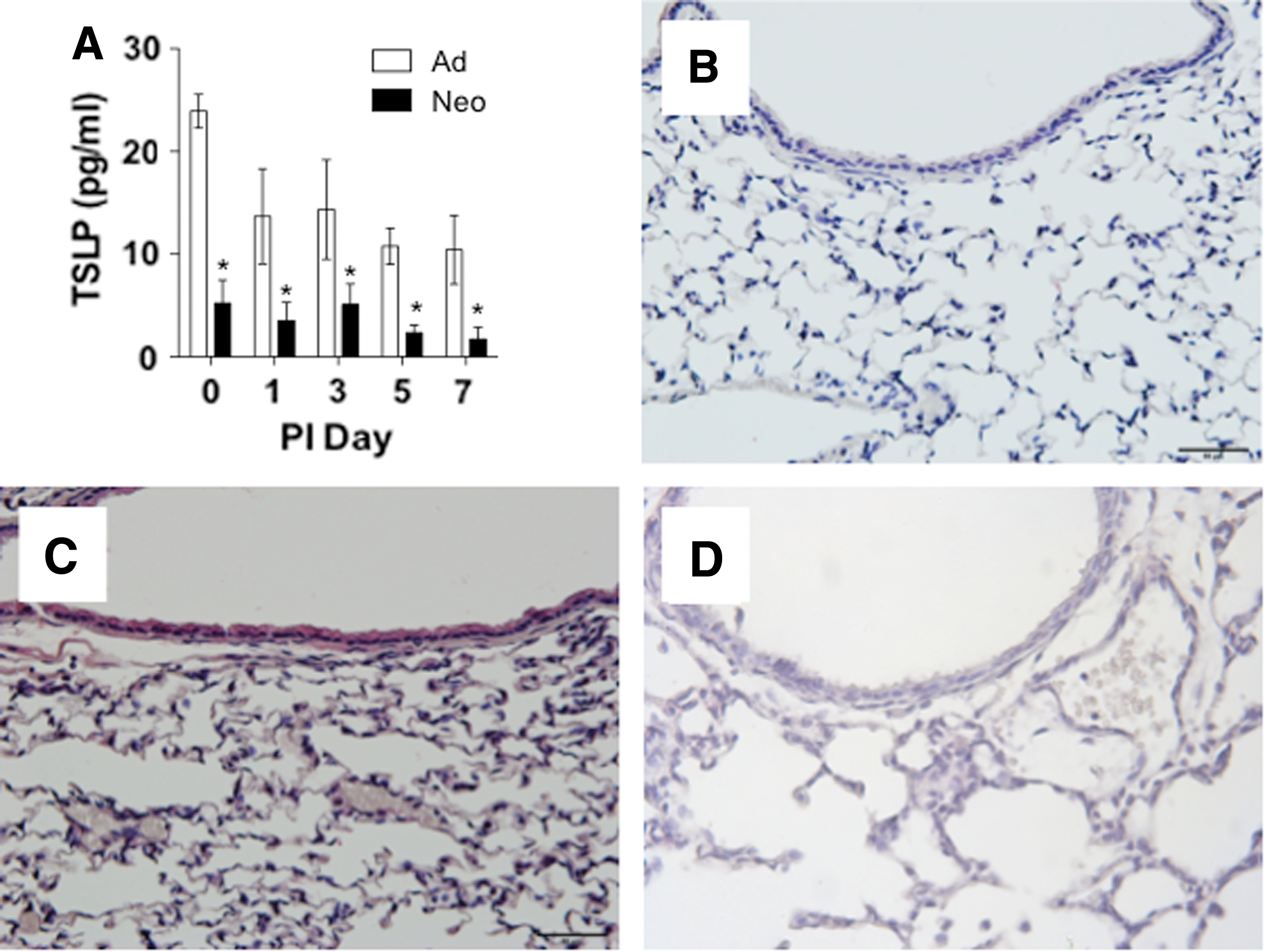
TSLP protein expression was lower in neonatal lungs compared with adults. Adults (white bars) and neonates (black bars) were infected as described.
Finally, to determine the mechanism of delayed cDC activation in neonates, PGD2 activity was blocked by treatment with the PGD2 receptor (DP1) antagonist (BWA868C). Adult and neonatal mice were infected intranasally with SeV with or without BWA868C, and lymph nodes were harvested on day 3 postinfection. Treatment with the DP1 antagonist increased the frequency of MHC Class II+/CD103+ cells in the draining lymph nodes on day 3, but this change was not significant (Fig. 7A). However, blockade of PGD2 DP1 signaling significantly increased MHC Class II expression on neonatal cDCs in the draining lymph nodes to levels similar to adults (Fig. 7B). Finally, neonatal MHC Class II+/CD103+ cells had significantly increased CCR7 expression after antagonist treatment (Fig. 7C). It is important to note that we also observed a significant population of eFluor+ cells (43.7%) that were MHC Class II+/CD103− (Fig. 1). Analysis of this population showed that there was no difference in frequency of MHC Class II+ cells in either the lung or lymph nodes between adults and neonates. Treatment with the DP1 antagonist also did not have an effect on the frequency of the MHC Class II+/CD103+ cells (Supplementary Fig. S1; Supplementary Data are available online at

Treatment with PGD2 DP1 receptor antagonist (BWA868C) restored lymph node cDC activation in neonates. Adults and neonates were infected with SeV (500 pfu/g body weight). One group of neonates also received the DP1 antagonist BWA868C. Flow cytometry of draining lymph nodes was performed on day 3 postinfection.
Discussion
The type and magnitude of an immune response to pathogens varies with the age of the host (10,16). We demonstrated here that activation of CD103+ cDCs that have migrated from the lung to the draining lymph nodes was delayed in neonates on days 3 and 5 after infection with SeV; this delay was associated with reduced expression of the activation markers MHC Class II and CCR7. Despite this delay in cDC activation, the numbers of SeV-tetramer+ CD8+ T cells were the same in SeV-infected adult and neonatal mice. This correlates with the observation that neonates were able to clear SeV with same time course and efficiency as adult mice (1). These results suggest that the delayed cDC activation does not impair viral clearance in neonates. Similarly, studies have shown that DCs from neonatal and adult mice are comparable in their ability to activate CD4+ T lymphocytes (3).
In contrast to the results with SeV infection, neonatal mice infected with the human paramyxovirus respiratory syncytial virus (RSV) showed reduced antigen processing and expression of costimulatory molecules on CD103+ cDCs; in this model, the reduced activation of the CD103+ cDCs resulted in attenuated and altered CD8+ T cell responses, which was not observed in the SeV model (21). These contrasting observations may be because of differences in the tissue tropism of the two paramyxoviruses. SeV is a natural mouse pathogen that infects and replicates in the airway epithelial cells; in contrast, human RSV infects the alveolar type II pneumocyte and does not replicate efficiently in the mouse (8). Consequently, a difference in innate sensor cells (pneumocyte vs. airway epithelial cell) may influence the activation of effector CD8+ T lymphocytes that are necessary for viral clearance.
The observed delayed migration of CD103+ cDCs in neonates during SeV infection was similar to the delayed migration of lung resident DCs in geriatric mice after infection with influenza A virus (IAV), severe acute respiratory syndrome coronavirus (SARS-CoV), or RSV (29). In contrast to neonates, delayed cDC migration in geriatric mice was associated with decreased virus-specific T cell responses and increased mortality. In order for CD103+ cDC migration to occur, the cells must upregulate CCR7 (24). Both CCL19 and CCL21 are CCR7 ligands and are important for dendritic cell survival, migration, and the expression of MHC Class II and costimulatory molecules (12). We found that CCR7 surface expression and CCL19 mRNA levels were significantly reduced in neonates compared with adults; however, CCL21 expression was significantly increased in neonates during infection. This suggested that the reduction of CCR7 and its cognate ligand CCL19 may contribute to the delayed migration of cDCs. To fully understand cDC migration in neonates, it will be important to analyze CCL21 expression in isolated lung endothelial cells and CCL19 levels in the lymph nodes. Delayed DC migration in geriatric mice during respiratory viral infection was also associated with reduced CCR7 expression (29). Together, these results suggest that, although there are some similarities, there are other important differences in the neonatal and geriatric immune responses to viral infection yet to be elucidated.
Prostaglandins are lipid mediators of immune response, and we observed that increased PGD2 levels and lipocalin PGD2 synthase levels in neonatal mice were associated with delayed cDC activation. This was consistent with the observation that instillation of PGD2 into adult mouse lungs inhibited the migration of lung DC to the draining lymph nodes in response to ovalbumin. In addition, increased levels of PGD2 were associated with delayed DC migration in geriatric mice during respiratory viral infections (11,29). Also, PGD2 receptor signaling protected against inflammation in a mouse model of LPS-induced acute lung injury. Mice deficient in hpgds had more severe endotoxin-induced lung injury, and treatment of these mice with a DP1 receptor agonist or PGJ2 (a degraded product of PGD2) both improved survival (20). The increased levels of lipocalin PGD2 synthase and PGD2 in neonates were associated with reduced inflammation (typified by reduced TNF-α and IFN-γ levels) (1). Interestingly, transgenic mice that overexpressed lipocalin PGD2 synthase through a constitutive β-actin promoter also demonstrated decreased IFN-γ (6). Together, these studies suggest that PGD2 has an anti-inflammatory effect in the lung and may be responsible for the reduced inflammation and delayed cDC activation in neonatal lungs during respiratory viral infection. Additional experiments will be needed to determine how PGD2 levels change with age in both infected and uninfected mice.
Our previous studies using bone marrow chimeras demonstrated that airway epithelial cells were critical in the innate response to SeV infection (23). Comparison of whole lung homogenates with freshly isolated tracheal epithelial cells suggested that lipocalin PGD2 synthase (ptdgs) expression was predominantly produced in the airway epithelium of neonatal mice. Although this cell preparation was highly enriched for airway epithelial cells, it did contain a mixture of cells types; future experiments will be needed to determine the precise cellular source of lipocalin PGD2 synthase.
Although the mechanism for increased PGD2 in neonatal and geriatric lungs is not known, it has been proposed that TSLP may contribute to increased levels of PGD2 (2). TSLP is important for DC recruitment to the epithelial cell surface after pathogen encounter; it induces DC maturation and expression of MHC Class II and costimulatory molecules, CD86 and CD40 (30). Dendritic cells activated by TSLP migrate to the draining lymph nodes and present antigen to naive CD4+ T cells. It has been suggested that viral infection may influence TSLP expression in vitro. RSV infection of primary airway epithelial cells and epithelial cell lines resulted in increased TSLP mRNA expression. We demonstrated, for the first time, differences in TSLP expression between adult and neonatal mice during respiratory viral infection. Results from ELISA and immunostaining suggested that there was basal expression of TSLP protein in adult lungs before infection and that levels decreased after SeV infection. In contrast, neonates showed significantly lower levels of TSLP protein at all time points. Because the levels of TSLP in neonatal lungs are very low, it is highly unlikely that it contributes to the observed increased PGD2 production.
To assess the role of increased PGD2 levels, neonatal mice were treated with the DP1 receptor antagonist BWA868C. This restored MHC Class II expression on lymph node cDCs to adult levels. In contrast, treatment with BWA868C reduced MHC Class II and CCR7 expression on the CD103− population in the lungs. These cells may be CD11b+CD11c+ and their role in the neonatal response to virus will require further investigation. Because the neonatal mice are still able to eliminate virus with the same time course and efficiency as adult mice, it is not clear what role PGD2 plays in the neonatal innate immune response. Additional studies will be needed to determine the effect of PGD2 blockade on inflammation and viral clearance in neonatal mice.
In summary, we demonstrated that neonatal mice expressed high levels of lipocalin PGD2 synthase and PGD2 in the lungs during respiratory viral infection. This was associated with reduced expression of the maturation markers MHC Class II and CCR7 on cDCs in the lymph nodes, and reduced expression of the chemokine CCL19 (Fig. 8). These differences did not affect the numbers of SeV-specific CD8+ T lymphocytes or the ability of the neonatal mice to clear virus from the lungs. Finally, treatment with the DP1 receptor antagonist BWA868C restored cDC activation in neonatal mice on day 3 postinfection, suggesting a potential role for PGD2 in the regulation of neonatal cDCs.
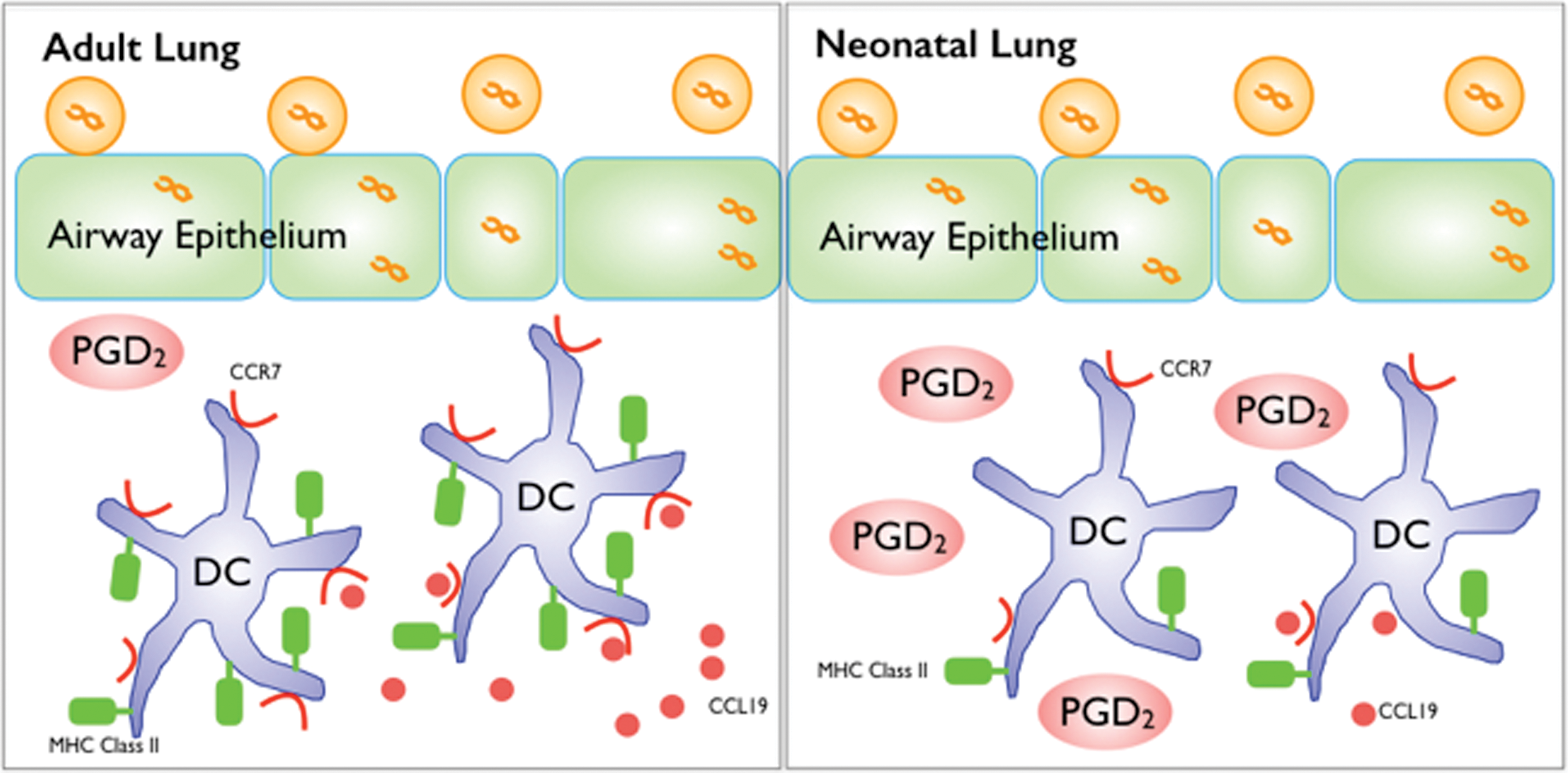
PGD2 delays cDC activation in neonatal lungs during respiratory viral infection. At early times postinfection adult lung cDCs have higher levels of MHC Class II and CCR7 expression. This is associated with increased levels of CCL19. In contrast, neonatal cDCs have reduced expression of MHC Class II and CCR7 and there is reduced CCL19 expression in the lung. This is associated with significantly increased levels of PGD2 in the neonatal lung. Color images available online at
Footnotes
Acknowledgments
The authors thank Shamaita Majumdar and Amber Wollam for technical assistance.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.