
Abstract
CD81 serves as an immune modulator, playing its role in tumor growth and metastasis of hepatitis C virus (HCV)-mediated hepatocellular carcinoma (HCC). CD81 serves as a coreceptor of viral entry and is found to be enriched in exosomes. HCV E2 protein when associated with CD81 may be responsible for B cell lymphoproliferative disorders, as extrahepatic manifestation. Studies predict that HCV association with exosomes, leads to the establishment of persistent infection, through immune evasion. Herein, we confirm the association of HCV particles with CD81+ exosomes. Breifly, exosomes were enriched from peripheral blood of chronic HCV patients who have developed HCC. Sideways, exosomes were also enriched from peripheral blood of healthy individuals, who exhibited normal liver function test profile and had no known infection. Isolation of subpopulation of CD81+ exosomes was performed through immunocapture, followed by detection using FACS. Scanning electron microscopy confirmed the physical association of a fraction of exosome with HCV. CD81+ exosomes from chronic HCV patients with HCC were more granulated and larger when compared with those enriched from a healthy individual and HCV RNA was also detected in enriched fractions of CD81+ exosomes from HCV-positive HCC patients only, through real-time quantitative polymerase chain reaction. We concluded that CD81+ exosomes carry HCV particles and the association plays a pivotal role in establishing persistent infection, through immune evasion, thus leading to HCC progression. Exosomal CD81 and its interacting proteins might, therefore, serve as a potential prognostic marker and therapeutic target in HCV progression mediated by active HCV infection.
Introduction
Tetraspanins are localized in microdomains, known as tetraspanin-enriched microdomains, on cell surface membrane of many cell types (14). They are transmembrane proteins that interact with integrins and other cell receptors, leading to different signaling cascades that involve the immune regulation of body and motility of cells. This includes interaction with the complement system, leukocytes, B cells, and T cells (17). The structure of tetraspanin superfamily includes two extracellular domains (EC1 and EC2) and four transmembrane domains (TM1, TM2, TM3, and TM4) (32). EC2 is the larger extracellular domain, which is responsible for interaction with majority of proteins; these include interaction of EC2 of CD81 with E2 protein of hepatitis C virus (HCV) (8,11). Many tetraspanins, including CD81, CD82, CD9, and CD63, are usually enriched in exosomes (5). However, since CD81 is an essential receptor of HCV, the “Trojan Horse” theory of viruses hijacking cellular mechanism of cellular pathways, including exosomes, for immune evasion and establishment of persistent infection is convincing (19,26). Immunomodulation by CD81 is observed both in viral hepatitis and cancer. These includes: (i) increased expression of CD81 in HCV-mediated B lymphocyte disorder (9,25); (ii) immunomodulation of plasmacytoid dendritic cells (7,10,13,19,23,31); (iii) lower, humoral IgG response to HCV associated with exosomes (1,7,22); and (iv) immunomodulation of T reg cells and myeloid-derived suppressor cells in tumor progression (27,28).
It has been reported that HCV is associated with CD81 in exosomes through E1 and E2 interactions. The two models proposed for this association are: (i) surface association on the external, and (ii) cargo association as internal, to either extracellular vesicle (EV; mainly exosomes) or low-density lipoprotein (8,20). Evidence has mounted that partial or full-genome HCV RNA present in exosomes released from Huh 7.5.1-infected cell line is able to successfully infect neighboring cells (6). This mode of viral genome transmission, is therefore, hypothesized to spread the virus in hepatic tissue (15). The mode of viral transmission have also been confirmed in vivo as well, mainly mediated by blood as a media of exosome spread carrying HCV-negative strand as a cargo, in both nonresponders and treatment-naive patients. Other than the genome and subgenome of HCV, viral proteins of HCV are also found to be enriched as cargo of exosomes (18,23).
Soluble CD81 present in exosome fraction was also observed to be corelated with serum ALT levels in HCV-infected patients. It was found that CD81 levels in exosomal fraction decreased in patients that showed sustained virological response (SVR), in comparison to treatment-naive and patients resistant to interferon therapy. Both SVR and healthy individuals gave similar levels of ALT and soluble CD81 (29).
Furthermore, an in vitro study confirms the secretion of HCV through HRS-dependent pathway. HRS-dependent pathway is an important juncture between both HCV secretion and normal exosome biogenesis pathway. However, both processes do not solely depend on HRS pathway; their levels of secretion in media did decrease on knocking down the HRS pathway (26).
Exosomes are enriched with microRNA (miRNA) and Ago proteins that stabilize them. miR-122 and Ago 2 have found to be associated with HCV genome not only inside cells, but also inside exosomes, for genomic stability and translation induction. HCV genome is also found to be associated with HSP90, enriched in the exosome complex. On inhibition of miR-122 and HSP90, exosomal secretion was reduced in Huh 7.5-infected cell lines, suggesting them as therapeutic targets (6).
Although many studies point toward the association and transfer of HCV using exosomes for spread and immune evasion, there are not many studies in vivo, which establish exosomal mode of transmission in chronic HCV infection. Understanding the role of exosomes/HCV complex in the mechanism of hepatocellular carcinoma (HCC) progression, persistent infection and extrahepatic manifestations owing to exosomes, shall help in tackling the HCV-caused health burden in a better way. These extrahepatic manifestations include transmitting HCV replication-competent genome to the peripheral blood mononuclear cell (PBMC), lymph nodes, and central nervous system of chronically infected patients (7). PBMCs infected with HCV in turn can be transferred from mother to fetus, leading to complications, even if the free viral particle is not being transferred (3). Keeping their importance in mind, we have isolated and detected CD81+ exosomes from peripheral blood of chronically infected HCV patients at different stages of HCC progression. The association of HCV with exosomal CD81 in disease progression shall further help in establishing its role in the development of HCV persistent infection leading to HCC.
Materials and Methods
Clinical sampling of peripheral blood of chronic HCV patients and enrichment of exosomes
We obtained ethics approval for clinical sample collection as well as handling them in BSL 2 conditions both from Rawalpindi Medical University, Rawalpindi, Pakistan and Atta-ur-Rahman School of Applied Biosciences (ASAB), National University of Sciences and Technology (NUST), Islamabad, Pakistan. Peripheral blood of both healthy controls and HCV-positive HCC patients was collected in 3.2% sodium citrate vacutainers, after informed consent. The samples were then transported on ice from the Centre of Liver Disease and Digestive Diseases (CLD) at Holy Family Hospital (HFH), Rawalpindi to ASAB and processed within 2 h of collection, for enrichment of EVs. Over a span of 6 months, a total of 40 samples were collected, among them 25 samples were of HCV-positive HCC cases and 15 were healthy controls. HCC was staged in accordance to Barcelona-Clinic Liver Cancer (BCLC) staging by consultant gastroenterologists at CLD, HFH; whereas HCV serology and active infection was confirmed by enzyme-linked immunosorbent assay (ELISA) followed by quantitative polymerase chain reaction (qPCR). Any patient that was coinfected with Mycobacterium tuberculosis, hepatitis C virus (HBV) and HIV was excluded from the study. Patients who were recommended blood transfusion or had received blood transfusion were also excluded from the study. Furthermore, complete blood cell count and liver function tests were performed as routine investigation at HFH. Healthy controls included in the study had normal blood cell count; normal liver functions tests; and were negative to HIV, HCV, and HBV ELISA tests. The tests for inclusion of healthy controls were conducted at the Atta-ur-Rahman Diagnostic Laboratory at ASAB, NUST. Clinical characteristics of patients and healthy controls are summarized in Table 1.
Summarized Clinical Characteristics of Participants of Study
BCLC, Barcelona-Clinic Liver Cancer; ELISA, enzyme-linked immunosorbent assay; HCC, hepatocellular carcinoma; HCV, hepatitis C virus; SD, standard deviation.
Preparation of platelet-poor plasma and enrichment of exosomes
Enrichment of EVs was optimized at Virology Laboratory, ASAB, with certain modifications to published protocols (26,27). Briefly, 10 mL of peripheral blood was transferred from 3.2% sodium citrate vacutainers to 15-mL sterile centrifuge tubes and centrifuged at 3,200 × g for 30 min at 4–8°C to obtain platelet-rich plasma, pelleting out buffy coat of mononuclear cells and red blood cells. Two third of platelet-rich plasma was transferred into a sterile 2-mL centrifuge tube and spun at 16,000 × g for 20 min at 4–8°C. The supernatant of platelet-poor plasma (PPP) was further processed, whereas the pellet of cellular debris and platelets was discarded. To obtain smaller EVs, the supernatant was then filtered across 0.20 μm pore size nylon syringe filter, allowing only vesicles of 200 nm and below to pass. To pellet down the larger EVs, including smaller apoptotic bodies and microvesicles, the filtrate was centrifuged at 23,000 × g for 25 min at temperature of between 4°C and 8°C. The pellet was discarded, the supernatant aspirated, and aliquoted. To each aliquot of 1 mL, 325 μL of 40% sterile PEG 4000 stock was added, to make 13% PEG final concentration in PPP, vortexed, and incubated overnight at 4°C. Next morning, the samples were vortexed and spun at 5,000 × g at 4°C for 20 min. The obtained pellet was resuspended in 0.1% bovine serum albumin (BSA) in 1× phosphate-buffered saline (PBS) and used in further downstream processing or stored at −80°C, immediately.
Scanning electron microscopy of enriched exosomes
Scanning electron microscopy (SEM) is used to visualize nanostructures, including exosomes and HCV particles. The samples of exosome-enriched pellet from HCV-positive HCC plasma and plasma of healthy controls were resuspended in 600 μL of 0.22 μm filtered 10% Dulbecco's modified Eagle's medium (DMEM) to make 1 mL of resuspension, followed by addition of 1 mL dimethyl sulfoxide (DMSO). The samples were then vortexed slightly for equal distribution of the exosomes. The samples were fixed onto glass slides, washed prior by acetone and ethanol under sonication. Two percent glutaraldehyde and 4% paraformaldehyde were used in combination as fixation agents. The samples and fixative agent mixture were left on the slide for 20 min, followed by washing off the excessive fixative with PBS for 15 min. The fixed samples were dehydrated by consecutive washing with ascending ethanol concentration gradient: 10%, 30%, 50%, 70%, 90%, and 100%. The slides were then air dried, carbon/gold coated, and analyzed within 2 days.
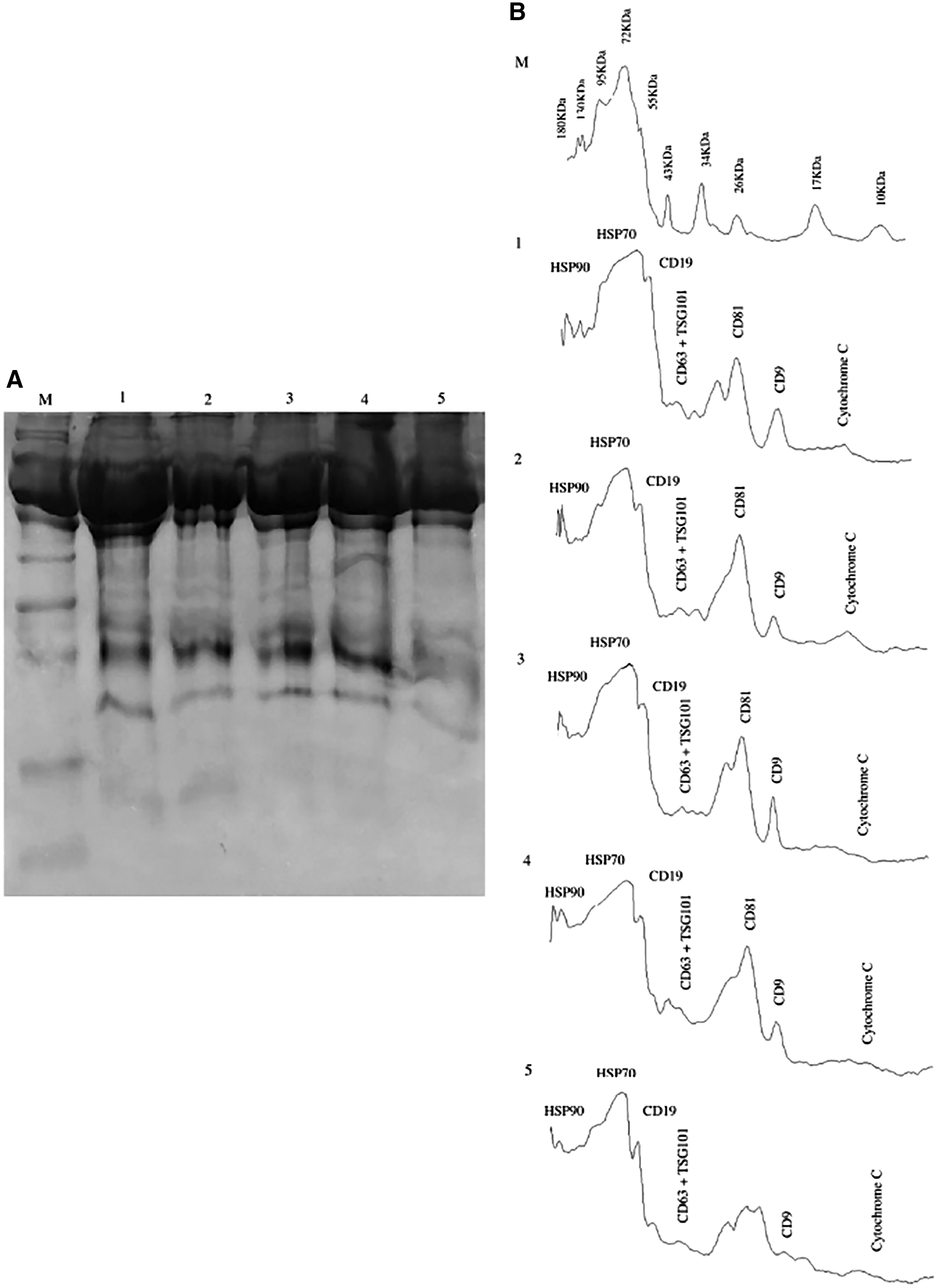
Presence of CD81 in enriched exosomes by sodium dodecyl sulfate/polyacrylamide gel electrophoresis
The confirmation of CD81 protein in the enriched pellet of exosomes was performed by one-dimensional (1D) sodium dodecyl sulfate/polyacrylamide gel electrophoresis (SDS-PAGE). For total protein isolation, the EVs were lysed with RIPA buffer (500–1,000 μL, depending on the size of pellet), containing PMSF as protease inhibitor, by incubation on ice for an hour, followed by water bath sonication on medium sonication at 35°C (JAC Ultrasonic; Jinwoo) for 5 × 3 min. The lysate was then centrifuged at 13,000 × g for 3 min at 4°C to remove cellular debris in the form of a pellet. The total protein in supernatant was then quantified through Branford method and 25 μg of total protein was resolved on 12% SDS-PAGE.
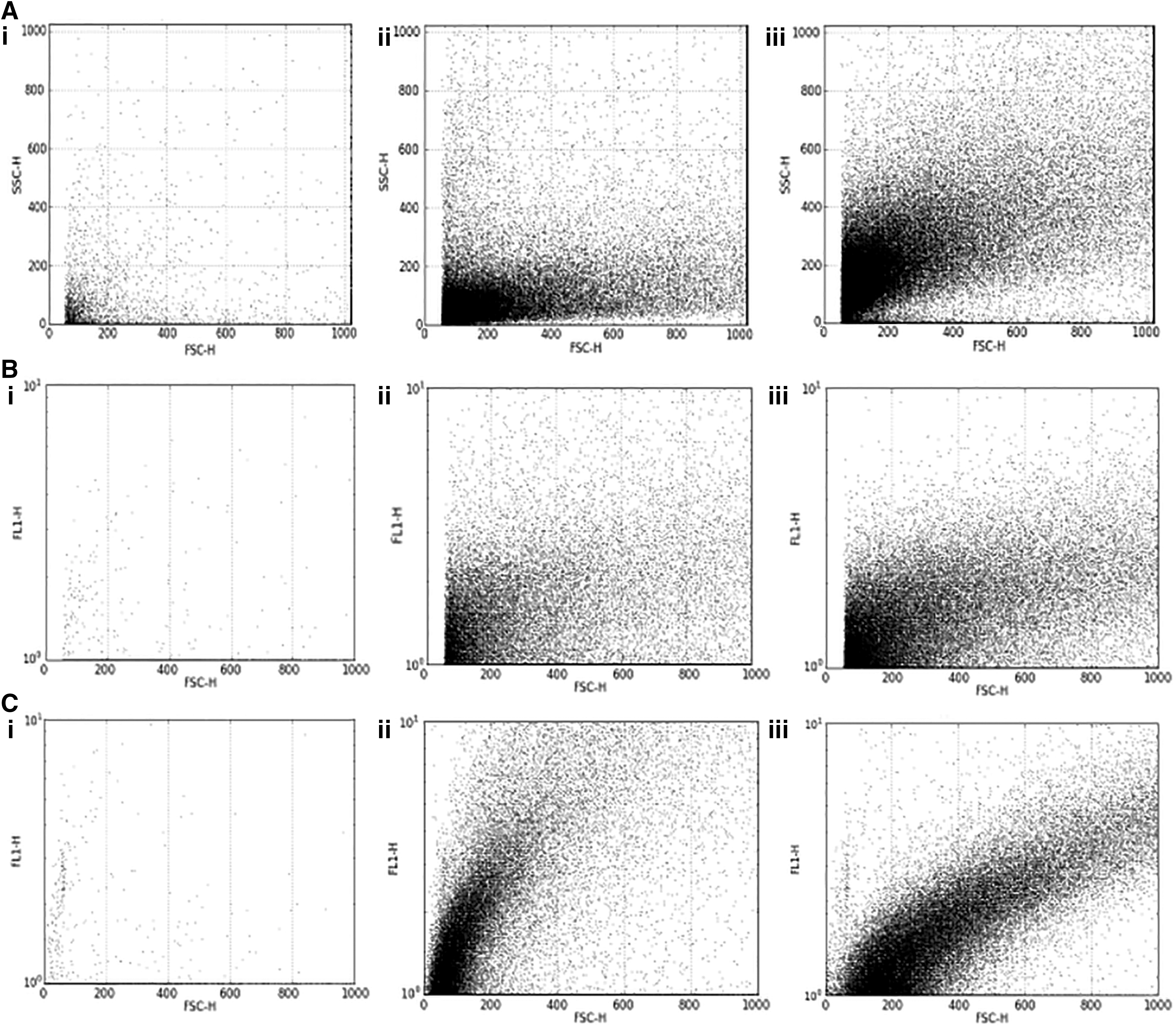
Detection of CD81+ exosome by flow cytometer
For the isolation and detection of a CD81+ subpopulation of exosomes, the approach used was: immunoprecipitation with CD81 antibody-conjugated Dynabeads (Exosome-Human CD81 Flow Detection Kit; Invitrogen), followed by flow cytometric detection with FITC conjugated. According to the manufacturer's instruction, 40 μL of washed CD81 coupled Dynabeads were added to 200 μL of exosome-enriched pellet, resuspended in 0.1% BSA in PBS, and incubated at 4°C for overnight with tilting and rotation. Next morning, the samples were briefly spun at 14,000 × g at 4°C, followed by washing of unbound exosomes with 300 μL of assay buffer on strong magnet. The pellet was then resuspended in same 100 μL of assay buffer and stained with FITC-conjugated antibody. Antibody titration was performed for initial optimization, using 0.2, 0.5, 0.7, 1, and 1.2 mg/mL of both CD81-TAPA FITC-conjugated antibody (Abcam) and IgGκ-1 FITC-conjugated antibody (Abcam) as isotype control. One milligram per milliliter of test antibody and isotype control were found to be optimum. Both control and positive were incubated for 45 min in the dark at room temperature. After staining, the beads were rewashed with assay buffer on magnet and resuspended in 1 mL of assay buffer and 1 mL of 7% paraformaldehyde in FACS tubes. The sample and isotype control were then detected by flow cytometry (FACSCalibur; BD Biosciences).
Detection of HCV genome in CD81+ exosome fraction
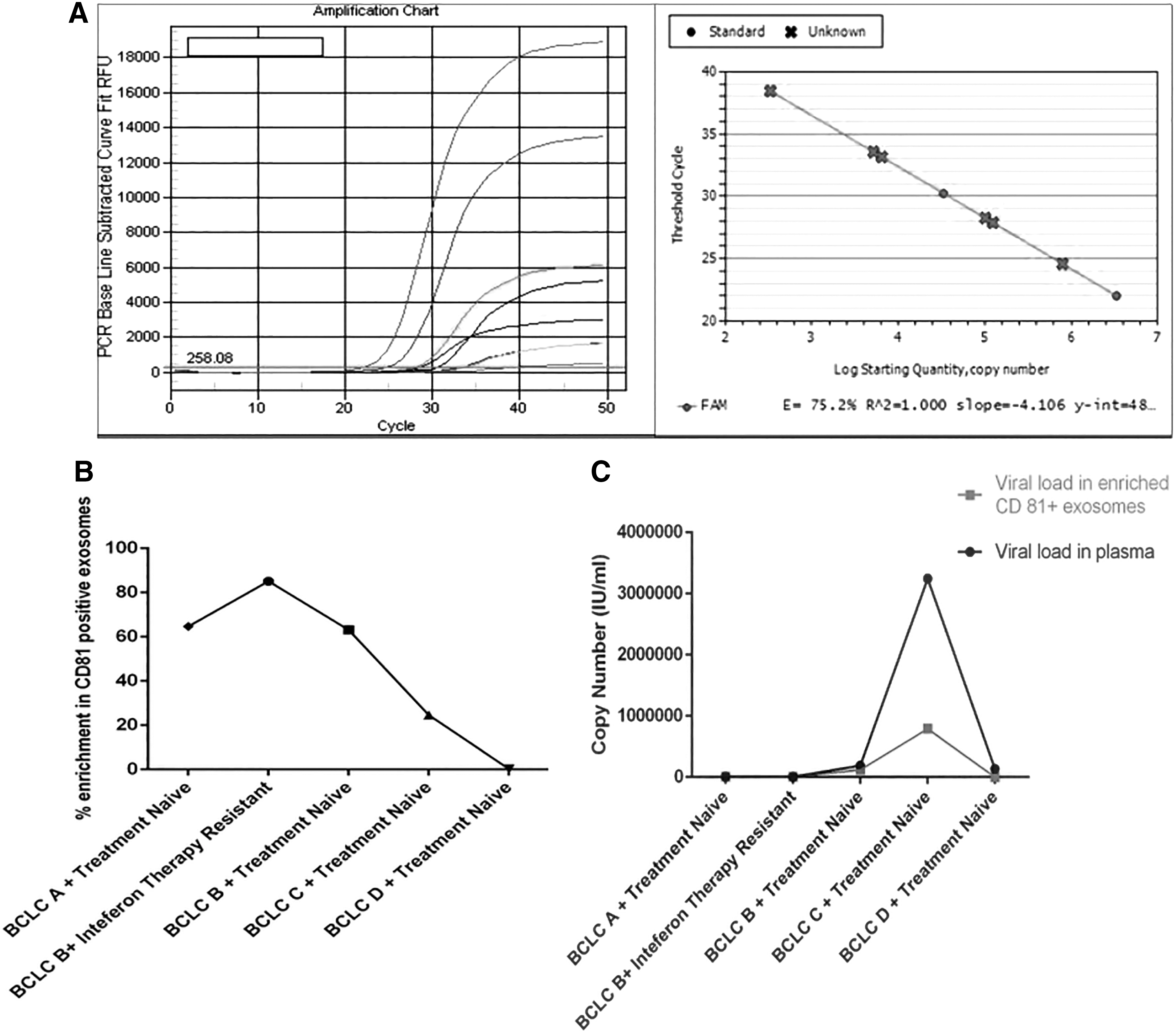
The viral RNA was isolated from CD81+ exosomes after immunoprecipitation using the NucleoSpin® RNA Virus Mini-Kit (Machery-Nagel) according to the manufacturer's instruction. Briefly, to 100 μL of resuspended dynabead-CD81+ exosomes, 600 μL of lysis buffer containing carrier RNA was added. This mixture was vortexed and incubated for 5 min at 70°C temperature. The lysate was separated from the beads on strong magnet. Around 600 μL of absolute ethanol was added to the lysate to enhance binding of RNA to the column filter. One thousand and two hundred microliters of this mixture was loaded onto the column filters, spun briefly at 8,000 × g and the filtrate was discarded. The column was washed with 500 μL of wash buffer and spun at 8,000 × g for a minute. The filtrate was discarded, the column was rewashed twice with concentrated wash buffer at 8,000 × g for 1 min, and then at 11,000 × g for 5 min, respectively. After the filtrate was disposed, 20 μL of elution buffer or RNase-free water was added unto the filter to elute the bound RNA from the column by spinning at 11,000 × g for 5 min. The isolated RNA was stored, temporarily, at −20°C.
The RNA eluted was then used for HCV RNA detection, using the GeneProof HCV PCR Kit (Life Technologies), according to the manufacturer's instructions. Briefly, 40 μL of master mix was added to each: 10 μL of isolated viral RNA, 10 μL of two calibration standards, and 10 μL of known viral RNA concentration as positive controls. The primers with FAM fluorophore were specific to conserved sequence present in 5′ untranslated region of HCV RNA genome, of HCV 1–7 genotypes, and calibrators. Whereas, the HEX-labeled primers were specific to internal standard (GAPDH was used as housekeeping). The tubes were placed in Bio-Rad iQ5 Real-Time PCR system and parameters were set in accordance to the manual. The automated threshold was set using the inbuilt Bio-Rad iQ5 software. The viral titer, in both copy number and IU/mL, were calculated based on standard curve built using calibrators and known HCV concentration. The samples were considered to be positive only if they gave detection above the threshold in FAM channel, whereas, the internal controls were positively detected in HEX channel for PCR efficiency.
Predicted protein interaction among HCV-encoded proteins and exosomal proteins
STRING 10.5 was used to identify the interaction network of CD81 with other cellular proteins and their biological pathways. Whereas, STRING Viruses Version 10.5 was used to study the interaction of HCV viral proteins with CD81, which is known to be expressed in both the exosomes found in circulation of control and HCC.
Results
Association of HCV particle with CD81+ exosomes
The SEM analysis has shown that the HCV virions are presented in three forms in peripheral blood: (i) as a cargo within exosomes, (ii) associated on the surface of exosomes, and (iii) in free isolated state (Fig. 1B).
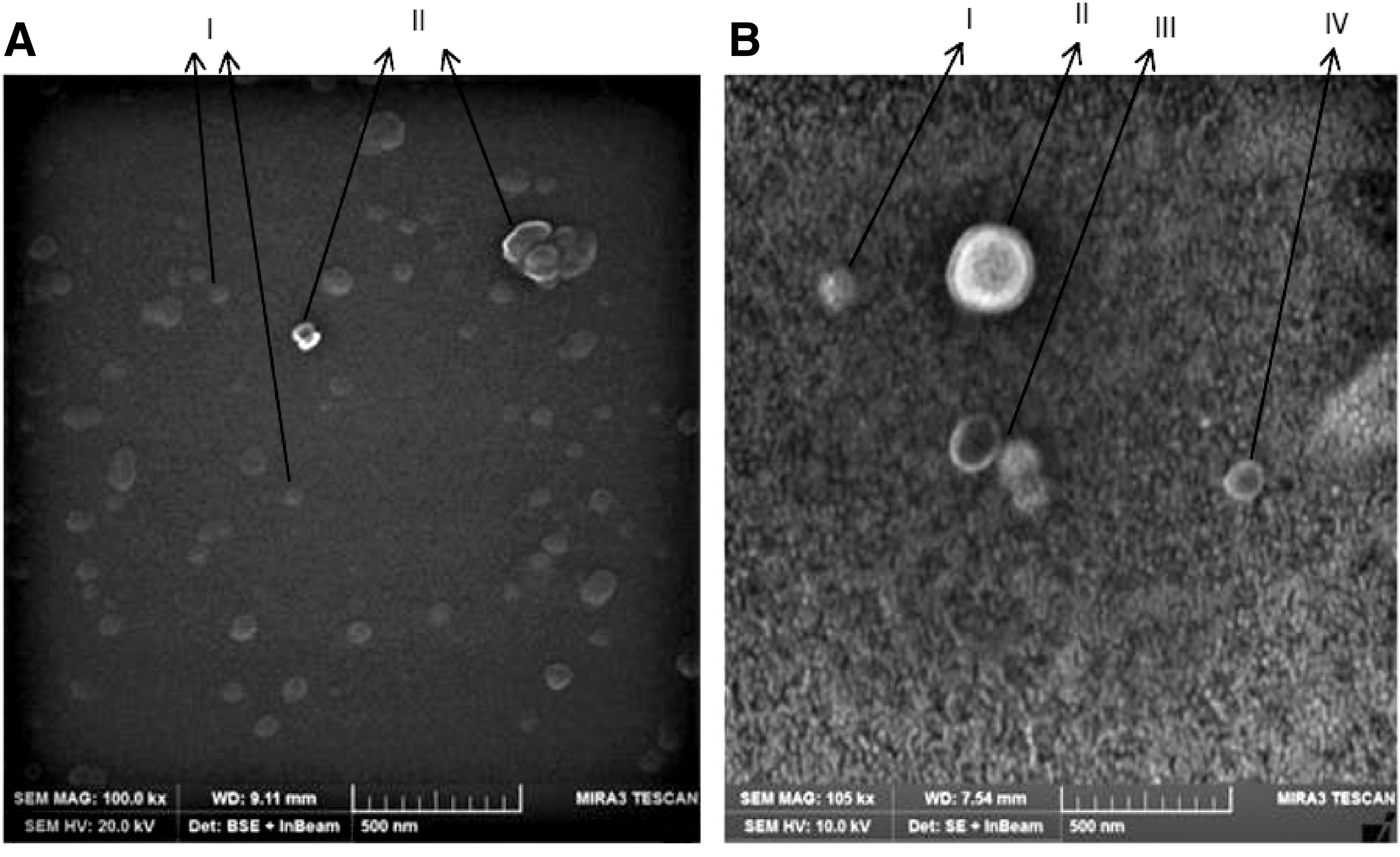
The presence of CD81 on exosomes enriched from healthy controls and HCV-positive HCC was confirmed through 1D-SDS-PAGE. The pattern of protein bands coincided with already published literature on exosomal enrichment. Band of size 23–25 kDa, confirmed the presence of CD9/CD81 in the samples (Fig. 2A, B). These enriched exosomes were then subjected to immunoprecipitation, to purify CD81+ subpopulation of exosomes. The CD81+ exosomes when detected in FACS showed higher size and granularity in case of HCV-positive HCC, when compared with healthy controls (Fig. 3A–C). This evidence proves the association of HCV virion, in specific, to CD81+ exosomes. The count of CD81+ exosomes was found to be variable between healthy controls and HCV-mediated HCC and among the different stages of HCV-mediated HCC. Overall, the average count was higher in HCV-mediated HCC when compared with healthy controls. It was also observed that the counts declined with the progression of HCV-mediated HCC. The nature of interaction of HCV particles with CD81+ exosomes remained uncertain, in terms of the major mechanism.
Real-time quantification of HCV genome in CD81+ exosomal fraction confirmed the presence of HCV genome in association with them (Fig. 4A, B). The HCV load in the CD81+ exosomal fraction was compared with the circulating HCV load in PPP to determine the percentage enrichment in exosomes. We found that the enrichment of HCV in exosomes can vary between as low as 3.2% to as high as 94.6% (Fig. 4C). This enrichment was found to be independent of the viral load observed in plasma and may or may not follow the trend of increased enrichment of HCV in exosomes with increasing viral load (Fig. 4D).
Determination of potential proteins in immune evasion and establishment of persistent infection as therapeutic targets
Since exosome biogenesis is a normal mechanism of intercellular signaling, even in normal physiology, when hijacked by virus in infection, certain exosomal proteins are usually involved in sorting of the virus in the exosome as cargo, including the ESCRT proteins (4). These proteins might be involved in biogenesis of the exosomes or might be enriched in the exosome as cargo. Among the host proteins, either the cellular proteins or the exosomal proteins might be targeted in combination with regular therapy to increase the efficacy and reduce the chances of relapse. Therefore, we found the host proteins that might interact with CD81-positive exosomes involved in immune evasion of HCV, and therefore are potential therapeutic targets for future studies. These proteins include: TSG101, CD19, CLDN 1, CR2, C3, and ITGB1/4 (Fig. 5A). These proteins were found to be interacting with HCV enriched in CD81+ exosomes through prediction and experimentally validated protein/protein interaction in literature (2,4,5,9,11,16,17,21,30). It is also noted that a majority of HCV viral protein, other than E2, can interact directly with CD81, including core p19, NS5A, NS5B, and RdRp (Fig. 5B). This suggests the enrichment of viral proteins in the form of subparticles, as a cargo, in CD81-positive exosomes.
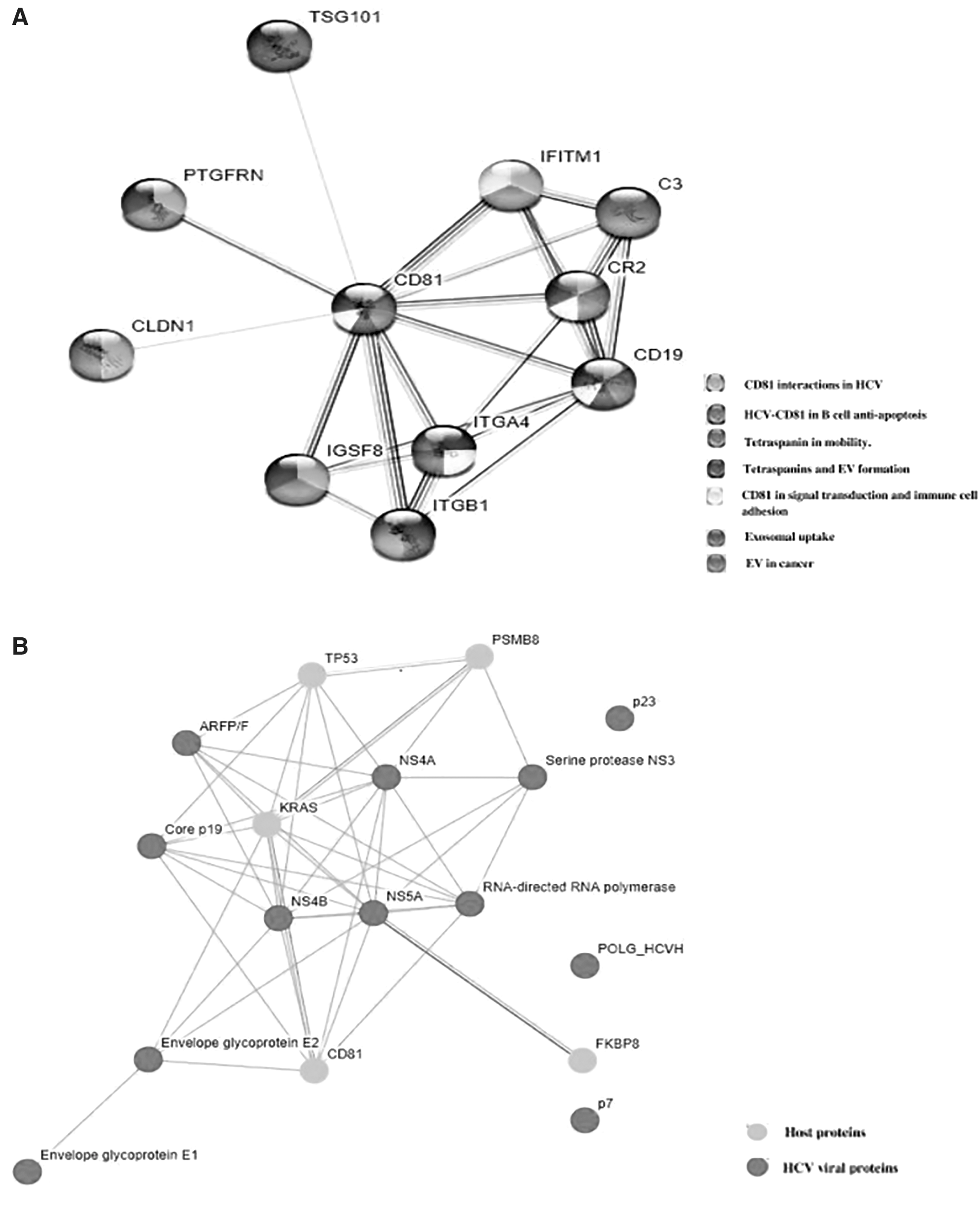
We suggest, that targeting host proteins along with CD81, will restrict the immune evasion of HCV in HCC patients, helping to clear the circulating virus, reducing the extrahepatic infection of HCV, and therefore slowing down the progression of HCV-mediated HCC.
Discussion
It is evident from our and previously published results that HCV particles are found to be associated with CD81+ exosomes. However, its role in immune evasion to develop persistent infection and HCC progression needs to be highlighted. CD81+ exosomes are not only encapsulating the HCV particles and subparticles to avoid interaction with neutralizing antibodies and protecting a fraction of HCV, but are also responsible for immunomodulation of B and T lymphocytes as well as the inhibition of complement system through C3 protein (20). The hijacking of host exosomal pathway for secretion of HCV is found to be an evolutionary beneficiary, for the virus's survival, causing the development of chronic liver disease and insufficient management using the current antiviral program. The currently available antiviral therapies focus on clearing the virus from the liver host cells and not on helping the immune system counter the immune evasion strategies of HCV. If both targets are combined, the clearing of virus from the circulation can be made more efficient.
Our results show that the enrichment of HCV in CD81+ exosomes is higher in early stages of HCC (i.e., in BCLC A and B, the enrichment is around 60% of all circulating HCV), when compared with the later stages (i.e., in BCLC C and D, the enrichment was found to be dropped to 22% and 3.2%, respectively) (Fig. 4B). It is also found to be independent of the circulating viral load in plasma, suggesting that the fraction of HCV secreted through the CD81 exosomes are not dependent of the total HCV being released into the circulation through other pathway (Fig. 4C). It was also observed that in the patient found to be resistant to interferon therapy in the past, the enrichment of HCV in CD81-positive exosomes was as high as 94.6%, suggesting the role of CD81-positive exosomes in immune evasion and resistance against interferon-based therapy (Fig. 4B). However, the limitation of all these deduction made in our study is the sample size tested. We suggest a larger clinical study that correlates the quantity of circulating CD81+ exosomes with the circulating HCV viral load, the stage of liver disease, and the response to therapeutic regime, to confirm the trend. We also suggest that host proteins that interact with CD81 and are responsible for immune evasion of HCV, should be considered as potential therapeutic targets in future studies.
Conclusion
To conclude, the study shows that HCV does hijack the exosomal pathway for secretion of viral particles. The scanning electron micrograph confirms that the associations of these HCV with circulating exosomes in peripheral blood of HCV-mediated HCC patients are found in two forms, as cargo and on surface association. The enrichment of HCV in CD81+ exosomes was confirmed by FACS and by real-time quantitative polymerase chain reaction of immunocaptured CD81+ exosomes from enriched exosome pellet. The HCV enriched in CD81+ exosome fraction was found to be associated with the stage of HCC and interferon therapy response, whereas it was found to be independent of circulating viral load. The literature and STRING analysis confirmed that CD81-E2 complex might be responsible for immune disorders associated with HCV chronic infection. TSG101, CD19, CLDN 1, CR2, C3, and ITGB1/4 were found to be interacting with CD81. These proteins are responsible in packing of viral particles and subparticles in the CD81+ exosomes, as well as inflammation, lymphoproliferative disorders, and decreased production of neutralizing antibodies. This further contributes to persistent infection and progression of liver disease.
Footnotes
Acknowledgments
The authors want to acknowledge the contribution of ASAB administration, ASAB diagnostics laboratory, and Institute of Space and Technology, Islamabad, Pakistan.
Authors' Contributions
S.M. conceived and designed the study, J.I.A.M. and M.A.M. performed the experiments, M.U. provided with clinical expertise and helped with sample collection, M.A.M. wrote the article, and S.M. revised the article.
Ethics Approval
The ethics approval to obtain patient sample was approved by IRF, Rawalpindi Medical University. While the ethics approval to handle the samples at BSL 2 facility was approved by ASAB, NUST.
Author Disclosure Statement
There is no conflict of interest among authors and institutions.
Funding Information
The project was funded by Atta-ur-Rahman School of Applied Biosciences, National University of Sciences and Technology, Islamabad, Pakistan. It was not a part of a grant or other funding source.