
Abstract
Despite the robust induction of humoral immune responses, a limitation of many adjuvants is their weak stimulation of cellular immunity. The development of synthetic gene-encoding adjuvants for simultaneous induction of both humoral and cell-mediated immune responses is under study. In this study, we examined the impact of toll/interleukin-1 receptor (TIR) domain of toll-like receptor 7 (TLR7) as molecular adjuvants on potency of inactivated infectious bursal disease (IBD) vaccines. A total of 60 specific pathogen-free week-old chicks were randomized grouped to receive either TIR-TLR7-adjuvanted IBD-inactivated vaccine or inactivated IBD antigen along with an unvaccinated control. Serum antibody titers were measured to estimate the humoral immunity, as well as lymphocyte proliferation activity for cellular immune responses. The protection was estimated after challenge with a very virulent IBD virus (IBDV) strain at 4 weeks postvaccination. The results indicated that one dose of IBD/TIR-TLR7 vaccine induced specific antibody responses, whereas a lower response after administration of inactivated IBD antigen was observed. The stimulation of splenocytes results indicated that the TIR-TLR7 adjuvanted IBD vaccine is capable of modulating cell-mediated immune response in treated chickens. A full protection against IBDV infection was achieved by injection of one dose IBD/TIR-TLR7 vaccine in the challenge trial. This study demonstrated that codelivery of TIR-TLR7 with inactivated IBD antigen resulted in simultaneous enhancing immune responses against IBD.
Introduction
Infectious bursal disease virus (IBDV) serotype I is an immunosuppressive virus that causes significant morbidity and mortality in young chickens. The most important feature of IBDV is evidenced by a severe prolonged immunosuppression of chickens infected at an early age (5,16,24). Whole inactivated vaccines are generally effective in preventing and controlling of IBDV infection (17). Consequently, the vaccines require strong adjuvant and boosting to ensure an appropriate immune response.
Besides improving antigen delivery to antigen-presenting cells (APCs), adjuvants induce the production of immunomodulatory cytokines. Through this modulation, T-helper types 1 and 2 immune responses were developed to vaccine antigens (2). Commercial vaccine adjuvants include alum, saponins, and oil emulsions for a wide range of viral diseases have been available for several decades. Most adjuvants can effectively stimulate T helper cells to promote the humoral immune response; however, this process is needed for the production of protective antibody levels in vaccination of chickens against an intermediate IBDV strain (21,26). Interest in the induction of cell-mediated and antibody responses, reduction of the amount of antigen, and longer duration of immunity has led to the development of new adjuvants such as immune-stimulating complexes, liposomes, nanoparticles, a wide variety of bacterial derivatives, and cytokines (13,19,23).
The use of cytokines as alternatives to existing adjuvants is attracting extensive attention. Interlukines provide signals to direct the immune response toward either an antibody-mediated or a cell-mediated response. Coadministration of plasmid-encoded chicken IL-18 and IL-2 with IBD vaccine effectively enhances the IBDV-specific antibody response (13,19). Providing high protection against IBDV challenges proposes the adjuvant effect of the immunoregulators. The immunoadjuvant activity of ChIL-1 has also been assessed using tetanus toxoid as an antigen. It significantly increased the antigen-specific response to injected antigen and promoted cell-mediated immunity (23).
Toll-like receptors (TLRs) and toll/interleukin-1 receptor (TIR) domain play crucial immune responses (12). The TIR domain of ca. 200 amino residues is composed of three conserved regions: box 1 (FDAFISY), box 2 (GYKLC-RD-PG), and box 3 (a conserved W surrounded by basic residues) specialized in interferon type I (IFN-I) production (18). This domain is also shared by the downstream cytosolic adapter protein MyD88, which formed a TIR–TIR interaction and couples the receptors to downstream signaling mechanisms. The TIR and non-TIR regions are important for inducing effective immune responses by targeting MyD88 to the correct cellular location (18). Signal transduction through recruitment of TIR domain ultimately leads to the activation of the nuclear factor-κB (NF-κB) transcription factor in B and T lymphocytes that regulates the expression of cytokines, lymphocyte activation, and immunoglobulin isotype switching (3,20).
We previously determined the potential role of TLR7-TIR domain in the induction of immune responses against IBDV infection in an in silico study and in the experimental trial using coadministration of the TIR domain sequence and the expressed IBDV VP2 in baculovirus (8,9). In this study we describe the immunization procedure of chickens using application of an inactivated IBDV antigen alone or in combination with TLR7-TIR through boost regimen to provide an outlook into introducing a new potential adjuvant.
Materials and Methods
Virus and antigen preparation
The IBD07IR (GenBank accession no. KT633995) intermediate strain of IBDV was injected onto the chorioallantoic membrane of 9–11-day-old specific pathogen-free (SPF) chicken embryos (Razi Institute, Karaj, Iran). The infective embryo and amnioallantoic fluid were harvested, pooled, and grinded. The antigen was clarified by centrifugation at 5,000 rpm for 30 min at 4°C. The viral fluids (EID50 = 108.17/mL) were inactivated with 3 mM of binary ethylene imine (Sigma-Aldrich). Inactivation procedure was confirmed by inoculating 0.1 mL of the inactivated virus into five 9–11-day-old SPF chicken embryos, harvesting the fluid, and injection into an additional five eggs for three times.
Adjuvant synthesis
Based on our previous studies (8,9), the cDNA encoding the conserved TIR domain of TLR7 (TIR-TLR7) contains IL-1 receptor with DIILIFLEKE amino acid sequence chosen as a biologic adjuvant.
Chicken immunization trial
A total of 10 μg of TIR-TLR7 were mixed with one dose of inactivated IBDV antigen and used for immunization of each bird through the intramuscular route. A total of 60 mixed-sex layer SPF week-old chicks were distributed randomly in three groups (20 birds in each group) as follows: unvaccinated control (given phosphate-buffered saline [PBS]), inactivated IBDV antigen, and IBDV/TIR-TLR7. Both treated groups received booster at 1-week postprime based on their prime injection. The animal experiments were carried out in accordance with the National Institutes of Health guide for the care and use of laboratory animals no. 8023, revised 1978.
Serum antibody responses in immunized chickens
Blood samples were collected from chickens before immunization and with 1-week interval up to 8 weeks postimmunization, and the IBDV-specific antibody responses were measured by enzyme linked immunosorbent assay (ELISA) (IDEXX). Least squares means (±standard deviation) were used for the effect of TIR-TLR7 on antibody titer against inactivated IBDV antigen in SPF chickens during the experimental period.
Cell-mediated immune response in immunized chickens
Cell-mediated immune response was measured using in vitro cell proliferation assay evaluated by MTT (3-[4,5-dimethylthiazol-2-yl]-2,5 diphenyl tetrazolium bromide). At 6 weeks of age, single cell suspension was generated from the harvested spleens from each group in PBS (pH 7.2). The splenocytes were pelleted for 10 min at 1,000 rpm and resuspended in RPMI-1640 medium (Sigma-Aldrich) supplemented with 10% chicken serum. The viability of cells was checked by trypan blue dye. The splenocytes were pooled separately and cultured with RPMI-1640 at a seed rate of 1 × 106 cells/well and incubated in triplicate with the respective IBDV vaccine antigens. Concanavalin A (Sigma-Aldrich) was used as the positive control to stimulate lymphocyte proliferation and cells were incubated in RPMI-1640 medium without antigen stimulation as the negative control followed by incubation at 37°C and 5% CO2. After 60 h, 20 μL of MTT (5 mg/mL) was added to all the wells and incubated for further 4 h. Then 100 mL dimethyl sulfoxide (DMSO) was added to the wells to dissolve the formazan crystals. The amount of MTT formazan produced during the incubation of splenocytes was measured by an ELISA reader at the wavelength of 570 nm. The results are expressed as a stimulation index (SI) calculated as mean optical density (OD) of stimulated splenocytes − mean OD of blank/mean OD of unstimulated splenocytes. The SI ≥2.5 was considered positive.
Determination of protection from challenge
Four weeks after second vaccination, five chickens from each group were challenged with 102 mean chicken infectious dose (CID50) per bird of a very virulent IBDV strain (GenBank accession no. EU224392) by the eye drop route. Protection against IBDV was assessed for 2 weeks after challenge. All birds were observed daily for 2 weeks for any clinical signs and mortality.
Statistical analysis
The comparison between the experimental groups versus the control was performed using one-way ANOVA and the Tukey test. p ≤ 0.01 was considered to indicate a statistically significant result.
Results
Humoral immune response in vaccinated chickens
Vaccinated and control chickens were evaluated for the presence of serum antibodies to IBDV antigens during the time course of the experiment. IBDV antibody levels, as monitored by ELISA, were detected in all chickens in the group vaccinated with IBDV/TIR-TLR7, whereas no specific antibody to IBDV could be detected in the control group that had been immunized only with PBS. According to the manufacturer's instruction, titers >396 are considered positive. The humoral antibody levels did show a significant difference (p < 0.01) between the IBDV/TIR-TLR7 group and the inactivated IBD antigen group (Table 1). This is supported by serological evidence that TIR-TLR7 may effectively enhance higher specific IBDV antibody titer in serum of immunized chickens. Different rates of decline between the two treatment groups were found, and by 7 weeks postvaccination, serum antibody levels in inactivated IBD antigen group were near detectable limits, whereas antibody titers remain stable in IBDV/TIR-TLR7 immunized group throughout the experiment period. The detected antibody levels indicated that the adjuvanted vaccine candidate could efficiently induce the humoral immune responses in SPF chickens.
ELISA Antibody Levels Against Infectious Bursal Disease Virus
Blood samples were taken from control and treated chickens immunized with inactivated IBDV antigen in the presence and absence of TIR-TLR7 at defined weeks postvaccination.
The prime and boost vaccinations were conducted at 1 and 2 weeks of age, respectively.
Different letters in each column indicate significant difference (p < 0.05). The overall p values for the effect of treatment, week, and the interaction of treatment by week was p < 0.0001.
IBDV, infectious bursal disease virus; SPF, specific pathogen-free; TIR, toll/interleukin-1 receptor; TLR, toll-like receptor.
Cell-mediated immune response in vaccinated chickens
Using splenocytes proliferation, the ability of IBDV/TIR-TLR7 to induce cell-mediated immune responses was measured. After IBDV recall stimulation, specific proliferation was observed in the IBDV/TIR-TLR7 by increasing the mean SI to 5.142 (Fig. 1). As shown in this figure, the lymphocyte response level of the IBDV/TIR-TLR7 group was significantly enhanced compared with the inactivated IBD antigen and control groups (p < 0.001). No significant differences were observed between the inactivated IBD antigen and control groups (p < 0.01). The magnitude of splenocytes proliferation indicated that TIR-TLR7 also plays an important role in the induction of cell-mediated immunity in vaccinated chickens.
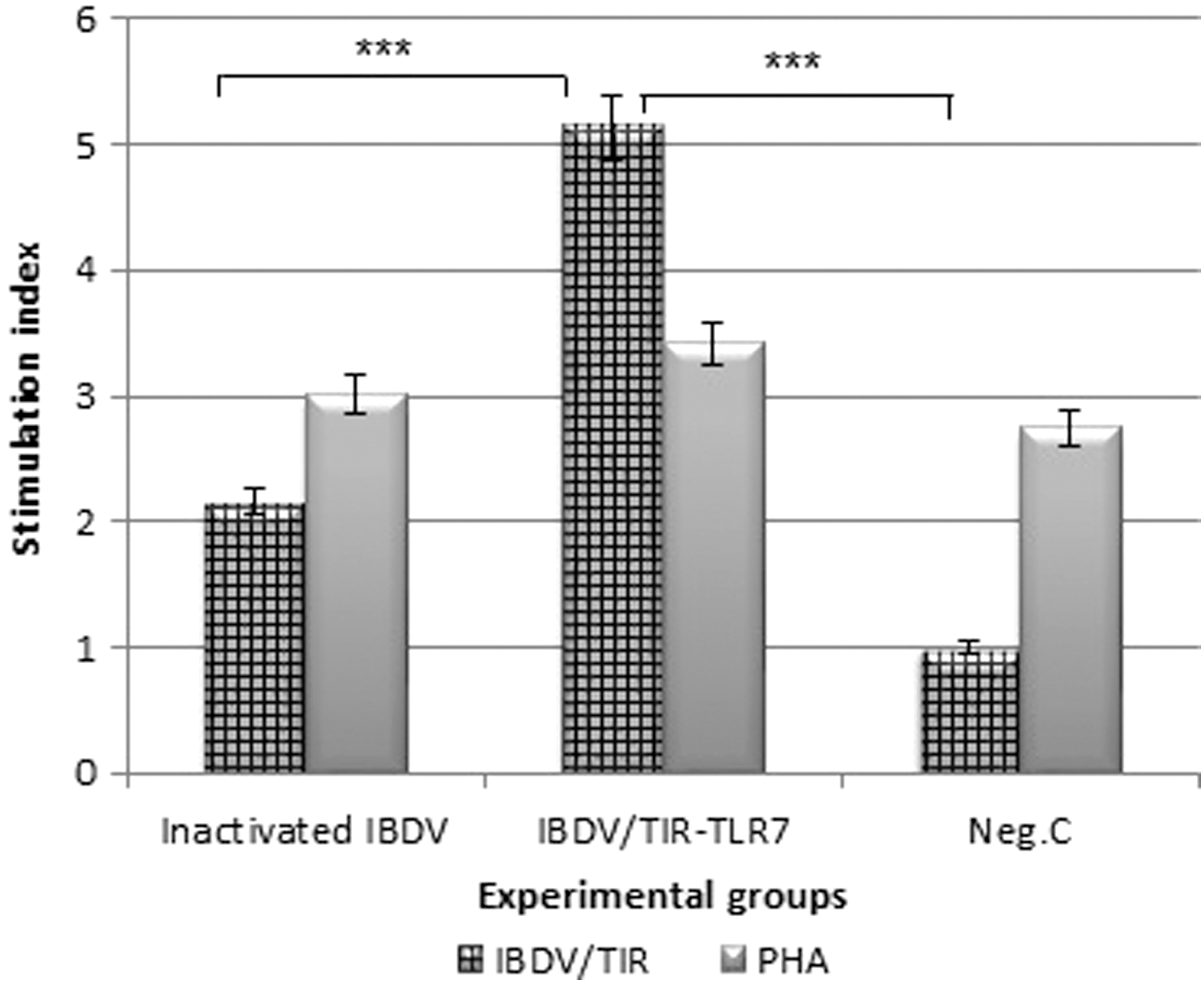
Cell-mediated immune responses to infectious bursal disease vaccination. The lymphoproliferative response shown as a SI of splenocytes from chickens immunized with the IBDV/TIR-TLR7 candidate vaccine and with inactivated IBD antigen compared with the control. The SI ≥2.5 is considered as positive response. Statistical differences were indicated by asterisk (***p < 0.001). IBDV, infectious bursal disease virus; SI, stimulation index; TIR, toll/interleukin-1 receptor; TLR, toll-like receptor.
Protection of vaccinated chickens
All of the chickens that were vaccinated with IBDV/TIR-TLR7 and then challenged with the vvIBDV were alive and did not exhibit any clinical symptoms of IBD (Fig. 2). In contrast, two chickens from the inactivated IBDV group showed the IBD clinical signs after the challenge test. All of the control group chickens showed the IBD clinical signs and two of them died at 4 dpi. To summarize, the administration of IBDV/TIR-TLR7 protected 100% of the SPF chickens against challenge with the vvIBDV.
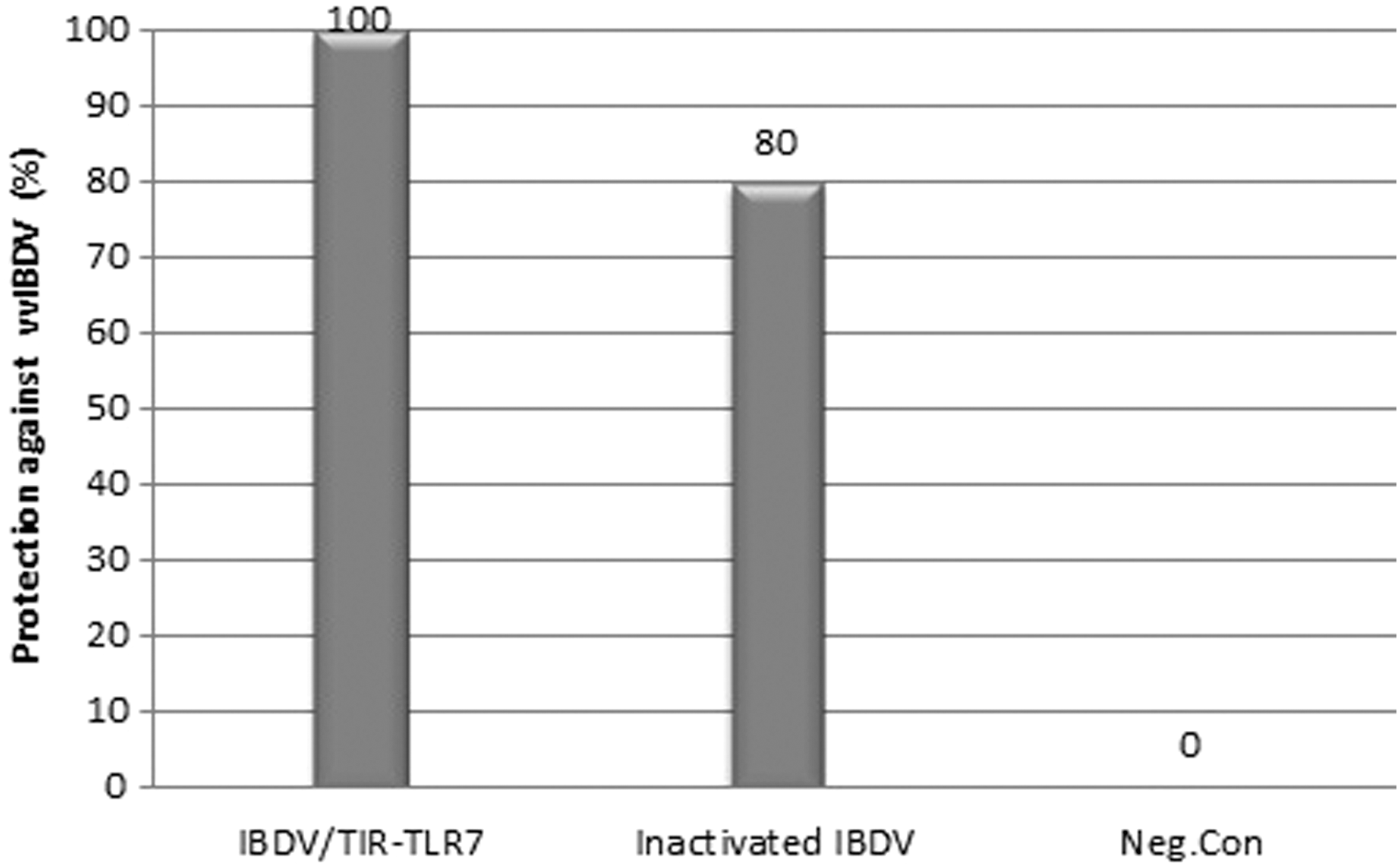
Protective efficacy of IBDV/TIR-TLR7 against vvIBDV challenge. All chickens in control group died after challenge with vvIBDV at 4 weeks of age, whereas 100% and 80% of chickens vaccinated with IBDV/TIR-TLR7 or inactivated IBDV antigen, respectively, were protected against IBDV challenge.
Discussion
In vaccine production industry, formulation with adjuvants is necessary for induction of an earlier onset of immunity, an overall stronger immune response, a longer duration of immunity to the vaccine, and interplay between innate and adaptive immunity. New adjuvants are being developed that are safe and highly effective for induction of both cell-mediated and antibody responses by targeting antigen to APCs and directing Th1 or Th2 immune responses. We coadministered an IFN-I inducer domain of TLR7 with inactivated IBD vaccine to enhance immune responses in SPF chickens. The results showed that TIR-TLR7 could significantly enhance the antibody titers in IBD vaccine immunized chicken as well as significantly promote the cell-mediated immunity as the comparison with inactivated IBD antigen alone group. The generation of a long-term protective immunity is an important measure of a vaccine's efficacy that is a desired outcome during vaccination. To assess such protective properties of the IBDV/TIR-TLR7, efficacy of the adjuvanted IBD vaccine was assayed by challenging chickens with a very virulent virus. An exciting finding is that chickens vaccinated with IBDV/TIR-TLR7 had complete survival versus inactivated antigen vaccinated and control groups. These results show that the formulations can further influence the protective efficacy against challenge with IBDV compared with the antigen alone. In vaccination against pathogens, the immune system relies on the ability of adjuvants to generate long-term memory and protection (1). Although the duration of humoral immunity is most important in immunization against IBDV, the development of efficacious vaccines seems to be essential to protect chickens by inducing T cell as well as B cell immunity without interfering with the maternal antibodies. Viral clearance is related to the immune response to IBDV, and CD8+ T cells are important contributors to virus clearance. The critical role of T cells in promotion of IBDV clearance indicates the importance of cell-mediated immunity in preventing IBD (22). Our important findings are the ability of TIR-TLR7 to mediate both humoral and cellular responses and also to provide a greater level of protection against IBDV infection.
TLRs are evolutionarily conserved innate receptors expressed in various immune and nonimmune cells, which also play an informative role in directing acquired immunity (7,12,20). TLR signaling is highly dependent on TIR–TIR interaction formation. Upon stimulation, the TIR domain of the cytoplasmic portion of the TLRs directly associates with the TIR domain in the C-terminal portion of MyD88. The interaction mediates in two different downstream pathways activation of AP-1 transcription factor through the mitogen-activated protein kinase cascade and activation of the IκB–kinase complex (11). Engagement of TLR signaling pathways triggers an adaptive immune response after immunization. The activated TLR upregulates expression of the costimulatory molecules CD80/CD86 and CD40 on the surface of dendritic cells (DCs), resulting in enhancement of APC activity (4,14). Besides their primary function in regulation of immune response, TLRs can also act as immunoadjuvant (10). A good vaccine adjuvant attracts innate cells such as activated macrophage and DCs. The ability of these cells to regulate immunity and to provide signals to naive T cells is dependent on their maturation (25). Mature and active DCs are inducers of regulatory CD4+T cells, which recognize MHC class II complexes presented by APCs and differentiate into different effector T cell subtypes or regulatory T cells. The antigen present on CD4+ and CD8+T cells in association with MHC classes I and II molecules is necessary for the development of both B cell and cytotoxic T lymphocyte (CTL) memory (1,14).
Owing to the IL-1 ability in augmentation cell-mediated immunity and also significant role of TIR in mediating IL-1 downstream signaling pathway, the domain and its interaction could be of potential target in adjuvant development. The protein–protein interactions between TIR-TLR7 and MyD88 adaptor protein followed by TLR7 signaling of DCs have a critical role in activation of signaling pathway specific to IL-1 (3,15). Triggering the IL-1 signaling cascade of protein kinases through this interaction leads to the expression of genes involved in proinflammatory responses and production of type I IFNs. The secretion of type I IFNs from DCs resulting in the initiation of other signaling cascades promotes the activation of a number of IFN-stimulated genes, which have a role in the initiation of innate immune response by the activation and regulation of the cellular components of innate immunity such as natural killer cells and adaptive immune response. Engagement of TLRs can instigate a range of immunological events through the activation of type I IFNs and NF-κB, resulting in production of proinflammatory cytokines. TIR domain may induce effective immune responses through the activation of conserved cellular signal transduction pathways. The signaling pathway ultimately leads to the activation of NF-κB and IFN-I regulatory factors and regulation of B cell and T cell responses (6,27,28). Compared to the group immunized with inactivated IBD alone, induction of a higher level of the protective antibody, which lasts 7 weeks after vaccination with IBDV/TIR-TLR7, may improve both effector and memory immune responses. The response leads to enhanced specific antibody titers against IBDV as well as CTL levels.
As a result, the increased immunogenicity of the adjuvanted vaccine was because of the presence of TIR domain of TLR7, which is expressed on immune-related tissues and cells of chickens, the splenic B cells, and bursa of Fabricius. Understanding how TIR-TLR7 enhances the adaptive immune responses and conferred full protection against IBD is our great interest in future studies.
Footnotes
Acknowledgment
This study was supported by the Razi Vaccine and Serum Research Institute of Iran under Grant No. 2-18-18-9311.
Authors' Contributions
M.M.E. performed all the experiments and interpretation of data and S.S. contributed to project management. The authors participated meaningfully in the study and have approved the final article.
Author Disclosure Statement
No competing financial interests exist.