
Abstract
The human papillomavirus (HPV) vaccine has not been widely used in developing countries because of its high cost and multiple subtype restrictions. The present study aimed to develop an economical, convenient, and effective vaccine to produce neutralizing antibodies. Using late protein 1 (L1) from the HPV16 subtype as the target antigen (HPV16L1) and Pichia pastoris as the antigen release system, integrated P. pastoris expressing HPV16L1 (named yeast-HPV16L1) was prepared and vaccinated directly into mice by subcutaneous multipoint injection. After immunization was performed thrice, high titers (greater than 1:40,960) of specific anti-HPV16L1 antibodies were obtained in immune serum and were observed to continuously rise over time. The indirect hemagglutination test and indirect hemagglutination inhibition test were used to detect neutralizing antibody activity in vitro, and the results demonstrated the hemagglutination ability of the immune serum and the reduction in or loss of the hemagglutination ability if preneutralized antigen was added to the immune serum. The protection conferred by immune serum to tumor-bearing mice at the early stages was confirmed, but the neutralizing activity disappeared when the tumor reached a size of 1 mm3. The neutralization activity of the immune serum was confirmed both in vitro and in vivo.
Introduction
Human papillomavirus (HPV), which has over 130 different genotypes (including low-risk and high-risk types), has been shown to lead to proliferative lesions on human skin and mucosa (8,9). High-risk types, such as HPV16 or HPV18, promote the development of cervical cancer, which is the third leading cause of cancer death in women (6,10,32,35).
The HPV vaccine is the first anticancer vaccine that was developed in the world and is based on virus-like particles (VLPs) that are self-assembled from the late protein 1 (L1) of the virus, and the VLPs only result in immunogenicity against the native virus without any danger of infection. Vaccination with HPV16L1 VLPs in women can produce a high titer of anti-L1 neutralizing antibody, which can protect against HPV infection or HPV-associated cervical epithelial atypical hyperplasia (12,29). Although the HPV vaccine has been authorized to be widely used for women of child-bearing age in many developed countries, many women in developing countries still cannot obtain its benefits. Naturally, it is necessary to develop an alternative immunization formulation that could be used to induce high levels of neutralizing antibodies against HPV infection.
VLP vaccines have been produced by genetic engineering technology in several expression systems, such as yeast or insect cells (17,19,26). Due to its advantages in terms of easy cultivation, low-cost mass production, bioactive protein expression, posttranslational modification, and, more importantly, its lack of pathogenicity, yeast is considered to be an ideal organism for the engineering and industrialized production of medical recombinant proteins (18,40). The purified VLPs are obtained in a yeast expression system by sucrose cushion centrifugation, cesium chloride gradient centrifugation, or size exclusion chromatography, but these purification processes require many steps, are highly technical, and produce variable protein recovery (27). The difficulty of their preparation and the possible side effects limit the application of VLP vaccines. This study aimed to identify an alternative immunization method that used integrated yeast expressing HPV16L1 for the production of neutralizing antibodies without a need to prepare VLPs in vitro.
There have been some reports that direct immunization with yeast can efficiently stimulate the immune response in vivo (5,20,38), although the underlying immune mechanisms in yeast are not yet completely understood. Presumably, yeast attracts dendritic cells (DCs) through the interaction between mannan, which is expressed on the yeast cell surface, and mannose receptors (CD206) on DCs (4,33) or through the interaction of β-1,3-D-glucan, which is a cell-wall component in yeast with adjuvant potential, expressed on active yeast with mature DCs (25,27,28); in addition, yeast mannosylate proteins, which induce MHC-restricted antigen presentation and T cell activation in mammals, can attract DCs (7,24). For example, vaccination with Saccharomyces cerevisiae expressing tumor-associated antigens can induce antigen-specific T cell responses and protect animals against tumor challenge (39), and heat-killed recombinant S. cerevisiae exhibited no toxicity in clinical studies, which makes S. cerevisiae a potential vehicle for vaccines against cancer and infectious diseases (2,13). Other studies have also reported that whole recombinant yeast-based immunotherapy has been used to combat HIV-1, hepatitis B and C, and other pathogens (11,16,30).
Over the past 10 years, Pichia pastoris has been utilized to develop a novel and effective eukaryotic expression system, which has an enormous advantage because of its high expression yield that is more than 60 times greater compared with S. cerevisiae. pPIC3.5K, which is a yeast integrative vector that is fused with a foreign target gene, can be inserted into a chromosome of P. pastoris and will never be lost. In the present study, pPIC3.5K was selected for the construction of the recombinant plasmid pPIC3.5K-HPV16L1, which was to be transferred into P. pastoris. After cultivation and inactivation, whole recombinant P. pastoris expressing HPV16L1 (yeast-HPV16L1) was vaccinated directly into animals, and the HPV16L1-specific antibody levels and antibody neutralizing activities in vivo and in vitro were analyzed. Our work is a preliminary attempt to identify alternative immunization methods that could be used to produce neutralizing antibodies against HPV infection.
Materials and Methods
Reagents
The pPIC3.5K vector (Invitrogen, Carlsbad, CA), bacterial strain Escherichia coli-DH5α, and the yeast strain P. pastoris GS115 were preserved in our laboratory. The pMD18-T vector was purchased from TaKaRa (Japan). The restriction enzymes (BamHI, EcoRI, and BglII), T4 DNA ligase, DNA marker, prestained protein marker, and 2 × PCR Master Mix were purchased from MBI Fermentas (Burlington, Canada). The Endo-free Plasmid Maxi Kits were purchased from Qiagen (Germany). The HPV16L1 monoclonal antibody (Abcam; cat. ab69) and the horseradish peroxidase (HRP)-labeled goat anti-mouse IgG (Abcam; cat. ab205719) were purchased from Abcam (England). Yeast extract peptone dextrose (YEPD) medium, buffered minimal glycerol-complex medium (BMGY), and buffered minimal methanol-complex medium (BMMY) were purchased from Oxoid (England). A HiTrap™ heparin HP column was purchased from GE Health Care. The primers were synthesized by Sanbio-Tech (Beijing, China). All other reagents that were used were analytical reagents.
Cells and animals
Cervical cancer SiHa cells (HPV16-positive) (ATCC-TCHu113) were purchased from the Shanghai Cell Bank of the Chinese Academy of Sciences and grown in RPMI1640 medium supplemented with 10% fetal bovine serum (FBS). Four- to 5-week-old female BALB/c-nu mice [from the National Science Academy of the Center for Laboratory Animals, Shanghai, China, certificate no. SCXK (Shanghai) (2012-0002)] and 6- to 8-week-old female BALB/c mice [certificate no. SCXK (Shanghai) (2010-0044)] were used for experimental procedures according to protocols approved by the Medical Ethics Committee of Wenzhou Medical University and in accordance with the recommendations for the proper use and care of laboratory animals in China.
Construction of the pPIC3.5K-HPV16L1 recombinant plasmid
The amino acid sequence of HPV16L1 was obtained from GenBank (no. AY177679), and the DNA sequence corresponding to HPV16L1 was modified based on preferred codon usage in yeast by the JCat software (
Screening of positive strains expressing HPV16L1
The recombinant plasmid pPIC3.5K-HPV16L1 was amplified in E. coli-DH5α, extracted and purified with an Endotoxin-free Plasmid DNA Purification Kit according to the manufacturer's instructions, and then electrotransformed into GS115 competent cells using Gene Pulser (Bio-Rad) at 1.1 kV, 200Ω, and 25 mF after linearization by digestion with the restriction enzyme BglII. Minimal dextrose (MD) medium without histidine was used for the preliminary selection of positive clones, and positive clones on the MD plate were further verified by PCR (using the same primers as above). The positive clones with integration of the recombinant plasmid into their chromosomes were named yeast-HPV16L1.
After 1, 3, and 6 months of preservation and 1, 10, and 20 cell passages, the integrated yeast-HPV16L1 clones were collected to identify the stability of pPIC3.5K-HPV16L1 by PCR using the same primers as above.
Expression and identification of yeast-HPV16L1 clones
Identified positive clones of yeast-HPV16L1 were first cultured in YEPD medium and were then transferred into BMGY for continuous growth at 37°C for an additional 18 h. The yeast cells were harvested by centrifugation for 10 min at 4,000 r/min, and the precipitants were suspended in BMMY; absolute methanol was added every 24 h to express the integrated gene. On day 5, the cell precipitants were collected and extruded by a high-pressure cell crusher. The lysed cells in buffer (2.68 mM KCl, 1.47 mM KH2PO4, 8.1 mM Na2HPO4, and 0.33 M NaCl, pH 7.0) were dialyzed in 0.01% Tween-80 for 3 h at 4°C. After dialysis, the sample was loaded into a HiTrap Heparin HP column (GE Health Care) for purification, and a 0.6 M–1.8 M NaCl gradient was used for washing. The eluted solution was separated on a 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and blotted onto a polyvinylidene fluoride membrane. The membranes were blocked with 5% skim milk and probed with a HPV16L1 monoclonal antibody (1:6,000) for 2 h, followed by incubation with HRP-conjugated goat anti-mouse IgG (1:6,000) for 1 h. All blocking, incubation, and washing steps were performed in PBST (Phosphate Buffered Saline with 0.05% Tween-20). The blot was visualized by an Enhanced Chemiluminescence Detection Kit (Pierce, Rockford, IL) according to the manufacturer's instructions.
Immunization protocol
After culture for 5 days in BMMY, yeast with integrated pPIC3.5K-HPV16L1 (yeast-HPV16L1), yeast with integrated pPIC3.5K only (yeast), and yeast without integration (empty yeast) were heat killed in a 56°C water bath for 15 min. Six- to 8-week-old female BALB/c mice were randomly assigned (n = 5) to groups that received 5 μg (equal to 2 × 102 yeast cells) of yeast-HPV16L1, yeast, empty yeast, or phosphate-buffered saline (PBS; blank control). The immunization schedule was carried out by subcutaneous multipoint injection and was repeated thrice at 7-day intervals. Serum was collected from the tail vein of the mice after immunization at weeks 1, 3, 5, 7, and 9 for the evaluation of HPV16L1-specific antibody (IgG) production.
Antibody detection by enzyme-linked immunosorbent assay
HPV16L1-specific IgG in serum was measured by enzyme-linked immunosorbent assay (ELISA). Microtiter 96 flat-bottom plates (Corning, Lowell, MA) were coated overnight at 4°C with 100 μL (10 μg mL−1) purified HPV16L1, blocked at 37°C for 1 h with blocking buffer (5% skim milk in PBST), and subsequently incubated at 37°C for another 1 h with the serum samples (diluted 1:100). After washing with PBST, the plates were incubated with HRP-conjugated anti-mouse IgG diluted 1:3,000 in blocking buffer at 37°C for 1 h to detect the bound antibodies. After washing with PBST, the chromogenic substrate O-phenylenediamine (OPD; Sigma) and H2O2 were added, and the plates were incubated at 37°C for 10 min. The color development was determined at 492 nm using an automated ELISA reader (Bio-Tek ELx800). All samples were analyzed in triplicate.
The immune serum obtained at week 9 was serially diluted twofold and used to detect the titer of the HPV16L1-specific IgG according to the procedure described above. Each dilution was also analyzed in triplicate.
Dot immunobinding assay
The specific binding characteristics of the immune serum were confirmed by a dot immunobinding assay (DIBA). Four films of nitrocellulose membrane were individually treated with 10 μL (1 μg/μL) purified HPV16L1 and incubated at 37°C for 30 min. The membranes were blocked with blocking buffer overnight at 4°C. After washing, the membranes were incubated with anti-HPV16L1 McAb or immune serum for 2 h. Both the McAb and the immune serum were diluted 1:3,000. After washing, HRP-conjugated anti-mouse IgG (diluted 1:5,000) was added, and the membranes were incubated for 2 h to assess the serum immunobinding capacity. The spots were visualized by a Chemiluminescence Detection Kit according to the manufacturer's instructions.
Indirect hemagglutination and indirect hemagglutination inhibition assays
The indirect hemagglutination test (IH) and the indirect hemagglutination inhibition test (IHI) were used to assess the neutralizing activity of the immune serum. To fix and acidify the erythrocytes, fresh blood with 20 U mL−1 of heparin was washed with PBS containing 0.1% bovine serum albumin (PBS-BSA) five times and fixed with 2.5% glutaraldehyde for 1 h on ice. After washing with PBS-BSA, the erythrocytes were acidified with tannic acid in a 37°C water bath for 15 min and were then resuspended in PBS to yield a 2.5% (v/v) cell suspension. To sensitize the erythrocytes, the erythrocyte suspension was mixed with 1 mL of HPV16L1 antigen (20 μg mL−1) and 4 mL of PBS (pH 6.4) at 37°C for 30 min. After centrifugation at 1,000 r/min for 5 min, the precipitated erythrocytes were washed twice with 1% mouse serum (before immunization) in PBS (pH 7.2) to remove the nonspecific agglutinogen. Then, the 2% sensitized erythrocytes were resuspended in PBS (pH 7.2). To perform the IH test, after inactivation at 56°C for 30 min, the immune serum was mixed and incubated with an equal amount of 2% erythrocyte suspension at 37°C for 30 min to remove the nonspecific agglutinin. After centrifugation at 1,000 r/min for 15 min, the serum supernatant was collected and serially diluted twofold in a “U”-shaped plate (shown in Table 1). Then, 0.25 mL 2% sensitized erythrocyte suspension was added to each well and incubated at 37°C for 1 h (shown in Table 1). The appearance of the agglutinated erythrocytes was observed. To perform the IHI test, 0.25 mL of diluted HPV16L1 antigen was added into each well of a “U”-shaped plate, and then 0.25 mL immune serum was added (shown in Table 2). After coincubation at 37°C for 1 h, 0.25 mL of sensitized erythrocyte suspension was added and incubated at 37°C for 1 h (shown in Table 2), and the appearance of the inhibited agglutinated erythrocytes was observed.
Experimental Program Used for the Indirect Hemagglutination Test (ml)

Experimental Program Used for the Indirect Hemagglutination Inhibition Test (ml)

Anti-tumor testing of immune serum
SiHa cells (HPV16-positive) suspended in FBS-free RPMI1640 medium were used as the tumor cells, and an area on the back of each nude mouse near the right axilla was used as the inoculation site. The neutralizing activity of the immune serum in vivo was determined as follows. The nude mice were divided into four groups (n = 3): group 1, which was inoculated with a mixture of immune serum and SiHa cells; group 2, which was inoculated with immune serum after 3 days of inoculation with SiHa cells; group 3, which was inoculated with immune serum once tumors had reached 1 mm3 in size; and group 4, which was inoculated with SiHa cells only (control group). A total of 3 × 105 SiHa cells were injected into each mouse, and the immune serum was injected by local infiltrating injection (0.1 mL per mouse, thrice, 1 day interval). Every 2 days after inoculation with cells, the long diameter (represented by “a”) and the short diameter (represented by “b”) were measured by a Vernier caliper, and the tumor volume was calculated according to the formula V = ab2/2 (23). On day 36, all mice received anesthesia, and the tumors were separated and weighed.
Statistical analysis
All data were expressed as the mean measured value ± standard deviation (SD). All comparisons were tested for significance by one-way ANOVA. Differences for which p ≤ 0.05 were considered significant. The statistical analysis was performed using SPSS statistical software (SPSS, Inc., 1989–1999, Chicago, IL).
Results
Identification of recombinant plasmid pPIC3.5K-HPV16L1
The yeast-preferred, codon-optimized HPV16L1 gene was cloned into pPIC3.5K to generate the recombinant plasmid pPIC3.5K-HPV16L1 (Fig. 1A). A 1,572-bp fragment of AOX primer-HPV16L1 was amplified by PCR (Fig. 1B, lane 1), and a 1,518-bp fragment of HPV16L1 was identified by restriction endonuclease digestion with BamHI and EcoRI (Fig. 1B, lane 3). A sequencing analysis revealed the presence of the target HPV16L1 gene in the plasmid (data not shown), and the results indicated the successful insertion of HPV16L1 into the pPIC3.5K vector.
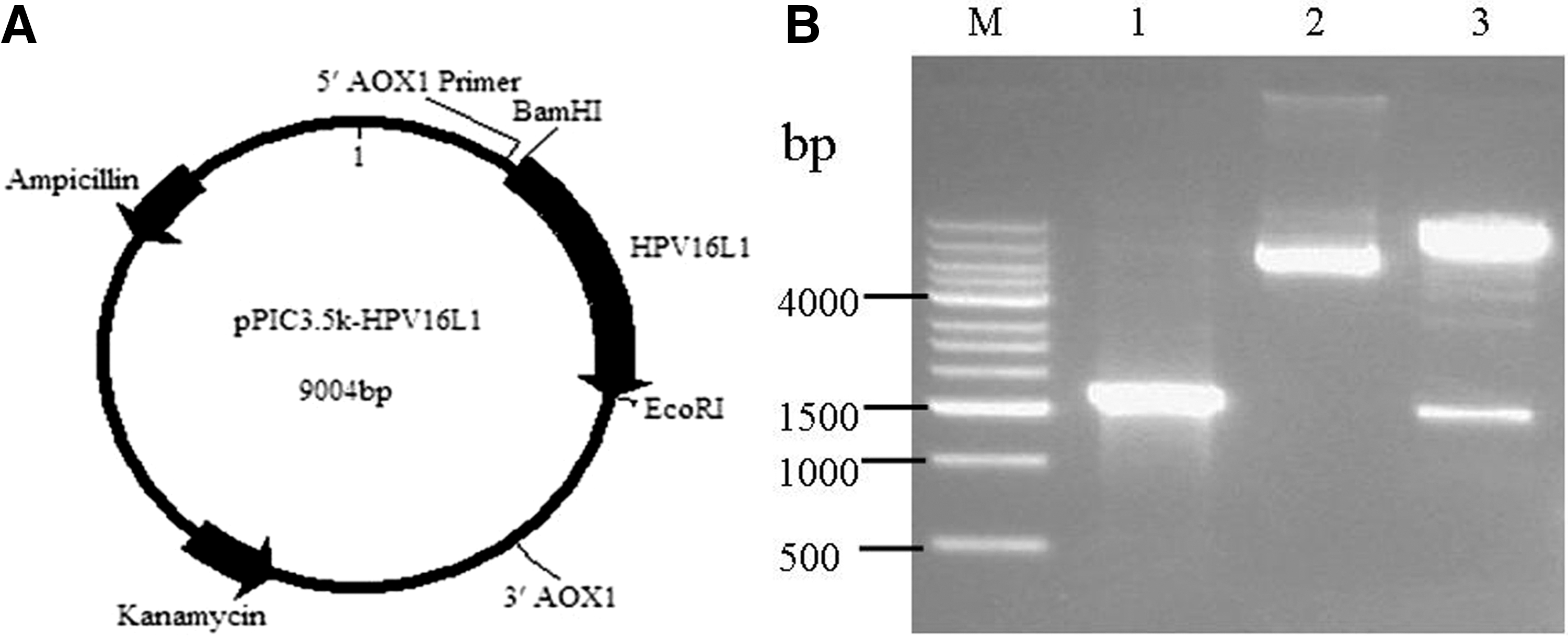
Construction and identification of the pPIC3.5K-HPV16L1 plasmid.
Identification of integrated recombinant yeast-HPV16L1
To exclude false-positive clones of yeast-HPV16L1 on the MD plates, PCR was used to identify the target HPV16L1 gene. A 1,572-bp fragment of AOX primer-HPV16L1 was amplified in a positive clone of yeast-HPV16L1 (Fig. 2, lane 3). Neither target gene was amplified from empty yeast in the absence of transformation (lane 2) or from pPIC3.5K-transformed yeast (lane 1). The stability of pPIC3.5K-HPV16L1 in P. pastoris after 6 months of preservation and 20 cell passages was confirmed by PCR. When checked at 0, 1, 3, and 6 months and after 1, 10, and 20 cell passages, the target gene of pPIC3.5K-HPV16L1 could always be amplified from yeast-HPV16L1 (Fig. 3).
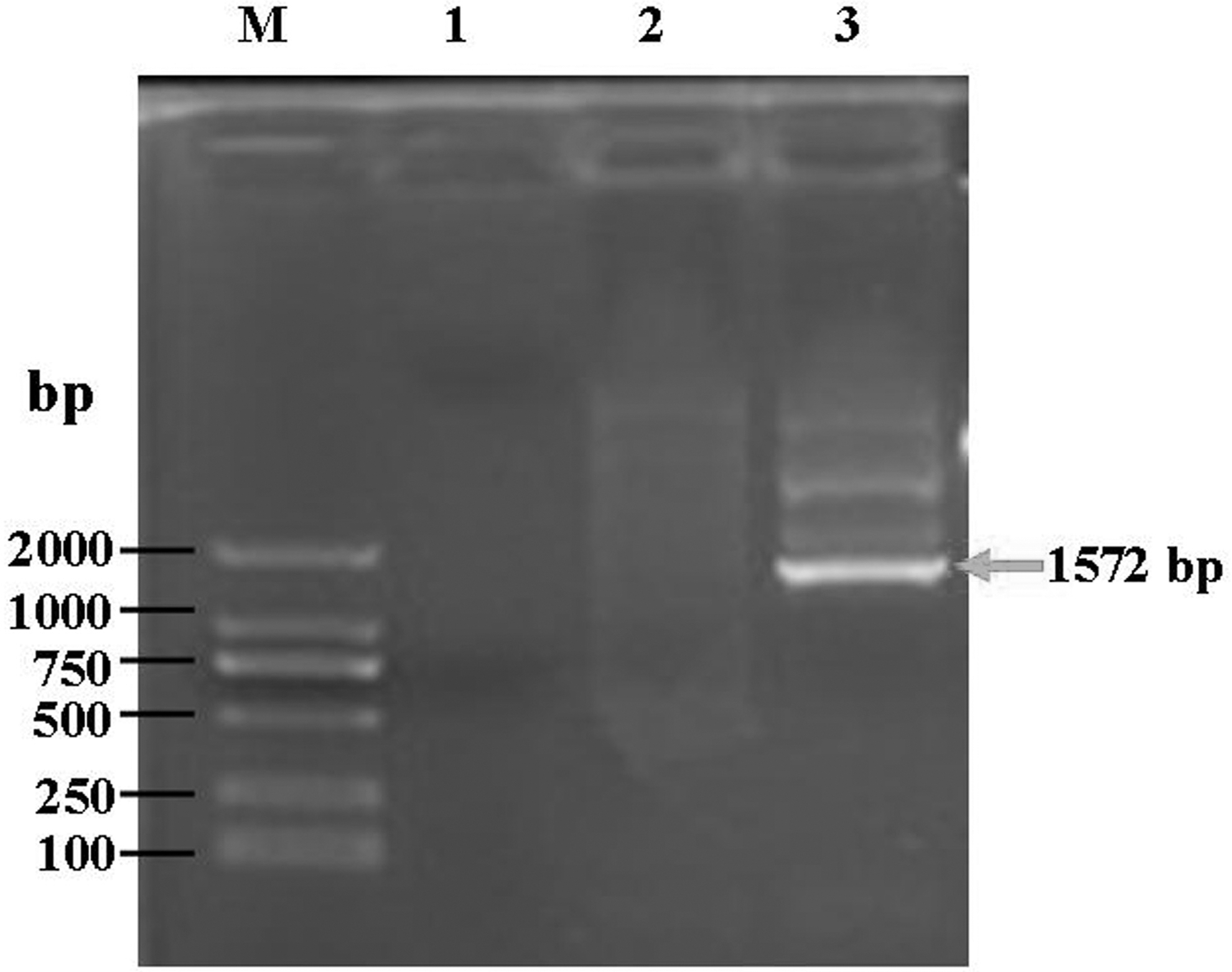
Verification of the integration of the HPV16L1 gene into a chromosome of Pichia pastoris by PCR. M, DNA marker; lane 1, empty yeast (without any transformation); lane 2, yeast (transformation with pPIC3.5K); lane 3, yeast-HPV16L1 (transformation with pPIC3.5K-HPV16L1). The arrow indicates the target gene fragment of HPV16L1.
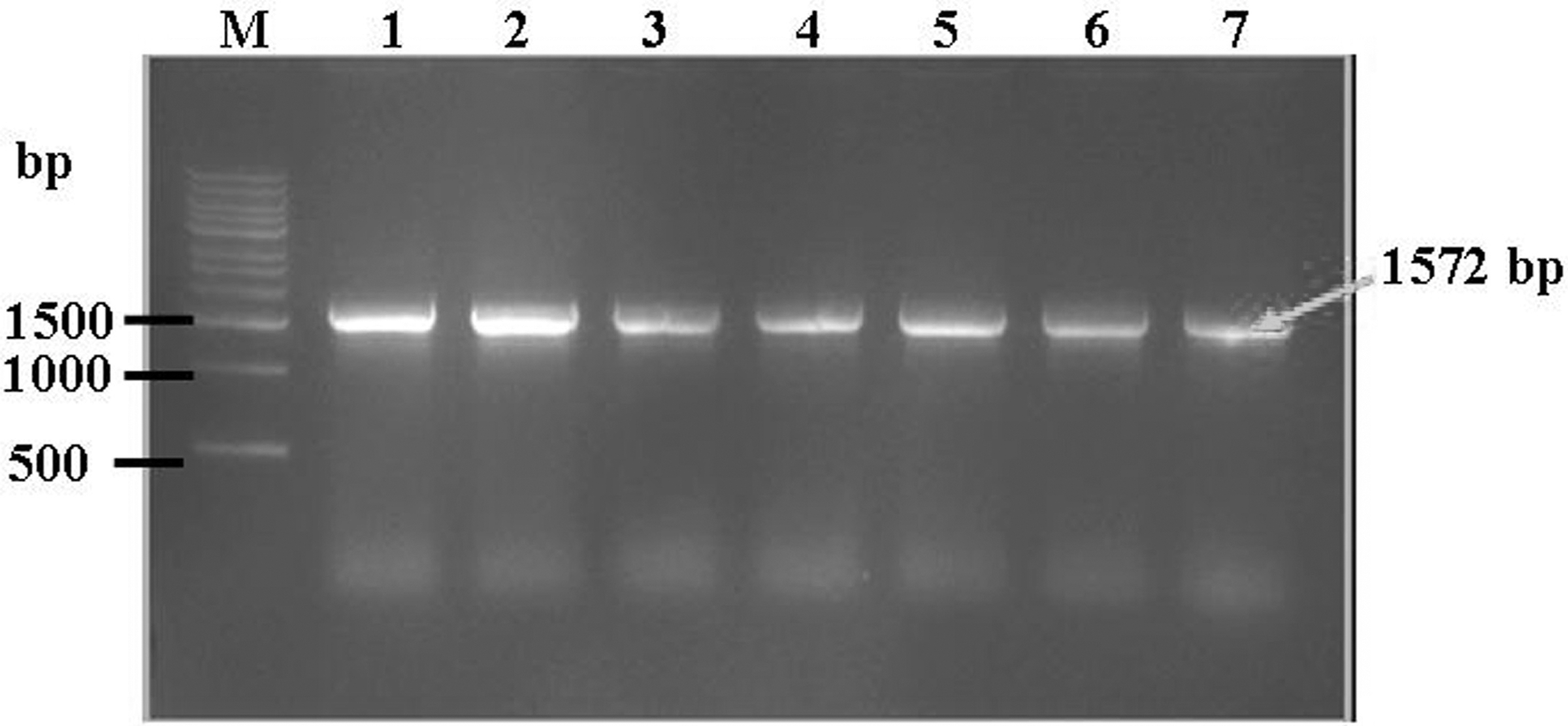
Verification of the stability of the integration of pPIC3.5K-HPV16L1 in P. pastoris. M, DNA marker; lanes 1–4, 0, 1, 3, and 6 months; lanes 5–7, 1, 10, and 20 generations. The arrow indicates the target gene fragment of HPV16L1.
Expression analysis of HPV16L1 in P. pastoris
Cultured yeast-HPV16L1, yeast, or empty yeast was extruded to release the expression products, and SDS-PAGE (Fig. 4A) and Western blotting (Fig. 4B) were used to detect the HPV16L1 target protein. A 57-kDa target protein band from yeast-HPV16L1 could be probed by an anti-HPV16L1 monoclonal antibody (Fig. 4B, lanes 3–4), while no corresponding protein bands were observed in empty yeast (without integration) (Fig. 4B, lane 1) or yeast (integration with pPIC3.5K only) (Fig. 4B, lane 2). The results showed the successful expression of HPV16L1 in integrated P. pastoris.
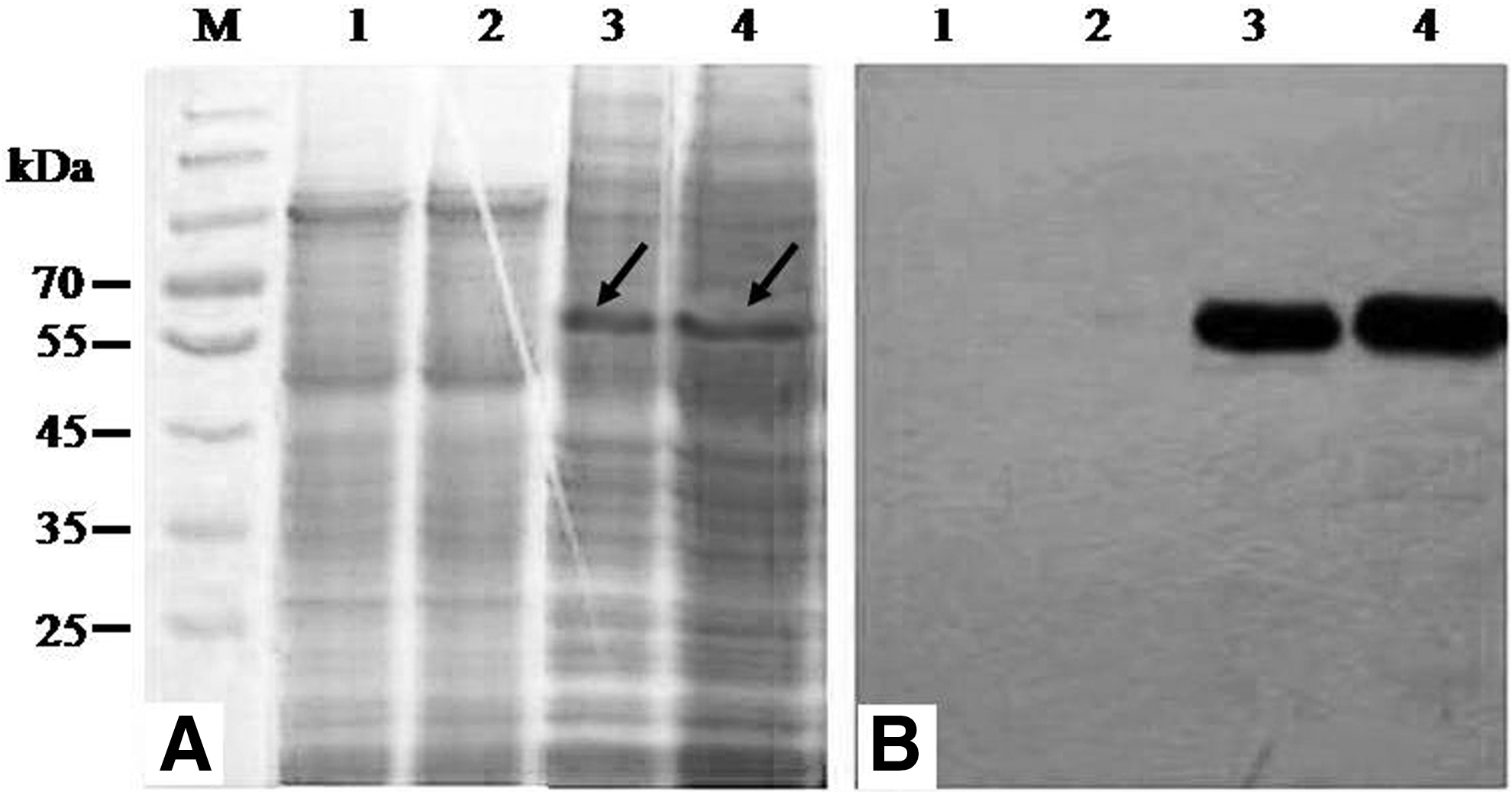
Analysis of HPV16L1 expression in recombinant P. pastoris.
Higher production of HPV16L1-specific antibody in immune serum
HPV16L1-specific antibody in immune serum with yeast-HPV16L1 began to be produced significantly at week 3 and increased over time, and the antibody levels were also significantly higher than those in the control groups of yeast or in PBS from week 3 to 9 (p < 0.05, Fig. 5A). The highest antibody production from yeast-HPV16L1 was obtained at week 9 (1.945 ± 0.083), but nonspecific antibodies were also produced in the yeast group from week 5 to 9 and were significantly increased compared to that in the PBS control group (p < 0.05) (Fig. 5A). The results revealed that a nonspecific immune response was also induced by the yeast, which can serve as an immune enhancement. At week 9, the maximum serum dilution of the specific antibodies in the yeast-HPV16L1 group was 1:40,960 (Fig. 5B), whereas the maximum dilution in the yeast group was almost equal to that of the yeast-HPV16L1 group (Fig. 5B). The serum dilution in the PBS group was always at the lowest level.
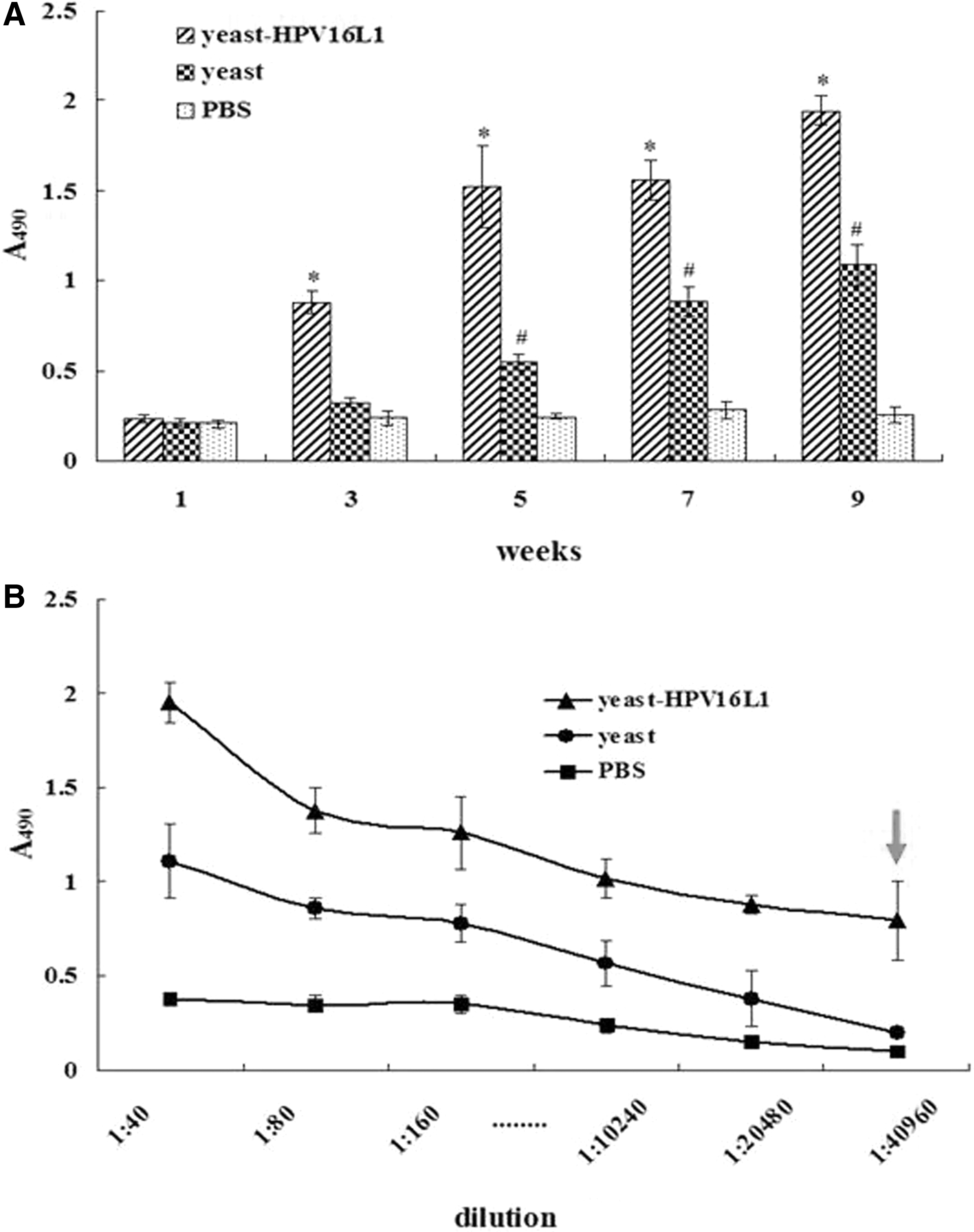
Detection of HPV16L1-specific antibody IgG in immune serum.
Antibody specificity for HPV16L1 determined by DIBA
The analysis of nitrocellulose films conjugated with HPV16L1 protein showed obviously combined dots upon blotting with immune serum from the yeast-HPV16L1 group or with HPV16L1 McAb (as a positive control); however, no combined dots were clearly observed when the films were blotted with immune serum from empty yeast or yeast (Fig. 6). The results further demonstrate the specificity of immune serum for HPV16L1.
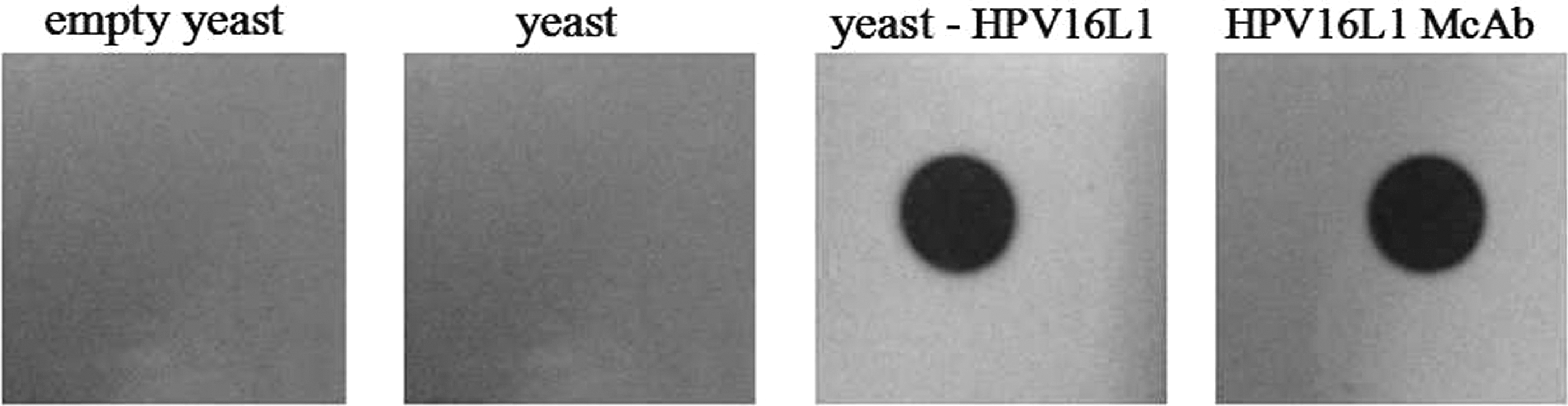
Analysis of antibody specificity to HPV16L1 in immune serum by immunodotting.
Neutralization activity of the immune serum in vitro
The neutralization activity of the immune serum in vitro was evaluated by IH and IHI assays. For the IH test, when diluted to 1:10,240 or 1:20,480, the immune serum from yeast-HPV16L1 could agglutinate half of the HPV16L1-sensitized erythrocytes (Fig. 7A); therefore, the hemagglutination unit was judged to be 1:20,480. For the IHI test, the agglutination activity of the immune serum decreased after it was coincubated with HPV16L1. The immune serum from the yeast-HPV16L1 group agglutinated half of the sensitized erythrocytes before coincubation with HPV16L1 protein that was diluted to 1:48.8 ng mL−1 or 1:24.4 ng mL−1 (Fig. 7B); therefore, the hemagglutination inhibition unit was judged to be 1:24.4 ng mL−1. Once the immune serum was incubated with more than 48.8 ng mL−1 of HPV16L1 protein, the majority of the erythrocytes precipitated instead of agglutinating. In addition, the neutralization activity of the immunized serum (Line 2 in Fig. 7) was nearly the same as that of the HPV16L1 McAb, for which there was only a difference of twofold dilution (Line 1 in Fig. 7) regardless of the results of the IH (A) or IHI (B) assays. The results indicated notable agglutination activity (i.e., neutralization activity) in the immune serum.

Assay of neutralization activity of the immune serum in vitro. Line 1: HPV16L1 McAb (positive control); line 2: immune serum from the yeast-HPV16L1 group.
Neutralization activity of the immune serum in vivo
For the control group (group 4), tumors began to form in mice on day 12 after inoculation with the SiHa cells and grew rapidly over time, whereas the tumor sizes of mice in both group 1 (co-inoculation with immune serum and SiHa cells) and group 2 (3 days of inoculation with cells followed by inoculation with immune serum) were always smaller than those of mice in the control group at any check point (p < 0.05). The results showed a good neutralizing effect of the immune serum in vivo. However, there was no significant difference in the tumor size between group 3 (after 1 mm3 tumor size was reached, followed by immune serum) and the control group (p > 0.05), which means that once the tumor grew to ∼1 mm3, the serum neutralizing effect disappeared (Fig. 8A). On day 36 after inoculation, the tumor tissues were isolated and weighed. The tumor weights in mice from both group 1 and group 2 were lower than those in mice from group 3 or the control group (Fig. 8B), which also confirmed the neutralization effect of the immune serum.
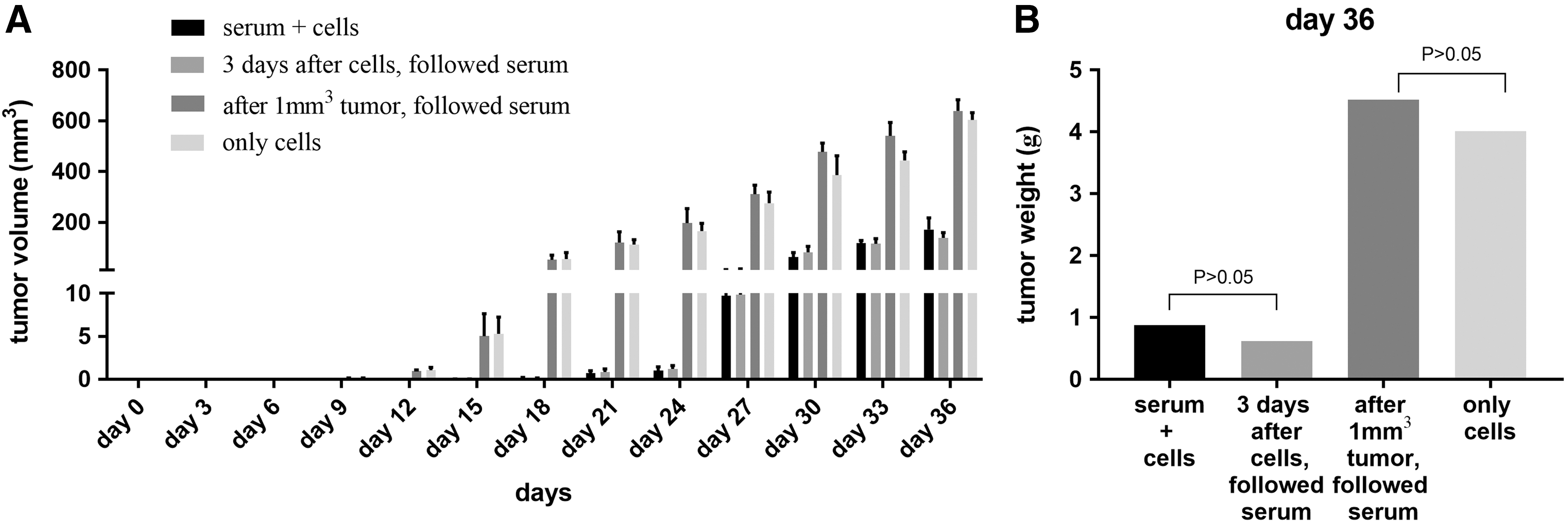
Assay of immune serum neutralization activity in vivo.
Discussion
The methanol-yeast expression system is an exogenous protein production system that was rapidly developed in recent years that can use methanol as the sole carbon source and can utilize several yeast species, including Hansenula polymorpha, Candida bodinii, and P. pastoris. Among these, P. pastoris is a widely used eukaryotic host for producing recombinant proteins with obviously high expression characteristics (21,31,36) and that has a production capability that could yield from 0.01 to 4.88 g L−1 protein (1,14). For example, larger quantities of epidermal growth factor were produced (450 mg L−1) in P. pastoris (∼60-fold enhancement) compared with S. cerevisiae (7.4 mg L−1) (3,18). The P. pastoris expression system has many advantages over bacterial or mammalian systems, such as the existence of powerful and tightly regulated methanol-inducible alcohol oxidase I (AOX I) promoters, the ability to grow quickly and economically on defined media at high cell densities, and the capability to produce many of the post-translational modifications performed by eukaryotic cells, including proteolytic processing and folding, disulfide bond formation, and glycosylation (15,34,37).
Until now, more than 40 kinds of exogenous proteins have been expressed successfully in P. pastoris, and in recent years, the expression of dozens of heterologous gene products has been reported annually in P. pastoris. Compared with mammalian cells, P. pastoris does not require a complex growth medium or culture conditions and can be manipulated genetically as easily as E. coli, which also has a eukaryotic synthetic protein pathway. For this reason, P. pastoris is considered to be one of the best and most widely used exogenous gene expression systems.
Some researchers have confirmed that immunization with whole recombinant yeast expressing a target antigen could induce efficient and specific immune responses in experimental animals. For example, Bian et al. (2010) reported that whole recombinant H. polymorpha expressing hepatitis B virus surface antigen (yeast-HBsAg) can enhance HBsAg-specific Th1 and Th2 immune responses and can induce potent HBsAg-specific lymphocyte proliferation and cytotoxic T lymphocyte responses, while in the alum group (HBsAg + alum) only enhancement of the Th2 immune response was observed, suggesting the possibility of developing yeast-HBsAg as an effective vaccine that could be used to control HBV infection. Another study by Stubbs et al. (2001) showed that whole recombinant S. cerevisiae used to produce a model tumor Ag was effective at inducing CD8+ T cell-mediated tumor protection in vaccinated mice, and the whole yeast was shown to be avidly internalized by murine DCs in vitro, leading to DC maturation and IL-12 production. To translate these important findings from the murine to the human system, Barron et al. (2006) investigated the interactions between whole recombinant yeast and human DCs, and their results suggested that S. cerevisiae could have potent adjuvant effects on human DCs and that recombinant yeast-derived antigens are processed by human blood DCs for MHC class-I cross-presentation. Considering the above facts, we wish to use recombinant yeast that express an HPV target antigen to produce a specific neutralizing antibody.
Because codon preference is closely related to the efficient expression of exogenous proteins in heterogeneous biological cells, in the present study, to improve the expression of HPV16L1 in P. pastoris, our first goal was to optimize the HPV16L1 gene codons based on the codon bias of P. pastoris. To achieve stable integration, the pPIC3.5K shuttle plasmid was selected for constructing the combined pPIC3.5K-HPV16L1 plasmid. The experimental results showed that effective HPV16L1 expression by the integration of pPIC3.5K-HPV16L1 in yeast was obtained, and the good stability of HPV16L1 was confirmed after long growth times and several passages in yeast.
Then, the constructed integrated recombinant yeast-HPV16L1 was directly vaccinated into mice to evaluate its ability to induce an immune response. The results showed that a higher level of specific antibody to HPV16L1 was obtained in immune serum and could steadily increase until week 9 (final detection time). The highest antibody titer was shown to reach more than 1:40,960 for yeast-HPV16L1, and the immunodotting experiments also confirmed the antibody specificity against HPV16L1. These experiments suggested the good immunogenicity of yeast-HPV16L1.
The goal of immunization with the HPV vaccine is to produce a specific protective antibody (a so-called “neutralizing antibody”) to prevent infection by HPV, which is a key indicator used to evaluate the vaccine effects in laboratory and clinical applications. In this study, we detected the neutralizing activity of the antibodies in immune serum both in vitro and in vivo. The IH and IHI experiments in vitro showed that the agglutination titer in immune serum with yeast-HPV16L1 is 1:20,480, while that in serum with HPV16L1 McAb is 1:40,960; the agglutination inhibition titer of the antigen in serum with yeast-HPV16L1 is 1:24.4 ng mL−1, while that in serum with HPV16L1 McAb is 1:12.2 ng mL−1. It is obvious that the antibody induced by yeast-HPV16L1 could exhibit neutralizing activity that is similar to that of the antibody induced by the mAb. For in vivo experiments, because of the strict species specificity for HPV, an infected animal model cannot be established (22). In this study, nude mice were used to establish an HPV16-positive tumor model for further verification of the neutralizing activity. The results showed that the immune serum could exhibit neutralization effects early in infection, but once the tumor grew to ∼1 mm3, this neutralization effect disappeared, which indicated that neutralizing antibodies may not reach the tumor germinal center. Although the immune serum failed to achieve complete protection, we thought that the number of inoculation cells far exceeded the amount of natural infection (but if there were not enough inoculation cells, a tumor was unable to form in vivo). There was no HPV16L1 monoclonal antibody designed to be used as a positive control in the test, and the immune serum neutralization effect was still obvious compared with that in the control group (without serum intervention).
In conclusion, our study provides preliminary work that could be used to develop another available immunization method based on the use of P. pastoris integrated with HPV16L1. At present, the HPV vaccine cannot be used by women who are still exposed to the danger of being infected. The immunization method introduced in this study could be useful as an alternative for practical applications, and a further attractive possibility may involve the cointegration of multiple target antigens into P. pastoris to deal with infection by different HPV subtypes.
Footnotes
Acknowledgment
This work was supported by the Public Welfare Foundation of Zhejiang Province, China (LGF18C010004).
Human and Animal Rights
All experimental protocols were conducted according to China Guidelines for Animal Care and were approved by the Medical Ethics Committee at Wenzhou Medical University.
Author Disclosure Statement
All authors declare that they have no conflicts of interest.