
Abstract
Stress-induced subclinical reactivation of varicella-zoster virus (VZV) has been studied previously. However, subclinical reactivation of VZV induced by the stress of pregnancy has not been investigated. The objective was to study varicella DNA and varicella antibody levels in mothers and their newborn babies. VZV immunoglobulin G (IgG) levels in 350 mother–newborn dyads were studied using indirect enzyme-linked immunosorbent assay testing. A subset of 73 dyads was selected, DNA was isolated from the serum samples, and quantitative polymerase chain reaction (qPCR) was performed. Nearly 15% (14.6%) mothers tested were positive for varicella antibodies (>100 mIU/dL) and 16% were borderline (<100 and >50 mIU/dL). Approximately 16.9% of the babies were positive, and 18% were in borderline. Among those tested for VZV-DNA, 70% of mothers with low VZ-IgG (<100 mIU/dL) and 11.32% of those with high VZ-IgG (>100 mIU/dL) were positive for DNA. Among the newborns, 60% of those with low VZ-IgG and 15% of those with high VZ-IgG were positive for DNA. Mothers who have had VZV infection in the past can transmit VZV DNA to their babies.
Introduction
The human virome is a part of the microbiome and it is made up of virus found in or on the human body, in the absence of symptoms of infection (16). Moustafa et al. analyzed the blood DNA virome in 8,000 humans and identified genetic material of 19 viruses among 42% of the asymptomatic individuals they studied. Many members of the Herpesviridae were identified but not the varicella-zoster virus (VZV) (13).
VZV remains latent after primary infection but it is known to reactivate when there is a decline in the cell-mediated immunity. VZV may be reactivated after surgical stress and Mehta et al. have demonstrated that the stress of space travel in astronauts can induce subclinical reactivation of VZV (11). The reactivated VZV may lead to typical symptoms of VZV infection or it could be subclinical and VZV has been detected in saliva of a few patients (6,12).
Reactivation of VZV induced by the stress of pregnancy has not been investigated previously. We look for VZV DNA and VZV antibodies in antenatal maternal blood and in their newborn babies.
Materials and Methods
Ethical approval
Ethical approval was obtained from the St. Stephens Hospital Ethical Committee, Delhi (dated July 1, 2016) and the Institutional Human Ethics Committee of the National Institute of Immunology, New Delhi (IHEC#99/17).
This was a prospective study conducted at a tertiary care hospital in Delhi, India, between June 2016 and June 2017. The study was performed jointly by the St. Stephens Hospital, Delhi and the National Institute of Immunology, New Delhi.
Sample size estimation was based on the results of Linder et al., who observed that 24% of 2-month-old babies carried maternal anti-VZV antibodies (9). The sample size required for detection of a prevalence of 24% with a precision of 5% (α = 0.05) was 281.
Mothers who delivered at St. Stephens Hospital were eligible for inclusion in the study except for those who received blood transfusion during pregnancy or were vaccinated against VZV. Consent of mothers for participation in the study and for collection of cord blood was obtained before delivery. The mother's sample was collected during routine antenatal blood checkups. Cord blood of the neonate was collected at delivery. To ensure we had suitable samples from 300 dyads, consent was obtained from 600 mothers assuming 25% of the samples from mother and babies may not be suitable for testing. We anticipated that, even pessimistically, we could expect 50% of the cases would line up as dyads.
Two milliliters blood from mother and 2 mL cord blood were collected in dry tubes and centrifuged for 15 min at 3,000 g. The serum was extracted and stored at −20°C before being sent to National Institute of Immunology, New Delhi for DNA testing and estimation of antibody titers.
VZV antibodies
The serum antibody to VZV was estimated by indirect enzyme-linked immunosorbent assay (ELISA) method. The serology results are stated as mean titers with standard error of mean.
Preparation of standards
For the standard preparation we quantified the concentration of the VZV positive serum from commercially available kit (varicella-zoster virus IgG, Serion ELISA classic; Würzburg, Germany) along with in-house serum sample. Quantification of standard (from kit) verses in-house serum was performed as described hereunder and the calculation was carried out as suggested in the kit.
“In house” VZV indirect ELISA
Varicella immunoglobulin G (IgG) antibody titer was assessed by an “in-house” indirect ELISA. Coster assay 96-well microtiter plates were coated with varicella vaccine live, 1,350 PFU/0.5 mL (Variped®; Merck Sharp & Dohme B.V., Haarlem, the Netherlands) diluted at 1:100 in 0.01 M phosphate-buffered saline (PBS), pH 7.2, and incubated overnight at 4°C.
Coated plates were washed with 0.01 M PBS Tween-20 (PBST), pH 7.2 once for 2 min. Tenfold dilution of the mother and cord serum samples were prepared in 0.01 M PBS, pH 7.2, and 1% bovine serum albumin; similarly, 1:100 dilution of in-house serum and ready-to-use VZV standard serum (from kit) was added to the plate and incubated for 60 min at 37°C in moist chamber. In the next step, goat anti-human IgG horseradish peroxidase (GeNei Laboratories, Bangalore) diluted 1:200 in 0.01 M PBS, pH 7.2 was added, and incubated for 60 min at 37°C. Furthermore, 100 μL of 3,3′,5,5′-tetramethylbenzidine (BD) substrate was added to each well. As soon as color started to develop, 50 μL of 2 M H2SO4 was added to stop the reaction. OD was read at 450 nm in ELISA reader, using 630 nm as a reference filter (Bio-Tek Instruments). In between the steps, wells were washed three times for 2 min each with 0.01 M PBS, pH 7.2, and 0.05% Tween 20. Varicella-zoster antibodies were expressed in IU/mL using the curve comparison method to convert optical density in concentration units.
According to the product's manual, IgG titers against VZV > 100 mIU/dL is positive, between 50 and 100 mIU/dL is borderline, and <50 mIU/dL is negative.
Varicella-zoster DNA
DNA isolation from patient's serum
DNA was isolated from the serum samples using MasterPure complete DNA and RNA Purification Kit (Epicentre) according to the manufacturer's instructions. In brief, 150 μL of 2 × tissue and cell lysis solution containing 3 μL proteinase K (provided with the kit) was mixed with 150 μL serum sample. The sample was incubated at 65°C for 15 min. The sample was cooled and 1 μL of 5 μg/mL RNase A was added and incubated at 37°C for 30 min. Protein was precipitated by adding MPC protein precipitation reagent to the sample in ice and spun to collect the aqueous supernatant. DNA was precipitated by adding isopropanol followed by washing with 70% ethanol. DNA was quantified by nanodrop spectrophotometer (Thermo Fisher).
Polymerase chain reaction
DNA was isolated using Epicentre kit. DNA was quantified and 150 ng of total DNA was used for the PCR. Primer and TaqMan probe for VZV DNA polymerase gene (Gene bank accession no. X04370) were referred (21) and its specificity was confirmed by primer blast search tool. Melting temperature (Tm) was between 55°C and 60°C and the amplicon length was 63 bp. TaqMan probe sequence (26 nucleotide) was customized with Fluorescein amidites (FAM) at 5′ end and Tetramethylrhodamine (TAMRA) at 3′ end.
qPCR was performed in a total reaction volume of 20 μL using Takyon™ ROX probe MasterMixdTTP Blue from Eurogentec (Seraing, Belgium) according to the recommendations of the manufacturer. Final concentration of 50 nM for the primers and 250 nM for the probes was used. The amplification conditions consisted of one-time Takyon™ activation for 3 min at 95°C followed by 40 cycles of denaturation at 95°C for 10 s and annealing/extension at 58.9°C for 60 s. Because of scarcity of sample volume, randomly selected 30% samples were repeated for DNA extraction followed by PCR amplification was repeated in 30% of the samples to reconfirm the findings.
The quantitative standard template
A synthesized 150 nucleotide sequence (Sigma) as given in Table 1 was used as an internal control. It has 50 and 37 bases upstream of forward primer and downstream of reverse primer, respectively. TaqMan probe designed for VZV detection was checked for cross-reactivity with other herpes viruses using NCBI primer blast software tool. The herpes viruses, human herpes type I, Homo sapiens, and viruses nonredundant databases were used to search any nonspecific target. No such nonspecific template was found indicating the TaqMan probe to be specific. Same probe and primers were used for the target sequence and the internal control.
Sequence of Varicella-Zoster Virus Primers, TaqMan Probe, and 150 Nucleotide Long Sequence
Nucleotide strings of forward primer sequence are given in bold followed by TaqMan probe sequence in bold and italics, and reverse primer sequence in italic for VZV.
VZV, Varicella-zoster virus.
Standard curve
Quantitative estimation of viral DNA was carried out by drawing the standard curve using known amount of 150 nucleotide VZV sequence. Serial dilution was prepared containing 109 to 101 copies per reaction. Enumeration of VZV DNA in mother and child serum was carried out by plotting cycle threshold (Ct) values against the standard curve.
Results
In this study, consent was taken from 600 mothers for participation in the study. Blood samples from 403 mothers and 547 cord blood samples were found acceptable for testing. The mean birth weight of the 547 babies studied was 2.86 kg. There were no preterm babies and very few had low birth weight. A total of 350 maternal blood samples were paired with respective cord blood sample of their babies as summarized in Venn diagram in Figure 1. For VZV DNA estimation, from 350 paired samples—73 mothers and their babies were selected across three VZV antibody categories, high (>100 mIU/dL), borderline (50–100 mIU/dL), and low levels (<50 mIU/dL). The median age of mothers in our study was 27 years and the median weight of babies was 2.9 kg. Among 547 babies, the sex ratio was 1,030 females per 1,000 males. A total of 403 mothers (91.6%) were pregnant for the first time.
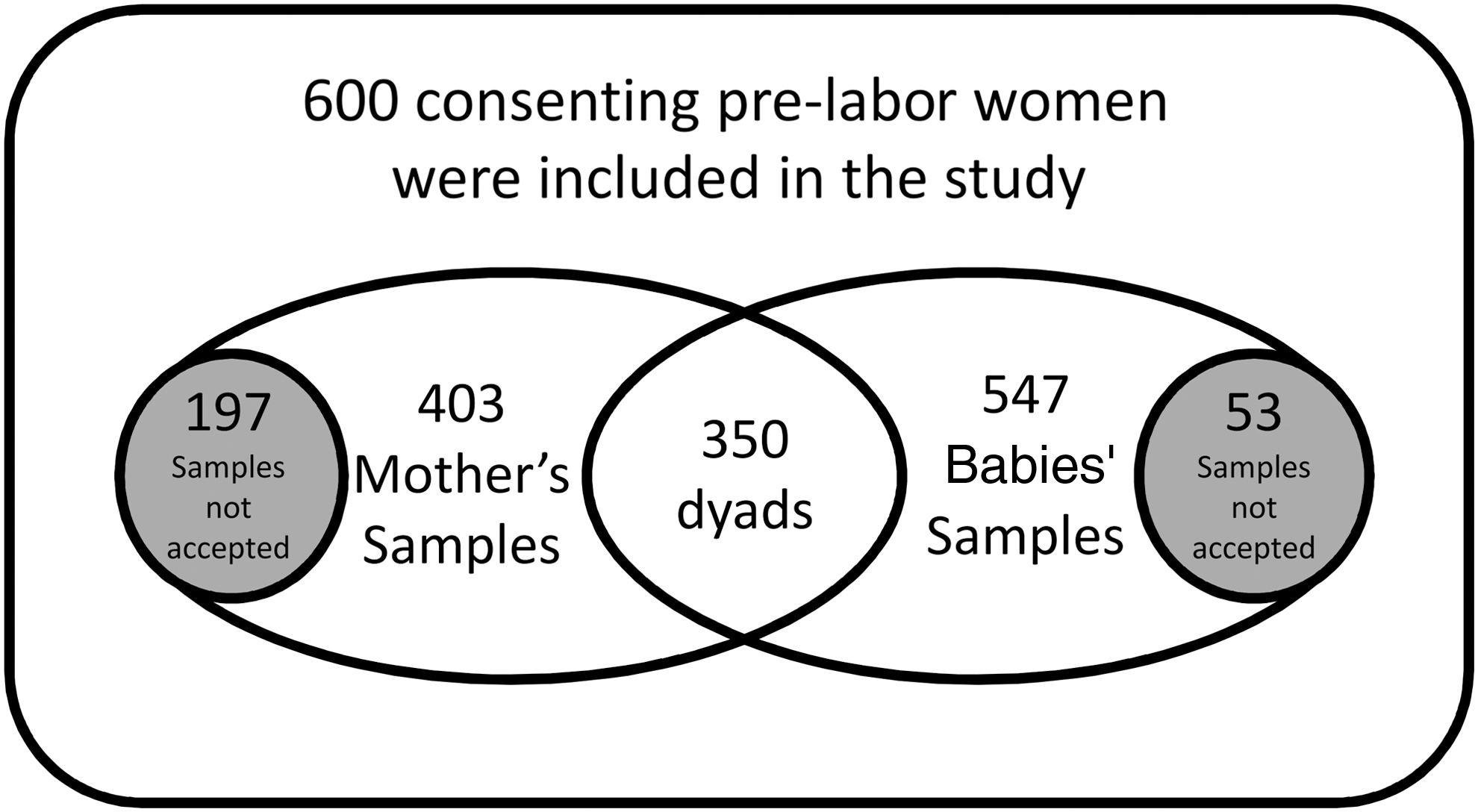
Summary of samples investigated in the study.
VZV antibodies titers
The mean IgG titer for anti-VZV in 403 mothers was 61.87 ± 166.4 mIU/dL, whereas in 547 babies the mean value of IgG titers for anti-VZV was 59.9 ± 127.6 mIU/dL. Among 350 mother–baby dyads, the mean VZV IgG antibody titers in mothers was 67.3 ± 177.61 mIU/dL and babies was 65.02 ± 144.4 mIU/dL.
Thirteen percent of the 403 mothers, and 15% of 547 babies had antibody titers >100 mIU/dL IgG. A total of 177 (50.57%) babies had higher IgG VZV antibodies titers than their mother and 41 babies (11.7%) had titers >50 mIU/dL than their mother's titers. In 350 mother–baby dyads, the IgG titers were correlated, with a correlation coefficient of 0.672 and p < 0.0001.
Among the 350 mother–baby dyads, 15% (14.6%) mothers tested were positive for varicella antibodies (>100 mIU/dL) and 16% were borderline positive (<100 mIU/dL and >50 mIU/dL). About 16.9% of the babies were positive, and 18% were borderline positive. IgG titers in the dyads were correlated, with a correlation coefficient of 0.672 and p < 0.0001.
We studied the maternal age and newborn weight against the neonatal VZV antibodies titers utilizing the receiver operating curve characteristics. The area under the curve was 0.53 and 0.52, respectively. Newborn antibody titers were unrelated to these factors.
Varicella-zoster virus DNA
We used TaqMan probe-mediated qPCR analysis to detect viral DNA transcytosed across placenta. DNA extraction was performed in samples obtained from 73 mothers and their babies. Of the 73 mothers tested for VZV DNA, 53 had VZV IgG antibody levels >100 mIU/dL and 20 had <100 mIU/dL. Six of the 53 (11.32%) with VZV antibody >100 IU/dL tested positive for DNA, whereas 14 of 20 (70%) with VZV antibody <100 IU/dL tested positive for DNA.
In the same way 8 of 53 (15%) newborns with VZV antibody >100 IU/dL tested positive for DNA, whereas 12 of 20 (60%) with VZV antibody <100 IU/dL tested positive for DNA.
When VZ-IgG was high, the viral load was less in maternal system (23–1,156 copies/μL) and fetal system (76–437 copies/μL). However, when neutralizing IgG was low (IgG <100 mIU/dL) and the viral load was higher in maternal system (113–1,406 × 102 copies/μL), thus more VZV DNA was transcytosed to the fetus (18–1,075 copies/μL). These findings are summarized in Table 2.
Relationship Between VZ-IgG Levels and DNA in Mothers and Babies
Discussion
Vaccination against chickenpox is not common in India. Chickenpox is mild disease in children and it usually results in protection against VZV infection for life (2). This disease is highly contagious with attack rate of >85% after exposure (17). Most people are immune by the age of 15 years because of natural infection. Wharton found that only 6% of adolescents 11–19 years were susceptible to VZV infection (22).
Primary varicella infection acquired in adulthood is associated with increased morbidity (7) and mortality (15) compared with infection acquired <14 years of age (5). The reasons for this difference in morbidity with age are not clearly understood. Baba and colleagues have found that the disease is milder in babies born to mothers who have a history of varicella infection. They have suggested this may be because of the partial protection afforded by antibodies to VZV, passively transferred to the newborn (1). However studies on the decay of passively acquired antibodies suggest that such protection is unlikely to last beyond infancy (14,20).
We did the study to observe potential vertical transfer of VZV viral DNA and to examine antibody titer in mothers and their babies at birth. We examined VZV DNA and VZV antibody levels only in serum as some variability has been reported regarding VZV DNA in saliva (6,12).
Ours is arguably the largest such study in India looking at varicella DNA and antibody in mother and baby dyads. Thirteen percent of tested mothers were positive for VZV antibodies, whereas 15% babies were positive. The maternal antibody levels were correlated to the neonatal antibody titers with a correlation coefficient of 0.672 (p < 0.0001). Our study supports the work of Leuridan et al. who demonstrated that the antibodies for infection like varicella and rubella in neonates at birth correlates strongly with the maternal values (8). There was no relation between maternal age and VZV antibody titers. Doroudchi et al. have also shown that maternal age and parity do not influence transplacental antibody transfer (3).
Although there was a correlation between maternal and fetal antibody levels, 21.7% babies had higher titers than their mothers (>25 mIU/dL), and 11.7% had titers >50 mIU/dL than their mothers. It suggests that there may be active transplancental transport of IgG from mother to the baby. This is supported by the work of Techasena et al. and Wutzler et al. (18,24). They found that placenta actively transports IgG to the baby. van den Berg et al. found that preterm babies had lower antibody titers with respect to term babies (19).
It is known that hFcRn mediates transcytosis of IgG across synctiotrophoblast. This mechanism plays a central role in protecting the fetus by passive immunization. In fetal disease for congenital VZV, the treatment of pregnant women with hyperimmune antiviral antibodies is known to suppress infection in the mother and limit fetal disease for congenital VZV (4), suggesting that hFcRn mediates transcytosis of VZV IgG across synctiotrophoblast.
Upon analyzing 73 dyads for VZV viral DNA using TaqMan probe qPCR study, 19% were positive for viral antigen. This is similar to the finding of Mehta et al. who found subclinical reactivation of VZV with stress (11). We found more copies of viral DNA in mothers with lower IgG titers (IgG <100 mIU/dL) and there was more likelihood of transmission of DNA to their babies. This indicates that the protective ability of the antibody is concentration dependent. Higher (>100 mIU/dL) neutralizing antibody renders higher viral clearance in mother and less transfer to the fetus as seen by lower viral load in such dyads. Given the active transplacental transfer machinery, more often higher viral load was seen in fetal system than maternal system. Our finding suggests a vertical transfer of VZV DNA. Moreover, qPCR analysis also shows the presence of viral DNA in newborns. This could be since the viral DNA can transcytose across the placental barrier and mediate a vertical transfer from mother to offspring.
Studies suggest hFcRn can cause transfer of viral DNA by the receptor-mediated endocytosis (10,23). One possibility is that the VZV DNA vertical transfer can potentially be mediated by this mechanism. Our findings suggest that when maternal and fetal IgG level is higher (>100 mIU/dL) the viral titer was less in mothers (23–1,156 copies/μL) and lower in fetal system (76–437 copies/μL). However, when neutralizing IgG antibodies were low (IgG <100 mIU/dL) the viral load was higher in maternal system (113–1,406 × 102 copies/μL), and more VZV DNA was transcytosed to the fetus (18–1,075 copies/μL). In such cases, the fetus receives lower IgG but higher VZV viral DNA.
It is likely that in such cases antibodies were developed actively in the fetus when viral DNA is high. It should be emphasized that the samples included in the study were asymptomatic of VZV infection and that we have not examined the infectivity and replication of the viral DNA in host system. Thus, the exact nature of antibody–viral DNA interaction in circulation needs to be elucidated. However, actively acquired antibodies are likely to last longer. It may explain why varicella infection is a mild infection at <14 years of age (5), well after the stage where passively acquired antibodies from the mother would have waned. More studies are needed to confirm our findings.
Footnotes
Authors' Contributions
J.P., V.R., P.U. conceived the project; V.R., S.N.L.; A.M., A.K., K.V.M. conducted the study, and writing the first draft; A.M., A.K., K.V.M., and P.U. did the testing. The final draft was by K.V.M., A.K., J.P., and P.U. All the authors have seen the final article and approved it. J.P. and P.U. stand as guarantors for the work and corresponding authors.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by the core grant received from the Department of Biotechnology, Government of India to National Institute of Immunology, New Delhi. Funders have no role in study design, collection, analysis, and interpretation of data.