
Abstract
Eosinophils have been mainly associated with parasitic infection and pathologies such as asthma. Some patients with asthma present a high number of eosinophils in their airways. Since respiratory viruses are associated with asthma exacerbations, several studies have evaluated the role of eosinophils against respiratory viruses. Eosinophils contain and produce molecules with antiviral activity, including RNases and reactive nitrogen species. They can also participate in adaptive immunity, serving as antigen-presenting cells. Eosinophil antiviral response has been demonstrated against some respiratory viruses in vitro and in vivo, including respiratory syncytial virus and influenza. Given the implication of respiratory viruses in asthma, the eosinophil antiviral role might be an important factor to consider in this pathology.
Introduction
Eosinophils, along with neutrophils and basophils, belong to a group of leucocytes known as granulocytes. They originate from bone marrow-derived CD34+ cells. Granulocyte-macrophage colony-stimulating factor (GM-CSF) (77), interleukin (IL)-3, and IL-5 (15,68) regulate eosinophil development, IL-5 being the most specific for the eosinophil lineage (111). Once mature, eosinophils migrate to peripheral blood with a half-life of ∼18 h (122).
Under homeostatic conditions, low numbers of eosinophils can be detected in peripheral blood (< 450–500 eosinophils/μL) (60). They rapidly leave the bloodstream and migrate into the adipose tissue, thymus, lungs, uterus, mammary glands, and mainly the gastrointestinal tract, where they are predominantly involved in the particular physiology of each tissue or organ (62,78,136). In contrast, during allergy, asthma, and parasitic infections, eosinopoiesis is increased, resulting in up to 5% of eosinophils in circulating blood (60).
The main feature of eosinophils is their high content in granular cationic proteins, which are responsible for their reactivity to the acidic dye eosin. In specific granules, also known as secondary granules, the major basic protein (MBP)-1 and MBP-2 are found within the granule core, whereas the eosinophil cationic protein (ECP), eosinophil peroxidase (EPO/EPX), and eosinophil-derived neurotoxin (EDN) are localized in the granule matrix (106). The granules of mature eosinophils also contain preformed cytokines, chemokines, and growth factors (136). Human eosinophils constitutively store interferon (IFN)-γ, IL-4, IL-6, IL-10, tumor necrosis factor (TNF)-α, IL-12 (p70), and IL-13, which can be rapidly secreted in response to specific stimuli without de novo transcription (121).
Eosinophils have been preserved during evolution, being present in vertebrates (62,72). For many years, the role of eosinophil in host defense focused on their response against helminth parasites. The dogma that these cells play crucial protective roles in helminth parasite infection began in the 1970s, and this notion was reinforced in the 1980 and 1990s, as demonstrated by the fact that either rodent and human eosinophils or their granular content was able to kill helminth larvae in in vitro assays (13,16,131).
With the development of eosinophil-deficient mice, new in vivo studies showed that this paradigm is mainly sustained to secondary infections (47,57). However, in the case of some helminths, such as Strongyloides stercoralis and Schistosoma mansoni, eosinophils are dispensable in host protection both in primary and secondary infections (85,126). In other cases, they may even play a beneficial role to the parasite, as the absence of eosinophils decreased Trichinella spiralis larvae growth and survival in skeletal muscle (34,36). In contrast, eosinophil presence or their products are associated with diseases that affect the skin, airways, and gastrointestinal tract (118). From an evolutionary point of view, a cell type would not persist if most of its functions were harmful to the host; so other unknown favorable ones may sustain its presence.
In recent years, several studies have demonstrated the functions of eosinophils that change their traditional description as a classic inflammatory end-stage cell involved in host protection against parasites. Among these were their involvement in homeostasis maintenance by secreting key cytokines, such as IL-4 that sustains alternatively activated macrophages in adipose tissue (141), and APRIL (a proliferation-inducing ligand) that generates and preserves immunoglobulin (Ig)A-expressing plasma cells in the gastrointestinal tract (17). In addition, the functions of eosinophils as effector cells have been reported in cancer response (19) and in bacterial, fungal, and viral infections (98), mainly against respiratory viruses (102).
Relationship Between Asthma, Eosinophils, and Respiratory Viral Infection
Asthma is a heterogeneous disease characterized by chronic inflammation of the airways. Symptoms include wheezing, chest tightness, shortness of breath, and coughing, which vary over time and intensity (39). According to the World Health Organization, there are ∼235 million people with this disease (5).
Patients with asthma can be classified based on the presented phenotype or endotype. While a phenotype describes “observable characteristics,” an endotype refers to “a subtype of a condition defined by a distinct pathophysiological mechanism,” and it is currently considered as a more accurate manner to classify asthma patients to choose their treatments (69). TH2-high endotype is the most prominent endotype and is mainly characterized by eosinophilic inflammation in the airways (125,137). Although it is the most studied endotype, its etiology is no clear (4).
Eosinophils release different molecules that participate in the pathogenesis of asthma. MBP has been found in sputum from patients with asthma and causes epithelial damage (37). It also induces the release of histamine from mast cells and basophils (86,94), which in turn induce bronchospasm and mucus secretion (138). Leukotrienes are potent bronchoconstrictors and cause the activation of eosinophils themselves, mast cells, basophils, monocytes, and T cells (80). Eosinophils also release cytokines that participate in asthma pathology. IL-13 induces airway hyperresponsiveness and mucus hypersecretion (40), while transforming growth factor (TGF)-β participates in tissue remodeling via fibroblast proliferation (28).
Asthma exacerbations can be triggered by environmental factors and, more frequently, by viral infections (127). Some of the viruses associated with wheezing illnesses are respiratory syncytial virus (RSV), human rhinovirus (HRV), human parainfluenza virus (PIV), influenza virus, metapneumovirus, coronavirus, bocavirus, and adenovirus (50). RSV is the main pathogen isolated from children younger than 3 years during winter months, and HRV is the most common virus during the rest of the year (46). Respiratory viruses are not only inducers of asthma exacerbations; they might also cause asthma development, at least in the case of RSV and HRV infections. Studies have shown that children with severe RSV bronchiolitis have a significantly higher risk of suffering from asthma at later ages (117). Meanwhile, others have found that wheezing illnesses caused by HRV predict asthma development in children (48,59,66).
Asthmatic individuals have a reduced innate immunity against respiratory viruses compared with non-asthmatics (30,49). The main difference between asthmatic patients and control subjects seems to be the impaired production of type I IFNs (IFN-α, IFN-β) (12,133) and type III IFNs (IFN-λ) (18) detected in asthmatics, which might be caused by eosinophil TGF-β production (71). However, more studies are needed because there are many reports that have failed to show differences in IFN production between these two groups; there is a general lack of data on asthma phenotypes analyzed in these studies; and most of them have focused on HRV (30).
Eosinophils are present in the airways in homeostatic conditions, so during viral infection with respiratory viruses, both elements may be in contact. In contrast, eosinophilic asthma is the predominant endotype of the disease (4); therefore, it is possible that eosinophils interact with respiratory viruses in this specific scenario. In fact, the involvement of respiratory viral infections in asthma exacerbations is widely accepted. In this review we summarize the eosinophil antiviral arsenal and analyze the in vitro and in vivo studies that support the eosinophil role against respiratory viruses, specifying when the antiviral eosinophil response is analyzed under an asthmatic condition.
Eosinophil Antiviral Arsenal
Eosinophils contain a group of receptors that allow the detection and response to viruses. They are also capable of developing different antiviral mechanisms and even participating in the initiation of adaptive T cell response (Fig. 1).
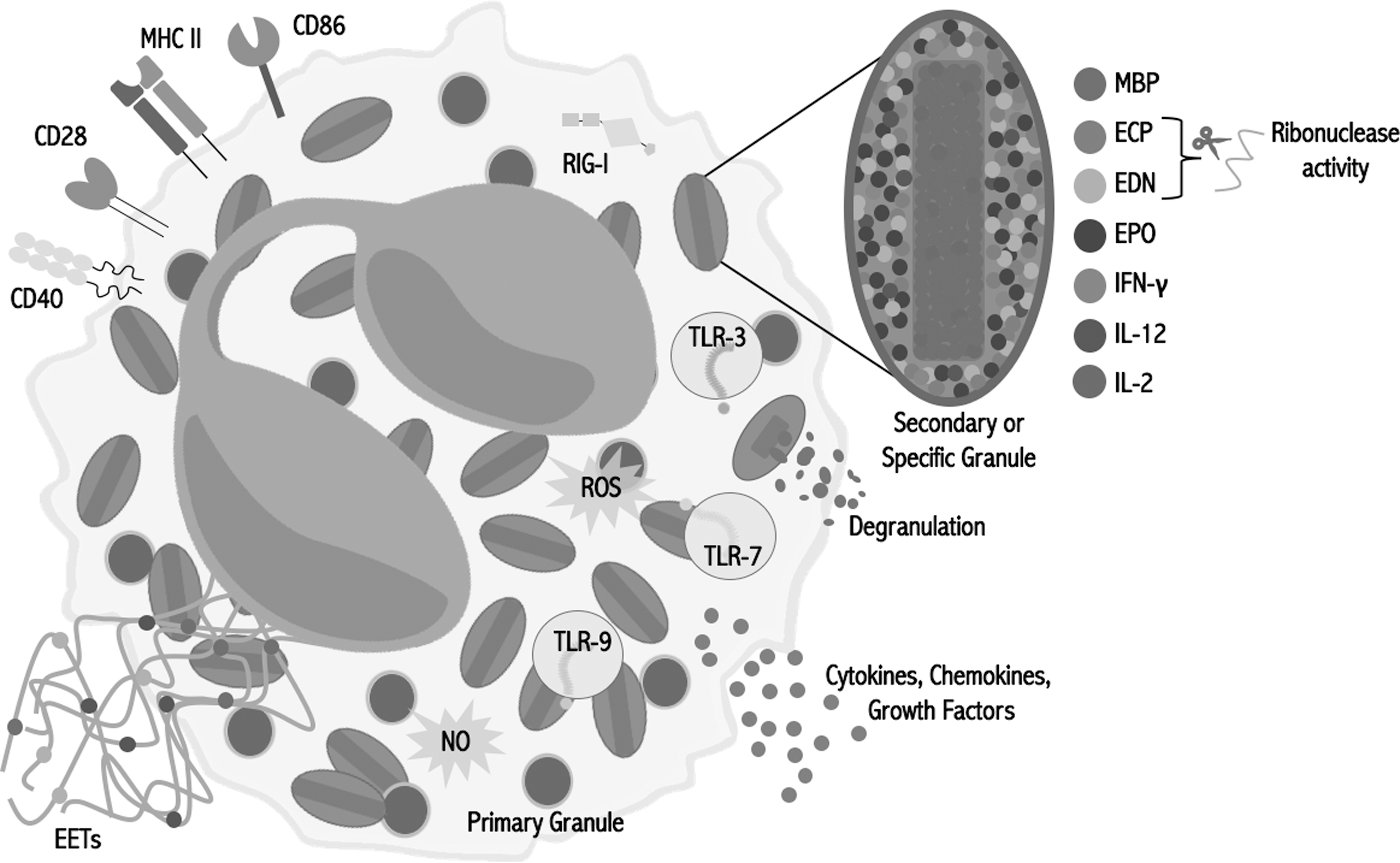
Eosinophil antiviral content. Eosinophils possess different molecules and mechanisms with potential antiviral activity, including TLRs, RLRs, preformed and inducible cytokines, antigen presentation molecules, EETs, and oxygen and nitrogen reactive species. ECP, eosinophil cationic protein; EDN, eosinophil-derived neurotoxin; EET, eosinophil extracellular trap; EPO, eosinophil peroxidase; MBP, major basic protein; NO, nitric oxide; RLR, RIG-I-like receptor; ROS, reactive oxygen species; TLR, toll-like receptor.
Viral recognition
Eosinophils express a large variety of toll-like receptors (TLRs) at the mRNA and protein levels, including those that participate in viral recognition such as TLR-3 (79,93), TLR-7, and TLR-9 (140). TLR-7, which recognizes single-stranded RNA (ssRNA) (23), stands out because its expression in eosinophils is higher compared with neutrophils (82,93). It has been shown previously that R-848 (Resiquimod), a TLR-7 synthetic ligand, prolonged survival and induced superoxide generation in eosinophils (82), suggesting the possible antiviral activity of this cell against ssRNA viruses.
Retinoic acid-inducible gene I (RIG-I) expression (mRNA and protein) has been found in eosinophils (61), which is a RIG-I-like receptor (RLR) that recognizes RNA sequences marked with 5′ triphosphorylated ends (67). However, RIG-I expression is low in eosinophils, especially when compared with neutrophils (61), and there are no studies that demonstrate its activity in eosinophils yet.
Antiviral immune response
Eosinophils possess and produce different molecules and develop several mechanisms with potential antiviral activity. ECP not only possesses antibacterial (14,65) and antiparasitic activities (14,41), it is also a member of the ribonuclease A superfamily along with EDN (38). It has been demonstrated that both proteins have RNase activity (101,119), so they could have activity against ssRNA viruses.
Eosinophils are also capable of producing oxidizing species by EPO, which uses hydrogen peroxide as a substrate to produce hypohalous acids (2,134). They also produce NO by inducible NO synthase (21), a molecule that inhibits viral replication by multiple mechanisms (1), and which is effective against several viruses, including influenza virus (99).
Even though eosinophils are frequently associated with TH2 responses, they are able to produce TH1, proinflammatory, and anti-inflammatory cytokines (115,121). Eosinophil granules contain preformed IL-2, IL-12, and IFN-γ (20,121), which are typical TH1 cytokines involved in antiviral responses (58,114).
Similar to neutrophils, eosinophils are able to produce extracellular traps (ETs) (142). Currently, there are no studies that demonstrate the antiviral activity of eosinophil ETs (EETs), but the antiviral effects of neutrophil ETs have been previously reported (112). Similar antiviral effect may be found in EETs.
Eosinophils as antigen presenting cells
Finally, although resting eosinophils do not express major histocompatibility complex (MHC) class II molecules, they could be induced in vitro by GM-CSF (70,95,132), IL-4, and IFN-γ (135). Moreover, eosinophils express co-stimulatory molecules such as CD80 (132), CD86 (132,139), CD28 (139), and CD40 (87,132). This is clinically relevant, since eosinophils recovered from bronchoalveolar lavage (BAL) of atopic patients after an allergen challenge (75,113) and from sputum of asthmatic patients (43) were found to express MHC II molecules.
Furthermore, it has been demonstrated that eosinophil acts as an antigen presenting cell (APC) in response to different antigens (95,132), including viral antigens (42,109), and is capable of inducing cytokine secretion by T cells (42,89,95,109,132). Additionally, eosinophils are able to migrate to lymph nodes, sites where they can present antigens to T cells (109,116,132).
In the next section we summarize and discuss the known antiviral activities of eosinophils against the main respiratory viruses in different models (Fig. 2).
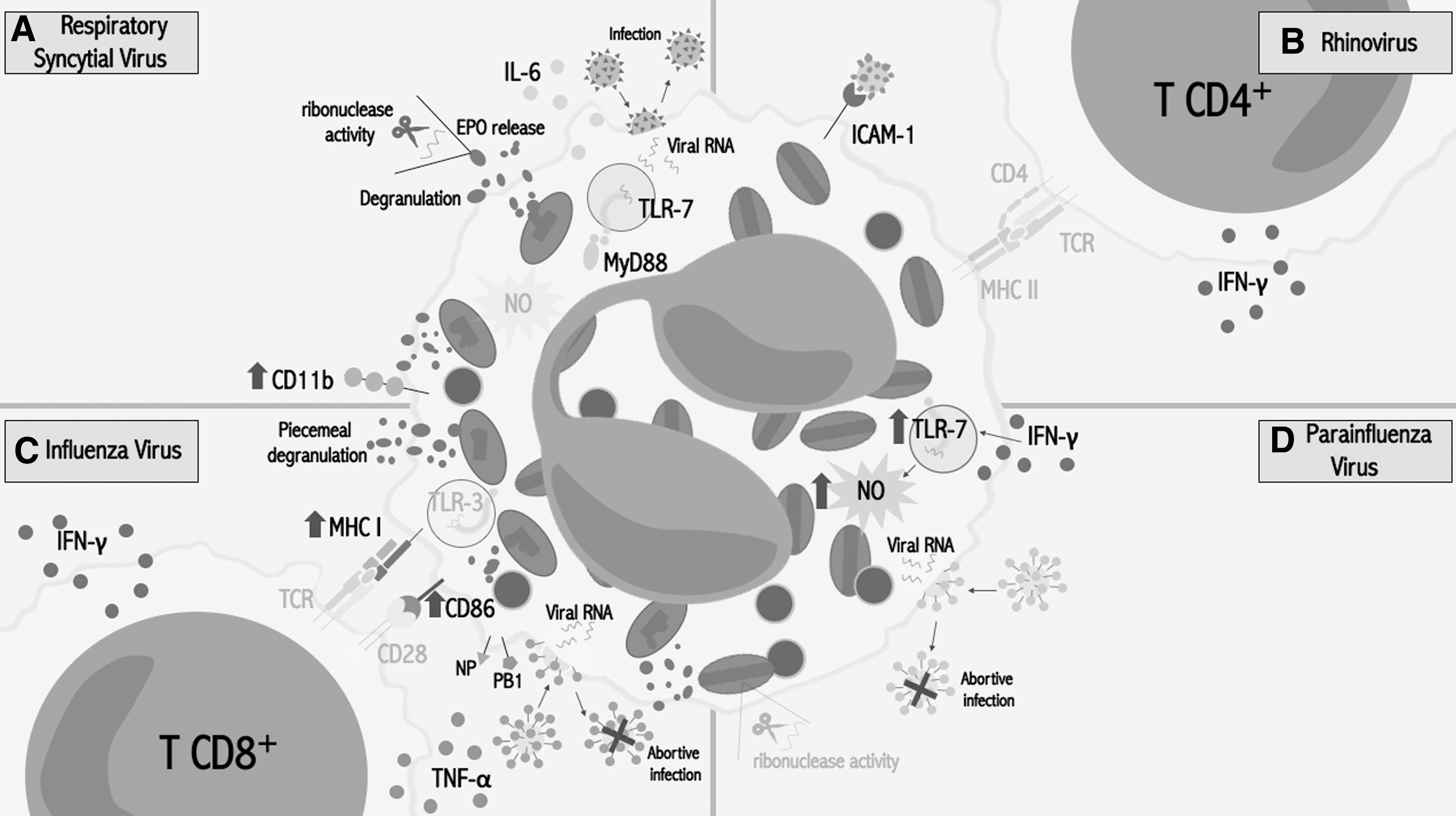
Eosinophil response against respiratory viruses.
Respiratory Syncytial Virus
RSV is an enveloped virus that contains a non-segmented, negative-sense ssRNA that belongs to the Paramyxoviridae family. RSV is one of the most important pathogens that causes airway infections during childhood (9). Approximately 50% of pneumonia cases and up to 90% of bronchiolitis cases during childhood are caused by this virus (26).
The first connection between RSV and eosinophils occurred in the late 1960s during trials of a formalin-inactivated RSV (FI-RSV) vaccine, which caused the death of two children after being exposed to wild-type (WT) RSV infection. Notably, this vaccine did not induce the production of protective antibodies (56). According to published histopathological findings, the fatal cases presented “peribronchiolar monocytic infiltration with some excess in eosinophils” (56,97). However, this histologic description was not accurate, as neutrophils and mononuclear cells were the most prominent infiltrating cells, as originally described in the autopsy and was also confirmed in another analysis of the original postmortem tissue (9,97). This inaccurate description caused researchers to conclude that eosinophils were responsible for the observed phenomenon (97).
Additionally, some reports have shown the presence of ECP and/or EDN in lung (44) and nasopharyngeal secretions (35) of children with severe RSV infection. These observations should be taken carefully, since none of these proteins are specific to eosinophils, as they can also be found in smaller amounts in other leukocytes such as neutrophils (124). Moreover, in BAL of RSV-infected individuals, neutrophils are the predominant cell type, and eosinophils constitute <1% of the cell population (33,74). A possible mechanism of eosinophil recruitment to the site of RSV infection is through Regulated on Activation, Normal T Cell Expressed and Secreted (RANTES) (54,105), which has been detected in the supernatant of human bronchial epithelial cells infected by RSV (7).
Controversies in these observations led some researchers to evaluate the eosinophil antiviral role against RSV. The fact that ssRNA is capable of inducing eosinophil degranulation and EPO release suggests its participation in RSV response (93). Some in vitro studies have demonstrated that lung epithelial cells infected by RSV trigger the release of ECP by eosinophils (88), which together with EDN have antiviral activity against RSV-B due to their RNase activity (24). Furthermore, when eosinophils are exposed to RSV-infected epithelial cells or to TLR-7 ligands, they overexpress CD11b (82,88,93), a surface molecule that has been reported as an activation marker in eosinophils (52,55), so they could be responding to viral infection.
The eosinophil antiviral activity against RSV has been also evaluated in in vivo models. Phipps et al. used IL-5 transgenic mice (IL-5Tg) (93), which are characterized by systemic eosinophilia (22), and WT mice. In these experiments an accelerated viral clearance was observed in mice with a higher number of eosinophils. In contrast, infected ΔdblGATA-1 mice (eosinophil-deficient mice (143)) had a lower viral clearance compared with WT controls (93). In the same study, the authors performed an eosinophil adoptive transfer from MyD88-deficient mice to WT mice. The mice that received MyD88-deficient eosinophils had a higher viral titer compared with those that received WT eosinophils. These results suggest that the observed antiviral effect depends on MyD88 signaling, probably initiated by TLR-7 (93).
Su et al. used eotaxin and IL-5 double-knockout (EotIL-5−/−) mice, which are characterized by undetectable eosinophils in blood and lungs (123). EotIL-5−/− mice that were immunized or not with FI-RSV presented a higher viral titer after RSV challenge compared with the controls. The adoptive transfer of eosinophils from IL-5Tg to FI-RSV-immunized and RSV-infected EotIL-5−/− mice led to a reduced viral titer and an increment in IFN-β production, compared with animals without eosinophil transfer (123). Notably, when a NO inhibitor was used, the effect of eosinophils against RSV was reversed (123), an event that has also been observed in IL-5Tg mice infected with RSV (93). These findings demonstrated the importance of NO in the observed antiviral effect, suggesting that it may be the main mechanism by which eosinophils reduce the viral titer (123).
Eotaxin-2/IL-5 double transgenic mice have been also used to study eosinophil antiviral activity against RSV (91). Besides systemic eosinophilia, these mice developed significant airway eosinophilia, and pulmonary pathologies representative of severe asthma, and presented eosinophils undergoing extensive degranulation (84). When eotaxin-2/IL-5 and WT mice were infected with pneumonia virus of mice (PVM—a severe RSV infection model in mice (100)), eotaxin-2/IL-5 mice presented a reduction in the virus recovered from lungs compared with the control strain, and they were also protected from a lethal inoculum of PVM (91).
In another assay, WT mice were sensitized and challenged with an extract of Aspergillus fumigatus and then infected with PVM. In the lungs of these mice, high levels of EPX and RNase activity, and degranulating eosinophils possibly by piecemeal degranulation, were found. These mice were also completely protected from a lethal inoculum of PVM, suggesting that, although activated eosinophils in the airways are considered detrimental to the host, they might promote the antiviral host defense (91).
Dyer et al. reported for the first time that mouse and human eosinophils can be infected by PVM and RSV, respectively. RSV infection induces the release of IL-6 by human eosinophils, while mouse bone marrow-derived eosinophils (bmdEos) infected with PVM release IL-6, IFN-γ-induced protein 10 (IP-10/CXCL10), monocyte chemoattractant protein 1 (MCP1/CCL2), and macrophage inflammatory protein 1-α (MIP-1α/CCL3). Interestingly, MyD88−/− bmEos presented an accelerated PVM replication and a decrease in IL-6 release, while the addition of exogenous IL-6 suppressed viral replication (29) (Fig. 2A).
Human Rhinovirus
HRVs are positive-sense ssRNA viruses, members of the Picornaviridae family. They are the most frequent cause of upper respiratory tract infection (51), being also the main viral pathogen isolated from patients with acute asthmatic exacerbations (53,120).
It has been reported that eosinophils bound to HRV 16 by intercellular adhesion molecule (ICAM)-1 in vitro (42), which was overexpressed in BAL eosinophils from allergic asthma patients (75). The incubation of peripheral blood eosinophils with HRV 16 demonstrated, for the first time, the capacity of these cells to act as an APC in response to viral antigens, inducing the proliferation of T CD4+ cells and the release of IFN-γ (42). In this context, IFN-γ might be increasing the expression of TLR-7 and TLR-8 on eosinophils (82), suggesting a cooperation between eosinophils and T cells (Fig. 2B).
In a recent publication, patients with mild asthma were treated with Mepolizumab (a humanized monoclonal antibody anti-IL-5) and challenged with HRV 16, which resulted in decreased but not abolished eosinophil count. This treatment did not prevent eosinophil activation upon HRV 16 challenge, and patients showed a higher viral load compared with those that received placebo (108), indicating a possible antiviral role of eosinophils.
Human Parainfluenza Virus
Human PIV is an enveloped, negative-sense, single-stranded, non-segmented RNA virus that belongs to the Paramyxoviridae family (11). They are major causes of lower respiratory infections in infants, immunocompromised patients, and elderly people (45), and have been detected in children with acute asthmatic exacerbations (53).
Adamko et al. used ovalbumin (OVA)-sensitized guinea pigs that were infected by PIV. Sensitized animals showed a ∼80% reduction in the viral content of lungs compared with non-sensitized guinea pigs. This effect was reverted by using anti-IL-5 antibodies, suggesting that the observed effect originated by the recruitment of eosinophils to the lungs (3).
Drake et al. used mouse models to evaluate the antiviral role of eosinophils against PIV. Similar to OVA-sensitized guinea pigs, C57BL/6 mice that were sensitized and challenged with OVA had a ∼90% reduction of viral RNA in the lungs 4 days post-infection (27). Because OVA sensitization and challenge also induces the recruitment of neutrophils and macrophages, authors infected NJ.1726 mice, which are characterized by a local accumulation of peribronchial eosinophils (64). The NJ.1726 mice also presented a viral RNA reduction compared with WT controls. Importantly, mice that constitutively expressed IL-5 but had eosinophil deficiency (NJ.1726-PHIL) behaved like the control mice after infection, suggesting that the antiviral effect was not due to IL-5. Furthermore, using EPX−/− mice, authors demonstrated that the antiviral effect was not caused by EPO (27).
The same group reported that human peripheral blood eosinophils isolated from healthy volunteers are susceptible to PIV infection. However, the viral progeny was not infectious, suggesting that the infection was abortive (27). Abortive infection has been also suggested in eosinophils infected with influenza virus (109), and it has been proposed as a passive mechanism by which eosinophils limit viral infection (27).
In the same study was also demonstrated that human eosinophils have antiviral activity to PIV in vitro, and this activity increased when eosinophils were pre-incubated with IFN-γ. This antiviral effect was mainly induced by NO production through TLR-7 (27). Although a NO-mediated antiviral effect of eosinophils against RSV was previously suggested by in vivo assays (93,123), this study demonstrated, for the first time, that NO production by eosinophils was indeed responsible for the decreased viral titer, at least against PIV. Besides, no RNase activity of eosinophil ECP and EDN was detected (27), which contrasts with a previous study reporting that RNase activity participated in the antiviral activity against parainfluenza (25). Contradictory results might be due to the different methodologies used (Fig. 2D).
Influenza Virus
Influenza viruses are enveloped, negative-sense, single-stranded, segmented RNA viruses that belong to the Orthomyxoviridae family (92,128). They are common causes of human respiratory infections, inducing high morbidity and mortality (128). Influenza virus induces acute asthma exacerbations in children and adults, especially during annual seasons (90). Interestingly, among patients hospitalized during the 2009 influenza pandemic, patients with asthma had less severe outcomes related to viral infection compared with non-asthmatics (10,31,73,81,83), although the asthma phenotype of these patients was not reported (130).
Samarasinghe et al. developed murine models of comorbidity by combining allergic asthma models (acute and chronic) with a model of influenza A virus (IAV) infection. In the acute asthma model, infected mice presented higher numbers of eosinophils in the airways and an accelerated virus clearance compared with infected mice with chronic asthma. Moreover, those mice with acute asthma presented higher numbers of CD8+ T cells and less epithelial damage compared with the chronic asthma model (110).
The same group evaluated the role of eosinophils against IAV by performing an adoptive transfer of eosinophils from A. fumigatus antigen-sensitized mice to the airways of IAV-infected mice. They observed a decrease in viral titer and an increase in virus-specific CD8+ T cells in recipient mice compared with non-transferred mice. Opposite results were obtained using infected ΔdblGATA-1 mice, supporting the role of eosinophils against IAV (109).
In the same study, through an in vitro model, it has been demonstrated that mouse eosinophils are susceptible to IAV infection, which triggers eosinophil piecemeal degranulation and an overexpression of MHC I and CD86 (109). Mouse eosinophils pulsed with viral peptides were able to induce IFN-γ and TNF-α release by CD8+ T cells obtained from IAV-infected mice, and to promote their proliferation (109). TLR-3, which was found to be elevated in the IAV model, might be recognizing the virus, although experiments have to be performed to demonstrate this. Authors suggest that a higher number of eosinophils in the airways of IAV-infected mice could induce better cellular immunity, considering that eosinophils have the capacity to prime CD8+ T cells, as well as to serve as APCs and also as a source of cytokines in the lung microenvironment (109) (Fig. 2C).
Perspectives
Recently a different population of eosinophils has been identified in the lungs of mice. This previously undescribed population, known as lung-resident eosinophils, differ from traditional eosinophils in their phenotype, function, and localization, being capable of inhibiting TH2 responses against inhaled allergens (76). It would be interesting to evaluate if the response against respiratory viruses from lung-resident eosinophils is similar to traditional eosinophils. In contrast, despite differences between eosinophils from healthy and asthmatic donors reported (6,52), the differences in antiviral responses from these cells have not been addressed.
The relationship between asthma and respiratory virus infection is not fully understood. Even though several studies have demonstrated the interaction between eosinophils and respiratory viruses using in vitro and in vivo models, knowledge about the exact mechanisms of eosinophil participation in the innate and adaptive immune response is lacking. For example, are eosinophils able to degranulate in response to PIV and RV as they do in response to influenza virus and RSV? Which granule proteins are released? Do they possess antiviral properties?
So far, the RNase activity of EDN and ECP has been observed only against RSV, failing to promote antiviral activity against PIV (27) or HRV (71). In contrast, EPO release has been observed when eosinophils are incubated with ssRNA, but its antiviral role has not been demonstrated against any virus yet. The clear lack of data is more evident in HRV infection, which is the main virus that induces exacerbations in asthmatics. Information is also needed to identify the relationship between specific allergens and the antiviral response, since it is known that in some models the antiviral response is allergen-dependent (91). Furthermore, most of the findings discussed above result from studies performed in mice. Although human and mouse eosinophils share many characteristics (63), they also possess differences (104), and the results should be carefully analyzed.
There are some reports that suggest the response and antiviral activity of eosinophils against viruses other than respiratory viruses, such as HIV (32,107), which requires further investigation. Likewise, the eosinophil antiviral activity against other respiratory viruses related to asthmatic exacerbations, including DNA viruses, has not been evaluated yet.
Although previously discussed studies clearly demonstrate the antiviral effects of eosinophils, whether eosinophils are necessary during viral infections in humans and their clinical significance remains unclear. Patients with asthma and chronic obstructive pulmonary disease (COPD) with high blood eosinophil counts have an increased risk of disease exacerbation (96,129). In fact, TH2-high asthma patients might have specifically an increased risk of virus-induced asthma exacerbations (8). Moreover, there is an increasing list of studies that demonstrate a reduced innate response against respiratory viruses in asthmatic patients (30).
The use of anti-TH2 approaches, for example, anti-IL-5, anti-IL-4, and anti-IgE biologics, has shown overall good results, reducing exacerbations in patients with eosinophilic asthma (30). It is plausible that their use could help to restore the defective innate immunity in these patients, but it needs to be demonstrated. Nevertheless, the antiviral role of human eosinophils should not be ruled out. Recent findings demonstrated that patients treated with anti-IL-5 presented a higher HRV load compared with the placebo group. These data suggest an important role of eosinophils in the control of viral infections (108).
As suggested by Rosenberg et al. (103), eosinophils might promote an antiviral response against respiratory viruses during specific scenarios, causing, however, a hyper-response attempting to eliminate the virus, leading to host damage (a double-edged sword). Further studies have to be performed to fully understand the relationship between eosinophils and respiratory viruses during certain conditions, such as asthma; this in turn will lead to a better understanding of eosinophil biology.
Footnotes
Acknowledgments
A.S.F.T. received a PhD scholarship from Consejo Nacional de Ciencia y Tecnología. We thank Sergio Lozano-Rodriguez, MD (“Dr. Jose Eleuterio Gonzalez,” Hospital Universitario, Monterrey, Mexico) for his review.
Author Disclosure Statement
No competing financial interests exist.