
Abstract
Porcine epidemic diarrhea caused by porcine epidemic diarrhea virus (PEDV) is a highly contagious disease in newborn piglets. The spike (S) protein is the surface glycoprotein of PEDV, which can induce specific neutralization antibodies and is a candidate antigen for vaccination attempts. In our study, the S1 region of PEDV strain CH/JLDH/2016 spike gene was inserted into the Lactococcus lactis expression vector, pNZ8149, resulting in recombinant plasmid pNZ8149-S1, and the immunogenicity of recombinant L. lactis pNZ8149-S1/NZ3900 was evaluated in mice. After immunization, significantly higher levels of anti-PEDV serum IgG antibodies and mucosal sIgA antibodies were detected in mice orally administered with pNZ8149-S1/NZ3900, compared with control groups pNZ8149/NZ3900, NZ3900, and phosphate buffered saline (p < 0.01). Lymphocyte proliferation assay results showed that the recombinant L. lactis pNZ8149-S1/NZ3900 significantly stimulated the proliferation of splenic lymphocytes (p < 0.01). In addition, the recombinant L. lactis vaccine could induce high levels of IL-4 and IFN-γ in immunized mice (p < 0.01). The results of our study suggest that the recombinant L. lactis pNZ8149-S1/NZ3900 can provide a promising vaccine strategy against PEDV infection.
Introduction
Porcine epidemic diarrhea (PED), characterized by vomiting, diarrhea, and dehydration, is a highly contagious intestinal disease caused by porcine epidemic diarrhea virus (PEDV). The infection rate of piglets is up to 100%, and the mortality rate of piglets is between 80% and 100% within 1 week (16). Many highly pathogenic PEDV variants have been isolated in swine from the farms in southern China since 2010. These viruses have rapidly spread to other countries in Asia, North America, South America, and Europe, resulting in huge economic losses (6,34). For example, there were about seven million piglet deaths due to the PEDV infection in United States in 2014 and the economic loss was $900 million to $1.8 billion (6). The epidemic of PEDV lasted till 2014. Moreover, PEDV is still the main cause of economic loss for swine industry in some Asian countries. Taking Japan as example, more than 490,000 piglet deaths were caused by PEDV infection between October 2013 and August 2015 (7). PED can result in high morbidity and mortality in newborn piglets (14). The morbidity and mortality of adult pigs infected with PEDV are relatively low (27). The mutation of S gene in prevalent strain was demonstrated to be a key factor avoiding the protection of vaccine among pigs (33).
PEDV, a member of the coronaviridae family, is an enveloped, single-stranded RNA virus whose genome length is 28 kb, comprising five open reading frames encoding an accessory protein and four structural proteins (2). The four structural proteins contain the 150–220 kDa glycosylated spike (S) protein, the 20–30 kDa membrane (M) protein, the 7 kDa envelope (E) protein, and 58 kDa nucleocapsid (N) protein (10). The spike (S) protein of PEDV is a type 1 membrane glycoprotein composed of 1,383–1,386 amino acids (aa) depending on the strain. The S protein can be divided into S1 (aa 1–735) and S2 (736–last aa) domains based on homology to the S proteins of other coronaviruses (9,12,17,29). As other coronavirus S proteins, the S protein of PEDV can interact with the cellular receptor when mediating viral entry and inducing neutralizing antibodies in the natural host (1,3). More specifically, previous studies have shown that the S1 domain includes the receptor-binding region and the main neutralizing epitopes (32). Furthermore, the S1 domain of S protein is known as a specific protein to determine the type of PEDV strains and considered a candidate protein for developing the diagnostic reagents for the detection of PEDV infection (4,17). Therefore, the S1 domain would likely be a suitable target for developing effective vaccines against PEDV.
Various methods have been used to control the outbreaks of PED. Many classic live vaccines, including CV777, Japan 83-5, and Korean DR13 strain without pathogenicity, eliciting a strong immune response, have been applied in sows and the classic PEDV infection was successfully controlled for 30 years (24,25,28). However, these traditional attenuated vaccines of PEDV usually showed low serum antibody titers against newly emerging PEDV strains since 2011, suggesting that the classic PEDV vaccine strain may only provide partial cross-protection against high-virulence strains (27). The low efficiency of immune protection is attributed to the significant antigenic variation of structural proteins and major antigenic sites between classical and emerging PEDV isolates (27,31). Inactivated and attenuated vaccines (intramuscular or subcutaneous injections) could induce maternal antibodies at low titers for a short time. Most of the maternal antibodies in the milk were digested by gastric acid and pepsin before entering the intestine (28). In addition, they could not induce enough virus-specific IgA antibodies to produce mucosal immunity (11).
IgG antibodies, also known as systemic circulating antibodies, generally exist in serum and play an anti-infective and toxin neutralizing role in the defense process. Different from IgG, sIgA can neither regulate immunity through Fc receptors of macrophages nor activate complements through classical pathways, so sIgA can't dissolve viruses and cells. sIgA antibody can directly bind to the adhesins of pathogenic microorganisms or cross-link with its polymers to prevent interaction between invasive pathogenic microorganisms and epithelial cells. It can isolate pathogenic microorganisms from binding to mucosal epithelial cells and achieve the purpose of isolating pathogenic microorganisms from invading organisms. In addition, sIgA antibodies can eliminate pathogenic microorganisms and remove them from the body. Therefore, choosing a food-grade expression vector to express heterologous antigens is an ideal direction for oral vaccine development.
Lactococcus lactis is a Gram-positive bacterium and does not produce spores. It has been identified as a safety-grade (generally recognized as safe [GRAS]) microorganism by the US Food and Drug Administration. Previous studies have shown that L. lactis has been widely used in genetic engineering, including as carrier presenting virus, bacterial antigens (13,18,37), recombinant L. lactis expressing specific allergens to regulate allergic reactions (15), and express cytokines to treat respiratory and digestive tract diseases (19,35,36). Therefore, L. lactis is an ideal antigen delivery vector to induce mucosal immune.
Developing a novel and efficient vaccine for oral administration based on L. lactis is a better formulation for immunoprophylaxis against PEDV infection. In our study, the S1 domain of PEDV strain CH/JLDH/2016 spike protein was used as the immunogen and L. lactis pNZ8149 was used as a delivery expression vector to construct genetically engineered recombinant plasmid pNZ8149-S1. Subsequently, the recombinant L. lactis pNZ8149-S1/NZ3900 was orally administered to mice. The results showed that L. lactis could induce mucosal, humoral, and cellular immunity of anti-PEDV in immunized mice, suggesting its application in oral vaccine development for PEDV.
Materials and Methods
Experimental animals
Eighty-six-week-old female BALB/c, pathogen-free mice were purchased from the Changchun Institute of Biological Products, China. All animal experimental procedures were carried out in accordance with the Regulations for Animal Experimentation of Jilin Agricultural University (JLAU08201409), and the animal facility was based on the National Institutes of Health Guide for the Care and Use of Laboratory Animals (NIH Publication No. 8023).
Virus, bacterial strains, plasmids
L. lactis NZ3900 was cultured in M17 broth medium at 30°C. The pNZ8149 vector and the recombinant pET-28a-S1 plasmid containing the S1 gene of a variant PEDV strain (PEDV CH/JLDH/2016; GenBank: MF346935.1) were stored or constructed at our laboratory.
Construction of pET-28a-S1 cloning plasmid
Total virus RNA was extracted from suspected positive pathogens with a commercial kit (Fastagen). First-strand complementary DNA (cDNA) was transcribed with Revert Aid M-MIV reverse transcriptase (Fermentas), and the cDNA template was used for polymerase chain reaction (PCR). Primers were designed on the S1 gene of PEDV CH/JLDH/2016 (forward primer: 5′-
Construction of recombinant L. lactis expression plasmid
The expression vector pNZ8149 was digested with SacI and NcoI restriction enzymes, purified using AxyPreP DNA gel purification kit (AxyPreP), and ligated with the S1 gene recovered from the plasmid pET-28a-S1. The ligated products were electroporated into NZ3900 cells using an electrotransformer (2,200 V, 200 Ω, 4.5 msec). The recombinant pNZ8149-S1 plasmids were identified by PCR (forward primer: 5′-TTCCACTTTGGTGCATGGTA-3′; reverse primer: 5′-TATGGCGAAGGTGGCTAAGT-3′) and sequencing. The positive recombinant L. lactis bacteria was named pNZ8149-S1/NZ3900 and the empty was named pNZ8149/NZ3900.
Western blot analysis
The recombinant L. lactis pNZ8149-S1/NZ3900 and pNZ8149/NZ3900 were cultured in M17 medium (containing 1% glucose) at 30°C for 24 h (OD600 ≈ 0.4) and induced by 1 ng/mL Nisin (Sigma) for 3–4 h. Cellular lysate samples were prepared; SDS-PAGE (sodium dodecyl sulfate polyacrylamide gel electrophoresis) and Western blot analyses were performed using Coomassie brilliant blue gel staining and anti-flag tag mouse monoclonal antibody or anti-PEDV hyperimmune pig serum as described previously (5).
Immunofluorescence assay
Both pNZ8149-S1/NZ3900 and pNZ8149/NZ3900 were induced by Nisin (Sigma). The bacterium solution (500 μL) was centrifuged at 12,000 rpm for 3 min at 4°C, and washed three times with 500 μL PBST. Then, the bacterial sediment was resuspended with 200 μL anti-PEDV hyperimmune pig serum (1:20 dilution) at 37°C for 2 h. Subsequently, cells were incubated with 200 μL fluorescein isothiocyanate (FITC)-conjugated rabbit anti-pig IgG (1:20 dilution) containing 1% Evans blue at 37°C for 2 h. The cells were visualized by fluorescence microscopy (Zeiss LSM710). pNZ8149/NZ3900 was used as negative control.
Vaccine preparation, oral immunization, and sample collection
Recombinant L. lactis was inoculated in M17 liquid medium (containing 1% glucose) and cultivated at 30°C for 24 h, and the process was repeated three times. The third generation of bacterium solution was cultured at 30°C until OD600 ≈ 1.0 and induced by Nisin, centrifuged at 12,000 rpm for 2 min at 4°C, and washed with phosphate buffered saline (PBS). Then, the bacterial sediment was resuspended in PBS with the titer of 1.3 × 1010 colony forming unit (CFU)/mL. The empty plasmid pNZ8149/NZ3900 and the host NZ3900 served as controls.
Eighty-six-week-old female BALB/c mice were randomly divided into four groups (n = 20) and orally immunized with pNZ8149-S1/NZ3900, pNZ8149/NZ3900, and NZ3900 with 1010 CFU per mouse or PBS. The entire immunization procedure was divided into three stages: first immunization (days 1–3), booster immunization (days 18–20), and last immunization (days 35–37). Five mice from each group were sacrificed by euthanasia at days 0, 21, 35, and 49 after first immunization. The sera, the contents of small intestine and cecum, and stools were collected. Meanwhile, the spleens were collected and washed with sterile PBS at day 35 after immunization. The contents of intestine and fecal samples were treated as described previously (20). Briefly, 0.1 g of fecal sample and the contents of intestine was suspended in 500 μL PBS containing 0.05 mol/L ethylenediaminetetraacetic acid and incubated at 4°C for 16 h. After centrifugation, the supernatants were stored at −80°C for evaluating humoral immune response.
Enzyme-linked immunosorbent assay analysis of antibody levels
The levels of IgG in the sera and sIgA in the contents of small intestine, cecum, and stools were measured using the mouse-derived PEDV-IgG enzyme-linked immunosorbent assay (ELISA) kit and mouse-derived PEDV-sIgA ELISA kit, according to the manufacturer's instructions, respectively (Shanghai Guyan Biotechnology Co., Ltd.). The standard curves were drawn according to the standard product in the kit to calculate the levels of IgG or sIgA antibodies in the samples.
Proliferation of lymphocytes from immunized mice
The recombinant strain pNZ8149-S1/NZ3900 induced by Nisin was prepared for SDS-PAGE electrophoresis. The gel was stained with 0.25 mol/L KCl for 10 min, and the target protein band was cut, ground, mixed with PBS, repeatedly frozen, and thawed at −80°C for 4 times, centrifuged at 12,000 rpm for 5 min at 4°C, and then the supernatant was collected to obtain purified S1 protein.
Splenocytes from immunized mice were prepared for lymphocyte proliferation assays as described previously (22). In brief, 100 μL cell suspension (2 × 106 cells/mL) were transferred to 96-well plates containing RPMI 1640 medium (added 10% fetal calf serum) in 5% CO2 at 37°C. The cells were stimulated with 5 μg/mL ConA as a positive control, 0.5 μg/mL or 5 μg/mL S1 protein were used to stimulate cells for 72 h, then added 10 μL MTT (5 mg/mL) for another 4 h incubation. The reaction was stopped by the addition of an equal volume of dimethyl sulfoxide for 10 min incubation at room temperature. Finally, absorbance at 570 nm was measured with Microplate reader (Thermo Scientific). Stimulation index was calculated with the equation: SI = OD570:sample/OD570:blank control.
Cytokine ELISA
ELISA assays were performed using IL-4, IFN-γ ELISA kits according to the manufacturer's instructions (Shanghai Lang Dun Biotechnology Co., Ltd.). IL-4 and IFN-γ concentrations of the sera were obtained from the standard curves from each ELISA plate.
Statistical analysis
All analyses were performed by two-tailed t-tests and one-way analysis of variance. GraphPad Prism 5.0 (GraphPad Software) was used to analyze the significance of differences between means. All data were obtained from at least three independent experiments, and the geometric means of titers were expressed as averages. All data were expressed as means ± standard error of the mean.
Results
Expression of PEDV S1 protein in L. lactis
The PEDV S1 gene's recombinant plasmid pNZ8149-S1 was first constructed (Fig. 1A). Then, both this plasmid and the empty vector (pNZ8149) were transformed into L. lactis NZ3900 by electroporation, resulting in the recombinant L. lactis pNZ8149-S1/NZ3900 and pNZ8149/NZ3900. The expression of S1 gene was detected in the cell pellets of pNZ8149-S1/NZ3900 and pNZ8149/NZ3900 by Western blotting using anti-flag tag mouse monoclonal antibody or anti-PEDV hyperimmune pig serum. The results showed that a specific immunoreactive band at 89 kDa was detected in the cell lysates of pNZ8149-S1/NZ3900 (Fig. 1B, lane 2, 4 or Fig. 1C lane 2), but not in pNZ8149/NZ3900 (Fig. 1B, lane 1, 3 or Fig. 1C, lane 1).
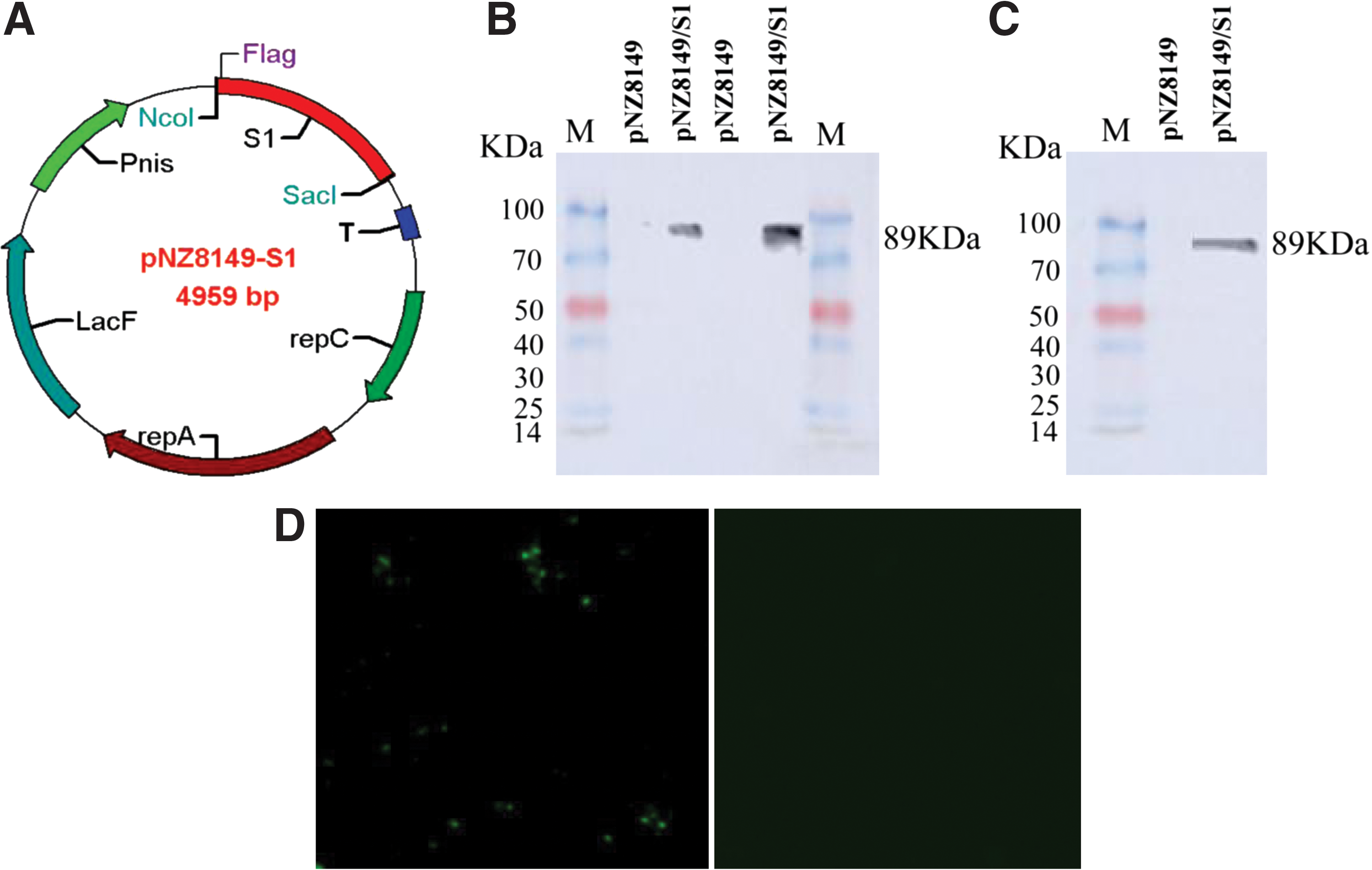
Construction and identification of PEDV S1 protein recombinant Lactococcus lactis expression plasmid.
Furthermore, the localization of the S1 antigen of pNZ8149-S1/NZ3900 was examined by immunofluorescence assay. The results showed that specific green fluorescence signals were observed on the surface of pNZ8149-S1/NZ3900 cells (Fig. 1D), confirming the presence of S1 on the surface of L. lactis. In contrast, no fluorescence signals were detected in pNZ8149/NZ3900.
Determination of anti-PEDV-specific IgG levels in sera of immunized mice
After oral immunization, the levels of anti-PEDV IgG in the sera of mice immunized with pNZ8149-S1/NZ3900, pNZ8149/NZ3900, NZ3900, and PBS were determined by ELISA. As shown in Figure 2, the mice immunized with pNZ8149-S1/NZ3900 produced a significant level of IgG antibody at 21 days post-immuned (dpi), which reached a peak at 35 dpi and decreased at 49 dpi, showing a significant difference compared with control groups (pNZ8149/NZ3900, NZ3900, and PBS; p < 0.001). The results showed that the recombinant strain pNZ8149-S1/NZ3900 elicited a significant level of humoral immune response in immunized mice.

Anti-PEDV-specific IgG antibody levels in the sera of mice, detected by ELISA. Mice (n = 5) were orally immunized with PNZ8149-S1/NZ3900, PNZ8149/NZ3900, NZ3900, and PBS, respectively. The anti-PEDV-specific IgG antibody level of recombinant L. lactis pNZ8149/NZ3900 immunized group was significantly higher than the control groups (**p < 0.01, ***p < 0.001). ELISA, enzyme-linked immunosorbent assay; PBS, phosphate buffered saline.
Determination of anti-PEDV-specific sIgA levels
Specific anti-PEDV sIgA antibody levels in the contents of small intestine, cecum, and stools of immunized mice were detected by ELISA. The sIgA antibody levels in stool samples are shown in Figure 3A, wherein a significant level of sIgA antibody was produced at 21 dpi in mice immunized with pNZ8149-S1/NZ3900 (p < 0.01), which continued to increase at 35 dpi, reaching the highest at 49 dpi, showing a significant difference compared with control groups (p < 0.001). Moreover, the sIgA antibody levels in the contents of small intestine (Fig. 3B) of the mice immunized with pNZ8149-S1/NZ3900 were significantly higher compared with the mice immunized with pNZ8149/NZ3900, NZ3900, or PBS (p < 0.01), having a tendency similar to that observed for stools (Fig. 3A). The sIgA antibody levels (Fig. 3C) in the contents of cecum of the mice immunized with pNZ8149-S1/NZ3900 were also significantly higher compared with control groups (p < 0.01). The results showed that the recombinant L. lactis pNZ8149-S1/NZ3900 could stimulate the mice to produce significant levels of mucosal immune response.
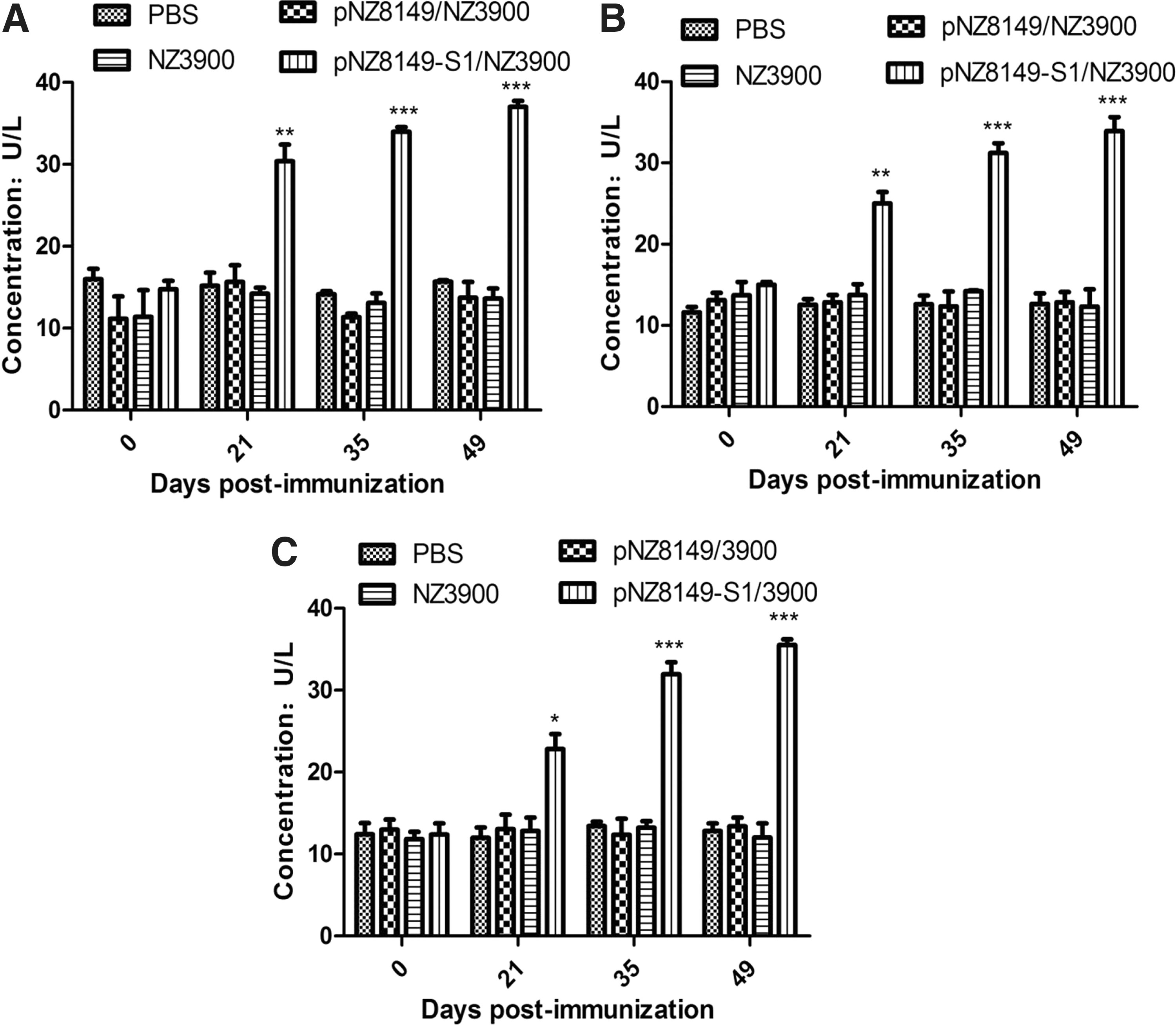
Anti-PEDV-specific sIgA levels in the stool
Lymphocyte proliferation
The mice were sacrificed and spleen cell suspensions were prepared for plating at 35 dpi. The proliferation of splenocytes upon stimulation with S1 protein was analyzed by MTT assay. As shown in Figure 4, in the pNZ8149-S1/NZ3900 immunized group, the stimulated index of mouse lymphocyte proliferation treated with 0.5 μg/mL PEDV S1 protein was significantly higher (p < 0.01) than the control groups (pNZ8149/NZ3900, NZ3900, or PBS) under the same conditions. When stimulated with 5 μg/mL PEDV S1 protein, the difference between pNZ8149-S1/NZ3900 and each of the control groups was extremely significant (p < 0.001).

Lymphocyte proliferation, determined by MTT assay, in response to recombinant S1 protein and ConA as stimulating agents in immunized mice. Different asterisks indicate significant differences (**p < 0.01 vs. control groups; ***p < 0.001 vs. control groups).
Determination of cytokines
The levels of cytokines IL-4 and IFN-γ in the sera of mice immunized with the recombinant L. lactis pNZ8149-S1/NZ3900 were detected by ELISA to assess cellular immune responses. As shown in Figure 5, the levels of IL-4 and IFN-γ in the pNZ8149-S1/NZ3900 group were significantly higher (p < 0.01) than the control groups (pNZ8149/NZ3900, NZ3900, or PBS), constantly increasing from 35 to 49 dpi, and the difference was extremely significant (p < 0.001). The results showed that the recombinant L. lactis pNZ8149-S1/NZ3900 could elicit Th1/Th2 mixed-type cellular immune response in immunized mice.
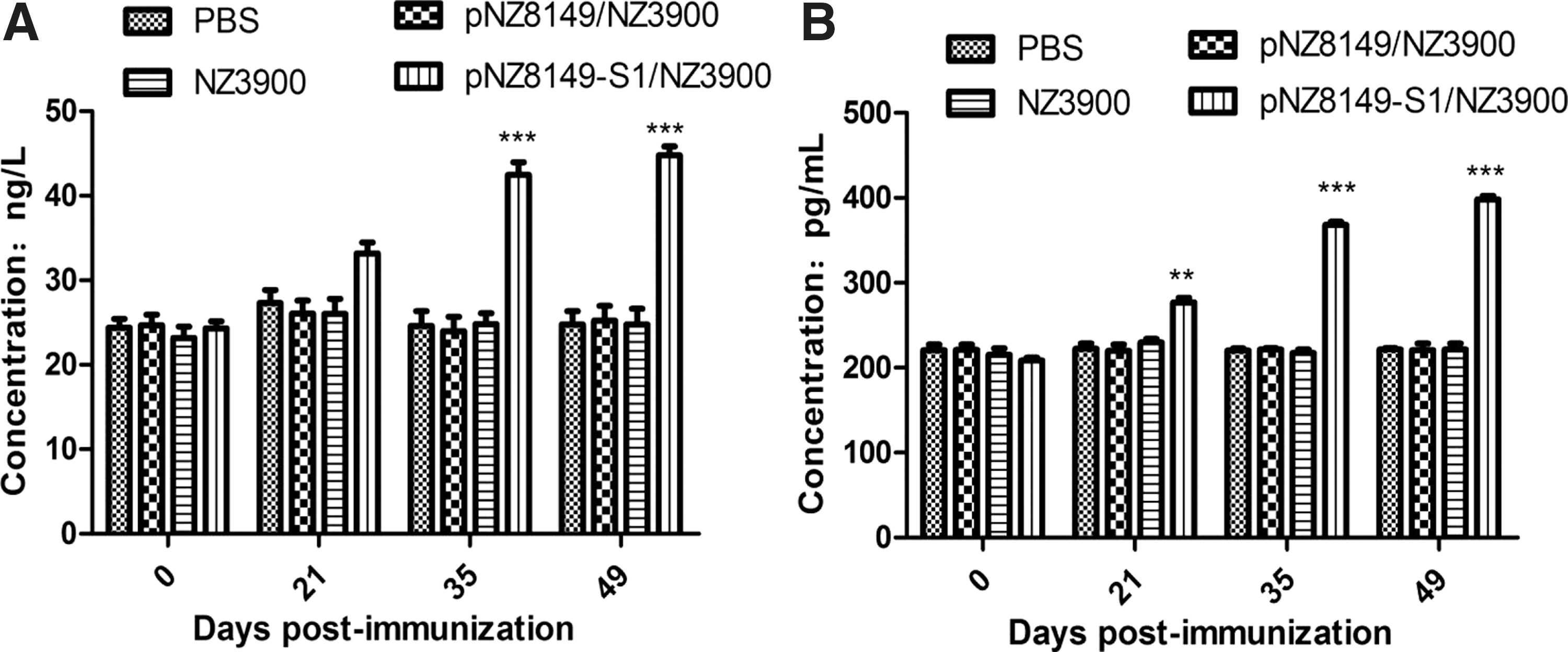
Cytokine response in sera of immunized mice. The levels of cytokines IL-4
Discussion
PEDV infects swine through the digestive tract and has intestinal tissue tropism. Intestinal villous epithelial cells of susceptible animals are target cells of the virus. When infected by PEDV, intestinal villi will atrophy and shed, leading to digestive disorders, acidosis, and dehydration. Anti-PEDV-specific IgG and sIgA antibodies could prevent the infection of PEDV. Therefore, vaccines must provide mucosal protection effectively in the intestinal tract. Mucosal immunity is more effective than systemic immunity in preventing virus entry into intestinal epithelial cells (18). Studies have shown that IgA's first line of defense in the intestine would be better than IgG in protecting piglets from PEDV infection (28). Therefore, developing an oral vaccine that can induce sIgA to anti-PEDV variant strain is necessary.
L. lactis, as an oral vaccine delivery vector, is safe, inexpensive, and stable, having the ability to stimulate the body to produce a comprehensive and systematic immune response to resist invasion by various pathogenic microorganisms (26). In this study, pNZ8149/NZ3900 expression system was used to express PEDV strain CH/JLDH/2016 S1 gene, and the recombinant L. lactis was orally administrated to mice. Results demonstrated that the recombinant L. lactis pNZ8149-S1/NZ3900 could elicit high levels of anti-PEDV-specific sIgA and IgG antibodies in the immunized mice. Rapid proliferation and differentiation of lymphocytes in a short time is an important process in the body to defend against all kinds of pathogens. When stimulated by antigen or ConA, lymphocytes could proliferate and differentiate—and in the process increasing the synthesis of lymphocyte nucleotides and proteins—into lymphoblasts. Therefore, we chose to detect the proliferation of spleen lymphocytes in vitro and to determine the immune protection of lymphocytes in making the body resistant to exogenous pathogens. In this study, our results showed that the recombinant bacteria could elicit cellular immune responses in immunized mice, consistent with the results of Meng et al. (23).
CD4+ Th cells could be divided into Th1 and Th2 subtypes, based on their biological functions (8). IFN-γ, secreted by Th1 cells, facilitates the synthesis of complement-associated IgG2a, and plays an important role in cellular immune response caused when pathogens invade the body. IL-4, secreted by Th2 cells, stimulates the proliferation and differentiation of B lymphocytes, playing an important role in the humoral immune response and promoting immune tolerance and mucosal immunity (30).
To explore the type of immune response induced by recombinant L. lactis pNZ8149-S1/NZ3900, the levels of IL-4 and IFN-γ were detected to evaluate the activity of T lymphocytes. Our data showed that the recombinant L. lactis pNZ8149-S1/NZ3900 stimulated the immunized mice to produce both Th1/Th2 cellular immune responses, and indirectly showed that pNZ8149-S1/NZ3900 has the ability to stimulate cellular immunity and humoral immunity in immunized mice. The changes in trends of IL-4 and IFN-γ levels in immunized mice were consistent with Liu et al. (21).
As far as we know, this is the first study to evaluate the immunogenicity of a recombinant L. lactis expressing S1 gene of PEDV mutant strain CH/JLDH/2016 against PEDV in mice.
Footnotes
Acknowledgment
The work was supported by the Key Scientific and Technological Project of Jilin Provincial Science & Technology Department (No. 20150204026NY).
Author Disclosure Statement
No competing financial interests exist.