
Abstract
In this study, the immunoadjuvant effects of recombinant porcine interferon alpha (rPoIFNα) on the killed virus vaccine (KV) of porcine reproductive and respiratory syndrome virus (PRRSV) in pigs were investigated. The experimental pigs were divided into six groups, including normal control group, rPoIFNα control group, PRRSV KV control group, KV+40,000 U rPoIFNα immunization group, KV+400,000 U rPoIFNα immunization group, and KV+4,000,000 U rPoIFNα immunization group. The experimental pigs were boosted immunized on the 28th day after the initial immunization, and the heparinized blood and serum samples were collected at different time points of these two immunizations to detect and evaluate the immune responses of pigs after immunization by ELISA assay, neutralization assay, flow cytometry, and so on. The results showed that the proportion of the levels of PRRSV-specific antibodies, neutralizing antibodies, stimulation index, IL-4, IFN-γ, and lymphocytes within the groups immunized with KV+rPoIFNα were significantly higher than that group immunized with KV alone. The humoral and cellular immune responses in pigs were markedly enhanced by rPoIFNα after the coadministration with KV vaccine. Therefore, we tentatively think that rPoIFNα is a potential immune promoter with prospects for future applications in the pig industry.
Introduction
Porcine reproductive and respiratory syndrome (PRRS) is a vicious infectious disease in pigs. It is a result of the infection of porcine reproductive and respiratory syndrome virus (PRRSV). Besides, it has a high incidence, can rapidly spread, and seriously jeopardizes the development of the global and Chinese pig industry. The immunosuppression and persistent infection caused by PRRSV infection in the body has always been a difficult problem in PRRS prevention and control (37).
At present, the main preventive measure of PRRS is still vaccination, but neither PRRSV-inactivated killed virus vaccine (KV) nor modified live virus vaccine (MLV) can sufficiently stimulate the immune system to produce a strong immune response for the virus clearance, only inducing little neutralizing antibodies (NAs) titers to limit the continued infection of PRRSV. Nowadays, the efficacy of PRRSV KV vaccine is not ideal (2). To improve the efficacy of the PRRSV KV vaccine, studies have been explored to increase and amplify its immunogenicity using a number of adjuvants, such as interleukin-2 (IL-2), interferon-gamma (IFN-γ), and CpG oligodeoxyribonucleotides (31,39).
IFNα belongs to type I IFN and is a major effector molecule for adjuvants such as CpG and Freund's complete adjuvant (4). It can significantly promote the protective immune response of the influenza vaccine and the foot-and-mouth disease subunit vaccine, helping to produce high levels of IgG2a antibodies (7,30).
Studies had demonstrated that IFNα could be used as adjuvants when co-administrated with antigens, such as some soluble proteins (17), inactivated virus vaccine (30), or genetically modified DNA vaccine (6). Administration of IFNα with an influenza virus subunit vaccine can protect mice from viral challenge and induces a Th1-type immune response; however, a solo subunit vaccine cannot (30). Similarly, in a research study reported by Cull et al., a DNA expressing IFNα was co-injected with DNA encoding the cytomegalovirus glycoprotein B gene (gB) to immunize mice against cytomegalovirus-induced myocarditis. The coadministration of both DNA has better immunoprotective effects than immunization of gB DNA alone (6).
It was thought that the administration of IFNα in PRRSV-vaccinated pigs might help increase IFN-γ response to PRRSV vaccine. In pigs, IFNα promotes IL-1 production of monocytes and cytotoxicity of lymphocytes (3). With regard to an adjuvant effect in vaccination experiments, type I IFNs have been shown to have potent activity when coadministered with inactivated vaccine preparations by the parenteral route (12,35). Humoral IgG and IgA levels were significantly elevated in a dose-dependent manner when IFN-α was coadministered with an inactivated influenza vaccine.
Porcine interferon alpha (PoIFNα) is a cytokine with a variety of biological activities. It has long been known for its powerful antiviral function, but little was understood for its immunoadjuvant activity. Previous studies reported that IFNα had a strong stimulating effect on porcine immune cells (4). And IFNα has been utilized in the form of expression plasmids (24,33) or expression plasmids plus recombinant porcine interferon alpha (rPoIFNα) (5) for PRRSV MLV. However, the immune effect of rPoIFNα on KV has not been reported.
To further evaluate the feasibility of using rPoIFNα in pigs, in this study, we mixed a soluble rPoIFNα (40) prepared in our laboratory with the PRRSV KV vaccine to coadminister the pigs to develop a new PRRSV-inactivated vaccine containing cytokine adjuvants. Our results indicate that compared with PRRSV-inactivated vaccine (KV) alone, the coadministration of KV vaccine and rPoIFNα can significantly promote the proliferation of peripheral blood mononuclear cells (PBMCs), produce Cytokines and antibodies, and enhance the immune response of pigs. This study focused on the understanding of the prospect of rPoIFNα as an adjuvant candidate for the PRRSV KV vaccine, and hopes to provide a new option for preventing pigs from PRRSV infection and for PRRS controlling.
Materials and Methods
Drugs and reagents
Concanavalin A (ConA), 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) were purchased from Sigma-Aldrich Corporation. The medium (RPMI-1640) for PBMC culture and fetal bovine serum (FBS) were purchased from Gibco Corporation, and the PRRSV-specific antibody ELISA kits were provided by Anhui Jiuchuan Biotech Co., Ltd. (Wuhu, Anhui, China) (Chinese patent number ZL201810752119.2) and IDEXX Co., Ltd. (IDEXX Laboratories, Inc., Westbrook, ME); the PRRS KV vaccine (oil emulsion type) was purchased from the Unibio Co., Ltd. (Yangzhou, Jiangsu, China). MARC-145 cells, a derivative of African green monkey kidney cells from MA-104 cells, were obtained from the China Center for Type Culture Collection (Wuhan, Hubei, China).
Ninety six-well cell culture plates were bought from Corning, Inc. (NY); Ficoll-Paque was purchased from Amersham Biosciences (Little Chalfont, UK); Vacutest tubes containing lithium heparin were purchased from HuNai Biotech Co., Ltd. (Hunan, China); the MiniBEST viral RNA/DNA extraction kit and the PrimeScript® one-step RT-PCR kit was bought from TaKaRa (China) group (Dalian, Liaoning, China); the IFN-γ/IL-4 ELISA kit was purchased from Biorbyt, Inc. (Cambridge, UK); and the T lymphocyte subset detection reagents were purchased from Lifespan, Inc. (Milpitas, CA). rPoIFNα protein was produced by Anhui Jiuchuan Biotech Co., Ltd. as previously described and stored in an ultralow-temperature refrigerator at −80°C (40).
Experimental animals and immunization grouping
In this study, twenty-four 30-day-old Landrace pigs were purchased from commercial herds that did not contain PRRSV and were raised at the Experimental Center of Anhui Medical University. They were fed independently during the whole experiment. To ensure that the experimental animals are PRRSV-negative, serum samples of pigs were collected before the experiment; PRRSV antibody ELISA kit (Anhui Jiuchuan Biotech Co., Ltd.) and IDEXX ELISA kit (IDEXX Laboratories, Inc.) were used to examine the presence of PRRSV-specific antibodies. PCR test was used to examine the viremia of PRRSV, porcine parvovirus (PPV), pseudorabies virus (PRV), classical swine fever virus (CSFV), and porcine circovirus type 2 (PCV-2) (14,19). All PRRSV-specific antibodies and viremia of PRRSV, PPV, PRV, CSFV, and PCV-2 were negative in all experimental pigs.
The pigs were then randomly divided into six groups of four in each group. Group A was a normal control, and pigs were immunized with phosphate-buffered saline (PBS); Group B was an IFN control, and pigs were immunized with 4,000,000 U rPoIFNα; Group C was a vaccine control, and pigs were immunized with PRRSV KV vaccine alone; Group D was a test group, and pigs were coadministered with 40,000 U rPoIFNα and PRRSV KV vaccine; Group E was a test group, and pigs were coadministered with 400,000 U rPoIFNα and PRRSV KV vaccine, Group F was also a test group, and pigs were coadministered with 4,000,000 U rPoIFNα and PRRSV KV vaccine.
The total volume of all preparations used for administration is 2 mL. After measuring the body temperature and the body weight of the pigs at 8 am on the test day, they were injected into the neck muscles of pigs through intramuscular administration according to the animal immunization protocol. The immunization was boosted on the 28th day after the initial immunization. All animal experimental procedures performed in this study are conformed to the policies of the National Institutes of Health Guide for the Care and Use of Laboratory Animals and were approved by the Bioethics Committee of Anhui Medical University.
Clinical symptoms monitoring
(i) The clinical symptoms such as mental state, appetite, respiratory state, and skin rash are recorded daily after immunization. (ii) Body temperature measurement: The acute toxicity was examined after immunization. The temperature of pigs was measured at 0 h (8 am on the test day), 4 h, and 10 h postimmunization, respectively. Then the temperature of pigs was measured at 8 am every day, and was continuously measured until the end of the experiment. (iii) Body weight measurement: The weight of pigs was measured at the same time point before the experiment, 6 days interval during the experimental period, and after the experiment. Relative daily weight gain (daily gain weight/original weight) was calculated. Clinical respiratory scores, whose range was from 0 to 3, have been also recorded as previously described (21).
Blood sample collections
Routine blood counting was determined to confirm the normal hematologic status of the experimental pigs. On the 0th, 7th, 14th, 21st, 28th, 35th, 42nd, 49th, and 56th day after initial immunization, blood samples were collected by jugular vein venipuncture using the vacuum-pumped lithium heparin (150USP unit) test tubes for immune responses measurement using PBMC proliferation assay, IL-4 and IFN-γ assay, and flow cytometry. At the same time, serum samples from nonanticoagulant specimens were collected and stored under −20°C before the measurement of PRRSV-specific antibody and detection of neutralization antibodies.
Isolations of PBMCs
As Linghua et al. reported, porcine PBMCs were isolated from the heparinized blood samples using the method of density gradient centrifugation (20). In short, first, a certain volume of peripheral blood was diluted with the same amount of PBS and then was carefully laid on Ficoll-Hypaque PLUS. After centrifugation at room temperature for 35 min at 1,000 g, the PBMC-containing layers were taken out and washed once with PBS solution. The hypoosmotic shocked erythrocytes were removed, and washing the remaining cells two times with 1640 complete medium (containing 2 mM
Proliferation assay of PRRSV-specific PBMCs
PRRSV purified antigen was prepared using MARC-145 cells infected with the North American prototype PRRSV strain VR2332, as previously described (10). Mock control antigens were prepared in the same manner using uninfected MARC-145 cells. Cultured PBMCs were incited with purified PRRSV antigens with a final concentration of 10 μg/mL. The PBMCs were incited with PRRSV mock control antigens (10 μg/mL) were set as mock control, and PBMCs incited with ConA (10 μg/mL) were set as positive control. After incubation for 72 h at 37°C, 5% CO2, the standard MTT [3-(4,5-dimethylthiazol-2-yl) 2,5-diphenyltetrazolium bromide] method was used to measure the proliferation responses of PBMCs.
The degree of lymphocyte proliferation was expressed in the form of stimulation index (SI), which was defined as the average of the experimental data divided by the average of the nonstimulated controls (8,25). It represented the ratio of the OD570nm in the stimulated well to the OD570nm in the nonstimulated well. The results are the geometric mean values of the SI plus standard deviations (SDs) for triplicate samples.
Determination of titer of PRRSV-specific antibodies
PRRSV-specific antibody titers in pig sera were analyzed using the PRRSV antibody ELISA kit (Chinese patent number ZL201810752119.2) (Anhui Jiuchuan Biotech Co., Ltd.) for primary screening and IDEXX ELISA kit (IDEXX Laboratories, Inc.) for subsequent confirmation. PRRSV-specific antibody titer is reported in the form of S/P ratio, and serum samples are considered as positive when the S/P ratio was 0.4 or higher. However, when the S/P ratio was <0.4, the serum samples are considered as negative.
Serum neutralization assay for PRRSV NA detections
Serum neutralization (SN) assays were conducted as described in the previous literature (13). In short, all serum samples collected from pigs were thermally inactivated (56°C, 30 min) and serially diluted at the ratio of 1:2. Then, the diluted sera were mixed with the same volume of 100 TCID50 pathogenic PRRSV suspensions. After the incubation of the mixture at 37°C for 1 h, they were transferred to the monolayer of MARC-145 cells in a 96-well cell culture plate. Next, the cytopathic effect (CPE) appeared in each well was examined after continuous incubation of the plate at 37°C for 5 days. At the same time, positive and negative controls were established, using PRRSV positive and negative sera from pigs. The titers of NA of each serum sample were computed using the method of Reed–Muench formula (32), expressed in the form of ND50 (the highest dilution for neutralizing 50% of viral activity).
Cytokine assay
Th cell subsets can be differentiated by its secreted expression pattern of cytokines. Peripheral blood lymphocyte proliferation assay and IL-4 and IFN-γ secretion after stimulation can reflect changes in cell-mediated immunity (CMI) status. The antigen-stimulated lymphocyte culture supernatants were collected at 72 h after stimulation. Then, the commercially available porcine IFN-γ/IL-4 ELISA kits (Biorbyt, Inc., Cambridge, UK) were used to determine the content of IFN-γ (Th1 type cytokine) and IL-4 (Th2 type cytokine) according to the producer's recommendations. The geometric mean and SD of triplicate samples were calculated (9,18,38).
Flow cytometry analysis of T lymphocytes in peripheral blood
As described previously (11), the phenotypes and frequencies of the lymphocyte subsets cell populations from 50,000 immunostained PBMC events were identified by flow cytometry. For intracellular IFN-γ staining, as described earlier, Monensin (GolgiPlug™; BD Biosciences, Franklin Lakes, NJ) was added during the last 6 h of incubation with PBMCs that were not stimulated or stimulated with PRRSV antigens. PBMCs were first surface labeled with porcine lymphocyte-specific mAbs (CD3, CD4, and CD8α) and then labeled with secondary fluorochrome-conjugated antibodies (anti-CD3-FITC, anti-CD4-PE, and anti-CD8-SPRD).
Surface-immunostained cells were fixed with 1% paraformaldehyde and permeabilized with the cell-permeabilization buffer (85.9% deionized water, 11% Ca2+ free or Mg2+ free PBS, 3% formaldehyde solution and 0.1% saponin) at 4°C for 18 h. Cells were washed and fluorescently labeled antiporcine IFN-γ (clone P2G10) or its isotype control monoclonal antibody (BD Biosciences) in 0.1% saponin containing fluorescence-activated cell sorting (FACS) buffer. Immunostained cells were analyzed using FACS Aria II (BD Biosciences) flow cytometry.
The gating strategy is to first gate the cells for CD3 marker and then gate CD3 + cells for CD4 and CD8 α marker. Next, the corresponding lymphocyte subsets (CD3 +CD4 −CD8 +, CD3 +CD4 +CD8 −, and CD3 +CD4 +CD8 +) was further gated for IFN-γ labeling, and the frequency of lymphocyte subsets in each group was obtained (CD3 +CD4 −CD8 +IFN-γ +, CD3 +CD4 +CD8 −IFN-γ +, and CD3 +CD4 + CD8 +IFN-γ +). All specific cell population frequencies are expressed as a percentage of total CD3 + lymphocytes.
Statistical analysis
Statistical analysis was performed using the SPSS 18.0 software (SPSS, Inc., Chicago, IL). The experimental data were expressed in the form of average ± SD. The average value comparison was performed using the t-test analysis, and p < 0.05 was set to be statistically different.
Results
Clinical signs of each group of experimental pigs
Pigs were weighed at different time points after initial immunization and booster immunization. Weight gain is expressed in kilograms. The results showed that the weight of pigs in each group increased, but the difference in weight gain among pigs in each group is not significant (Fig. 1). Besides, the rectal temperature of each group of pigs also changed, but all of them were <41°C, and there was no fever, only a small fluctuation was observed (Fig. 2). In addition, there were no major changes in the clinical manifestations, mental status, and behavior of the pigs (data not shown). These results indicate that rPoIFNα prepared by Escherichia coli has weak cytotoxicity and good safety.
Weight gain of pigs after immunization in different groups. Pigs were weighed at different time points after the initial immunization and the booster immunization. The weight gain is expressed in kilograms in the form of average ± SD of the pigs at the time of measurement. SD, standard deviation.

Average rectal temperature of pigs in each group after immunization. The rectal temperatures ≥41°C are considered as fever temperature. The rectal temperature is expressed in degree centigrade in the form of the average ± SD of the test pigs at the time of measurement.
Specific antibody titers against PRRSV
The responses of PRRSV-specific antibodies in six experimental groups were measured by ELISA, and the developmental kinetic results are presented in Figure 3. Sera collected at different time points after the initial immunization was subjected to the evaluation of PRRSV-specific antibody levels. At 7 day postimmunization (dpi), PRRSV-specific antibodies were detected in the KV+4,000,000 U rPoIFNα group; however, none of the antibodies was detected in the other five groups. At 28 dpi, anti-PRRSV-specific antibodies in pigs vaccinated with KV+400,000 U rPoIFNα, KV+4,000,000 U rPoIFNα increased significantly after booster immunization. At 42 and 56 dpi, IgG levels from the groups of KV+400,000 U rPoIFNα and the KV+4,000,000 U rPoIFNα were higher than that from the group of KV alone (p < 0.05).
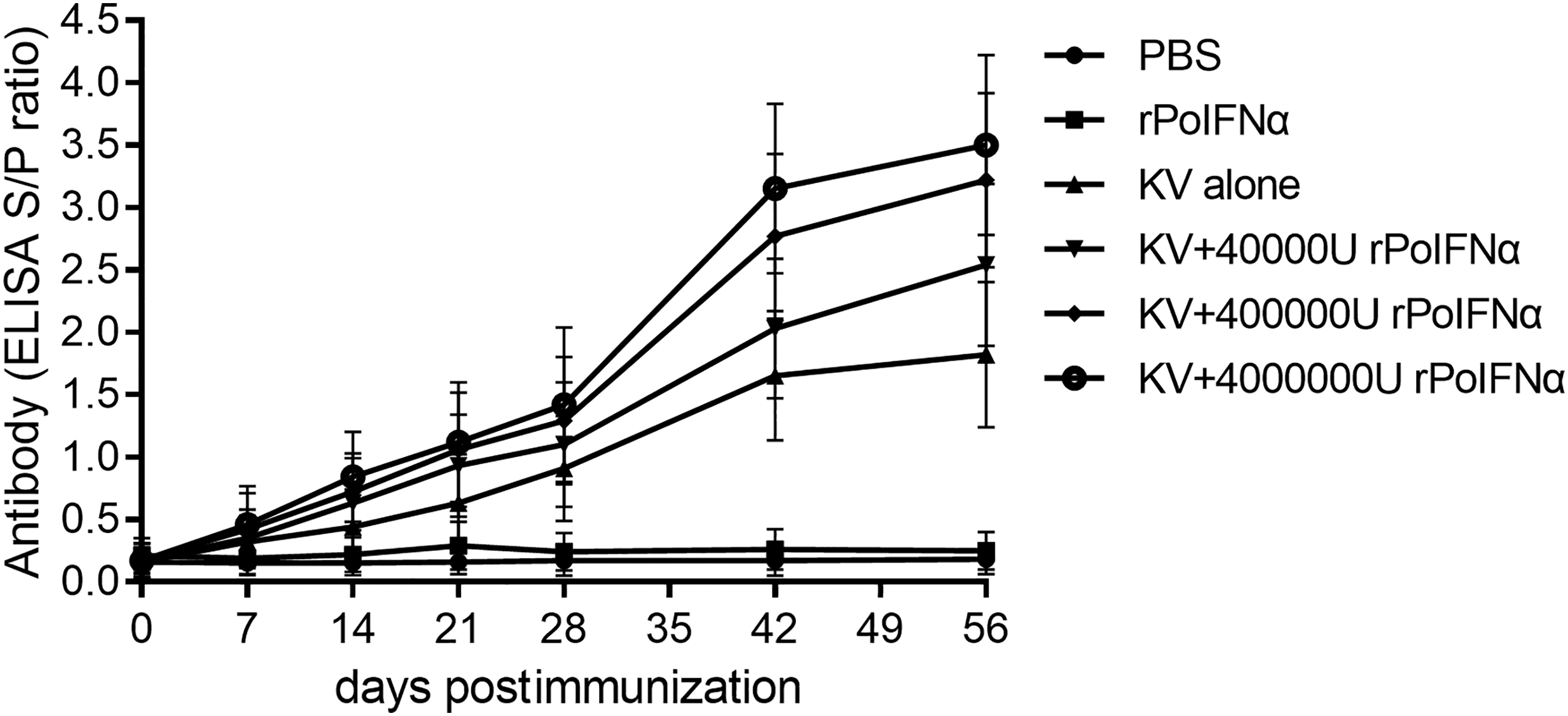
Kinetics of specific antibody responses to PRRSV in different groups of pigs as determined by ELISA. Six groups (n = 4) of pigs were immunized through the intramuscular route (im) at 30 days of age. Each value represents the average S/P ratio ± SD of PRRSV-specific antibodies determined in triplicate. An S/P ratio <0.4 was considered as negative; however, an S/P ratio ≥0.4 was considered as positive. The statistical difference was significant (p < 0.05). PRRSV, porcine reproductive and respiratory syndrome virus.
After immunization, pigs from the groups of KV+400,000 U rPoIFNα and the KV+4,000,000 U rPoIFNα had higher antibody titers than the group of KV alone (p < 0.05). The maximum adjuvant effect was achieved with KV+4,000,000 U rPoIFNα and was observed for at least 8 weeks. However, the difference between the KV alone group and the KV+40,000 U rPoIFNα group was insignificant (p > 0.05). Furthermore, during the period of postimmunization, PRRSV-specific antibodies were not detected in the PBS group or the 400,000 U rPoIFNα alone group. Therefore, enhanced antibody responses through rPoIFNα are likely to occur in a dose-dependent manner.
PRRSV NA titer
The titers of PRRSV NAs in serum samples were further measured using SN assay. The results showed that before the booster immunization (28 days after the initial immunization), no NA was detected in the KV alone group. Also, the pigs of the KV+4,000,000 U rPoIFNα and KV+400,000 U rPoIFNα groups produced antibodies at a titer <1:4. After booster immunization, pigs in the KV alone group produced 1:4–1:8 NAs at 42 dpi and 1:12–1:16 NAs at 56 dpi. However, pigs vaccinated with KV+4,000,000 U rPoIFNα rapidly increased NAs at 42 dpi and reached peak levels at 56 dpi. In fact, pigs immunized with KV+4,000,000 U rPoIFNα produced 1:16–1:32 NAs at 42 and 56 dpi.
Overall, the levels of NAs in the groups of KV+4,000,000 U rPoIFNα and KV+400,000 U rPoIFNα were both significantly higher than those in the group of KV alone at 42 and 56 dpi (Fig. 4). However, the PRRSV NAs were not detected in pigs groups immunized with 400,000 U rPoIFNα alone or PBS. Thus, the KV+4,000,000 U rPoIFNα and KV+400,000 U rPoIFNα groups induced higher antibody levels than the KV alone group, increasing the humoral immune response of PRRSV.
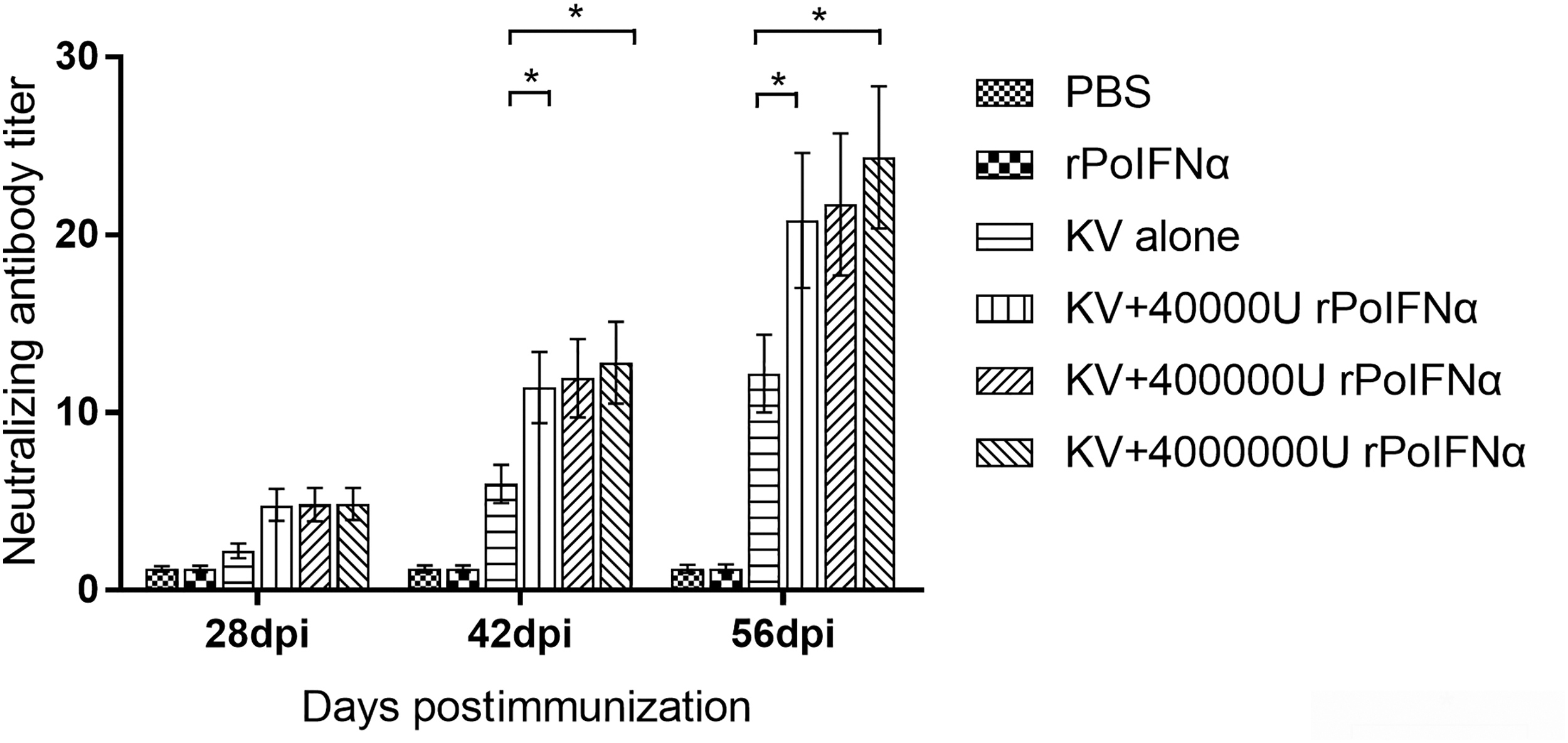
PRRSV-specific NA titers in immunized pigs at different time points. The Y-axis represents the NA levels in each pig group immunized with different reagents such as PBS or KV or rPoIFNα or rPoIFNα+KV. Serum samples from each group of animals at different time points were collected and assayed by serum neutralization assay. The titer of the NA is expressed as the reciprocal of the final serum dilution neutralizing 100 TCID50 PRRSV in 50% of the wells. Each bar represents the average ± SD (n = 4) of triplicate determinations of this group. *The difference between the two groups is significant (p < 0.05). KV, killed virus vaccine; NA, neutralizing antibody; rPoIFNα, recombinant porcine interferon alpha.
Specific cytokines secretion in porcine PBMC culture supernatant
Cytokines play a leading role in regulating the effectiveness of immune responses to infection or vaccination. To monitor the expression of cytokines, supernatants of lymphocytes stimulated with pure PRRSV viral antigen were collected to detect changes of IFN-γ and IL-4 using the commercial ELISA kits. The peripheral blood lymphocytes of pigs were stimulated with specific antigens at different points of time after immunization.
The ELISA results showed that the IFN-γ secretion in the groups of KV alone was lower than that in the group of rPoIFNα alone. However, the secretion of IFN-γ in KV+rPoIFNα groups was significantly higher than that in KV alone group (p < 0.05). The differences in IFN-γ and IL-4 levels produced between the groups of PBS and KV alone were insignificant (Figs. 5 and 6). These data suggested that CMI in experimental pigs increased in response to rPoIFNα coadministration. However, this increase did not happen in the absence of rPoIFNα.
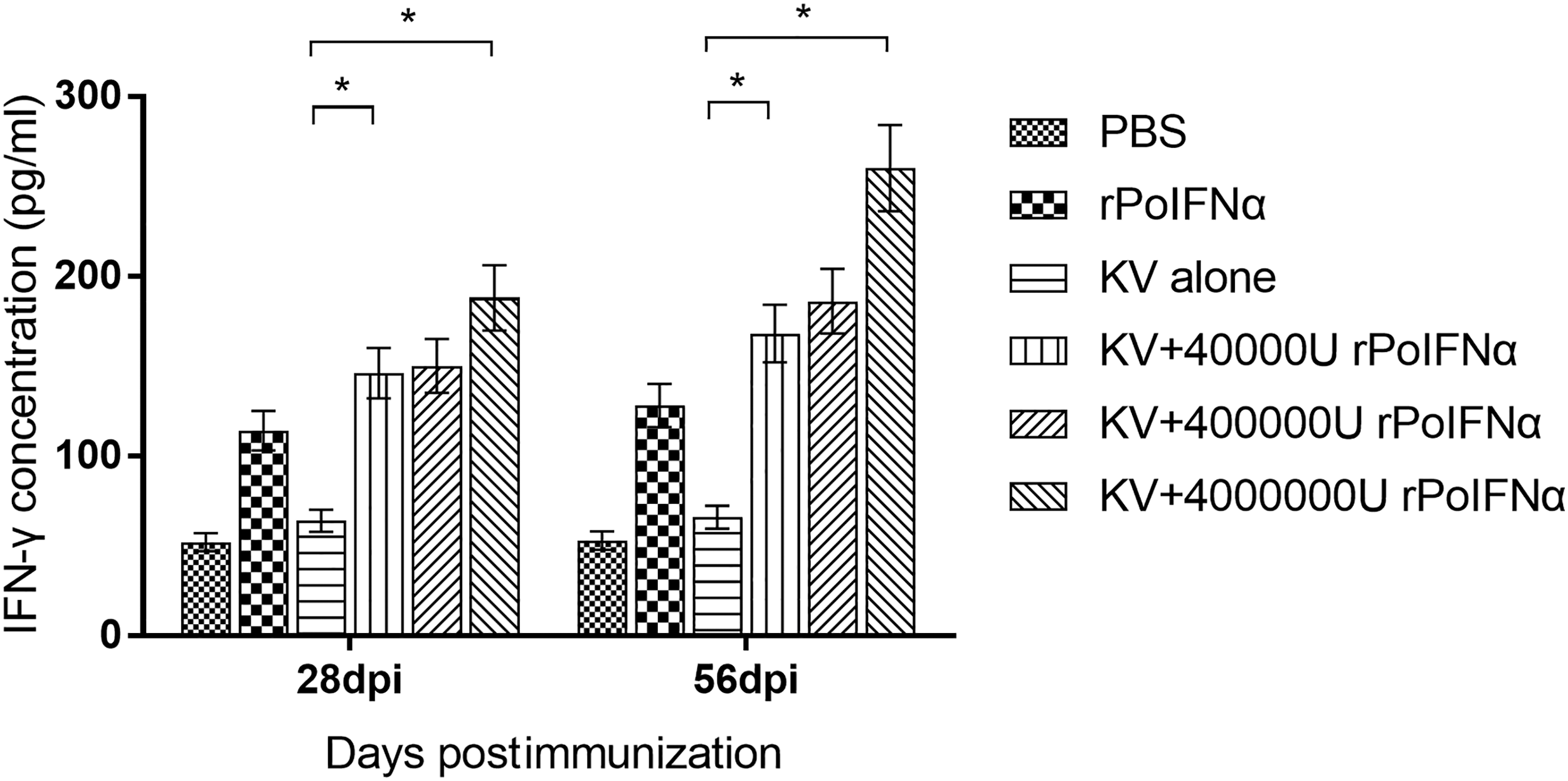
Production levels of IFN-γ at 28 and 56 dpi in porcine PBMC supernatants as measured by ELISA assay. Each bar represents the average ± SD (n = 4) of triplicate determinations of this group. *The difference between the two groups is significant (p < 0.05). dpi, day postimmunization; IFN-γ, interferon-gamma; PBMC, peripheral blood mononuclear cell.

Production of IL-4 in 28 and 56 dpi porcine PBMC supernatants as measured by ELISA assay. Each bar represents the average ± SD (n = 4) of triplicate determinations of this group. *The difference between the two groups is significant (p < 0.05).
For KV+rPoIFNα group, with the increase of rPoIFNα dosage, the concentration of IFN-γ and IL-4 cytokines in pigs increased, indicating that rPoIFNα cytokine could enhance the immune effect of porcine cells (Figs. 5 and 6). Therefore, coadministration of rPoIFNα with KV vaccine enhanced Th1 and Th2 cytokine responses, and the increase in CMI in pigs was a response to rPoIFNα.
Induction of PRRSV-specific PBMC proliferation
The lymphocyte proliferation mediated by rPoIFNα was expressed as the SI through the computation using the MTT colorimetric assay. The results of PBMC proliferation studies in each group are shown in Figure 7.
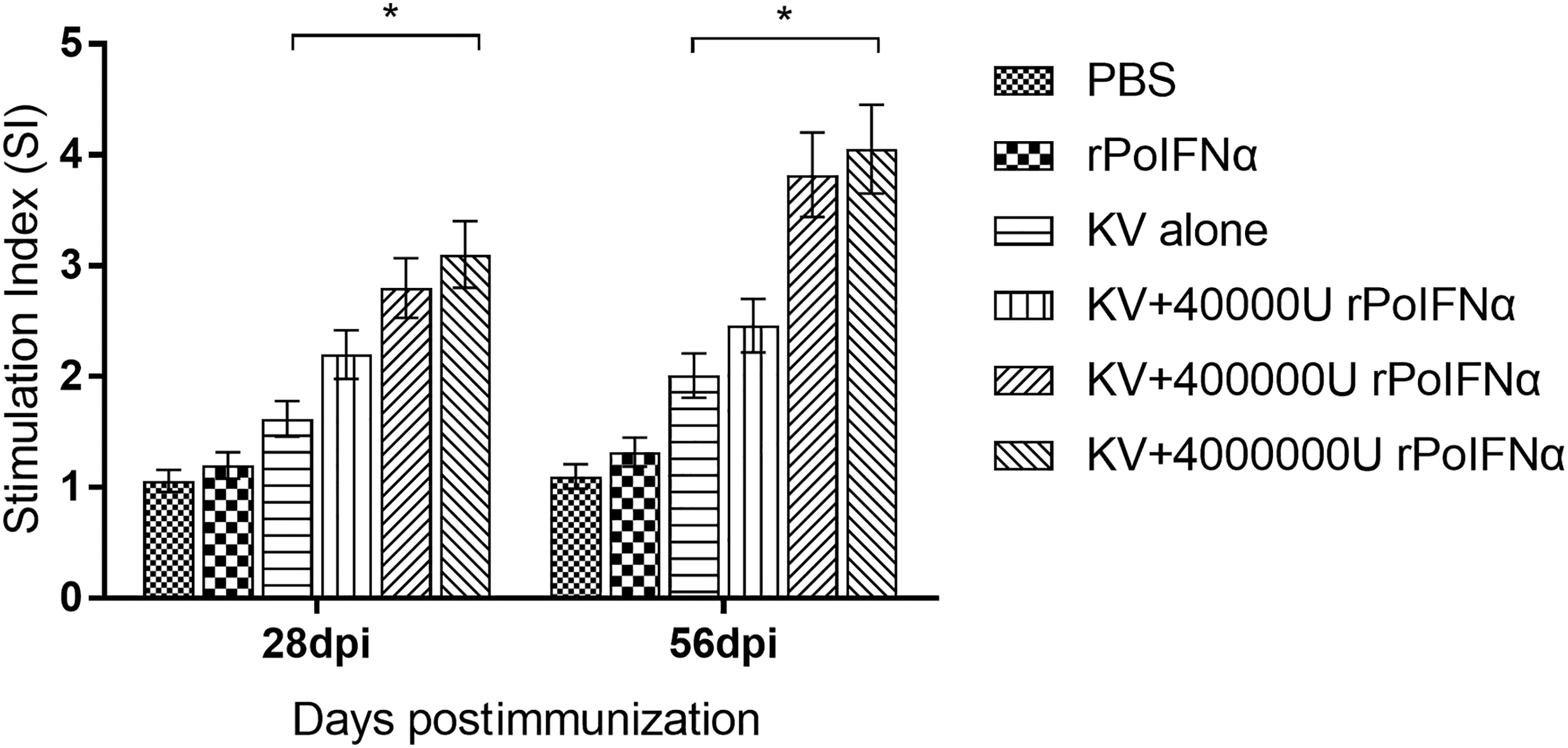
Proliferation of PBMC in different groups of experimental pigs. Each bar represents the SI values determined in triplicate, and the data are expressed as the group average ± SD. Concanavalin A was used as a positive control; *the difference between the two groups is significant (p < 0.05). SI, stimulation index.
Compared with basic line SI value (0.34 ± 0.19) and positive control SI value (6.18 ± 2.53), immunization with KV alone produced a deficient specific lymphocyte proliferative response before 14 dpi, whereas in the groups of KV+400,000 U rPoIFNα or KV+4,000,000 U rPoIFNα immunized pigs a stronger proliferative response were induced. At 14 and 28 dpi, pigs from groups immunized with KV+400,000 U rPoIFNα and KV+4,000,000 U rPoIFNα produced a higher SI than those from groups immunized with PBS and the KV alone. On day 56 after initial immunization (56 dpi), the SI of the PRRSV KV+4,000,000 U rPoIFNα group reached the highest level, which was significantly higher than those of the groups of PBS and the KV alone (p < 0.05).
In the entire immunized population, the SI of the group immunized with KV+4,000,000 U rPoIFNα was higher than those of KV+40,000 U rPoIFNα group and the KV+400,000 U rPoIFNα group (Fig. 7). These results confirmed that the addition of rPoIFNα improves the CMI response of KV in pigs.
Analysis of lymphocyte subsets in pig blood
The changing profiles of CD3 +, CD4 +, and CD8 + cell populations examined by flow cytometry are shown in Figure 8. CD3 +, CD4 + cell, and CD8 + counts increased slightly in most groups at the early stage after the initial immunization. These populations are gradually increased in the pigs' blood of each group at 28 dpi, then increased significantly at 56 dpi after booster immunization (Supplementary Figure S1). Similarly, the changes of lymphocyte subsets (CD3 +CD4 −CD8 +IFN-γ +, CD3 +CD4 +CD8 −IFN-γ +, and CD3 +CD4 + CD8 +IFN-γ+) cells counts showed comparative growth trend.
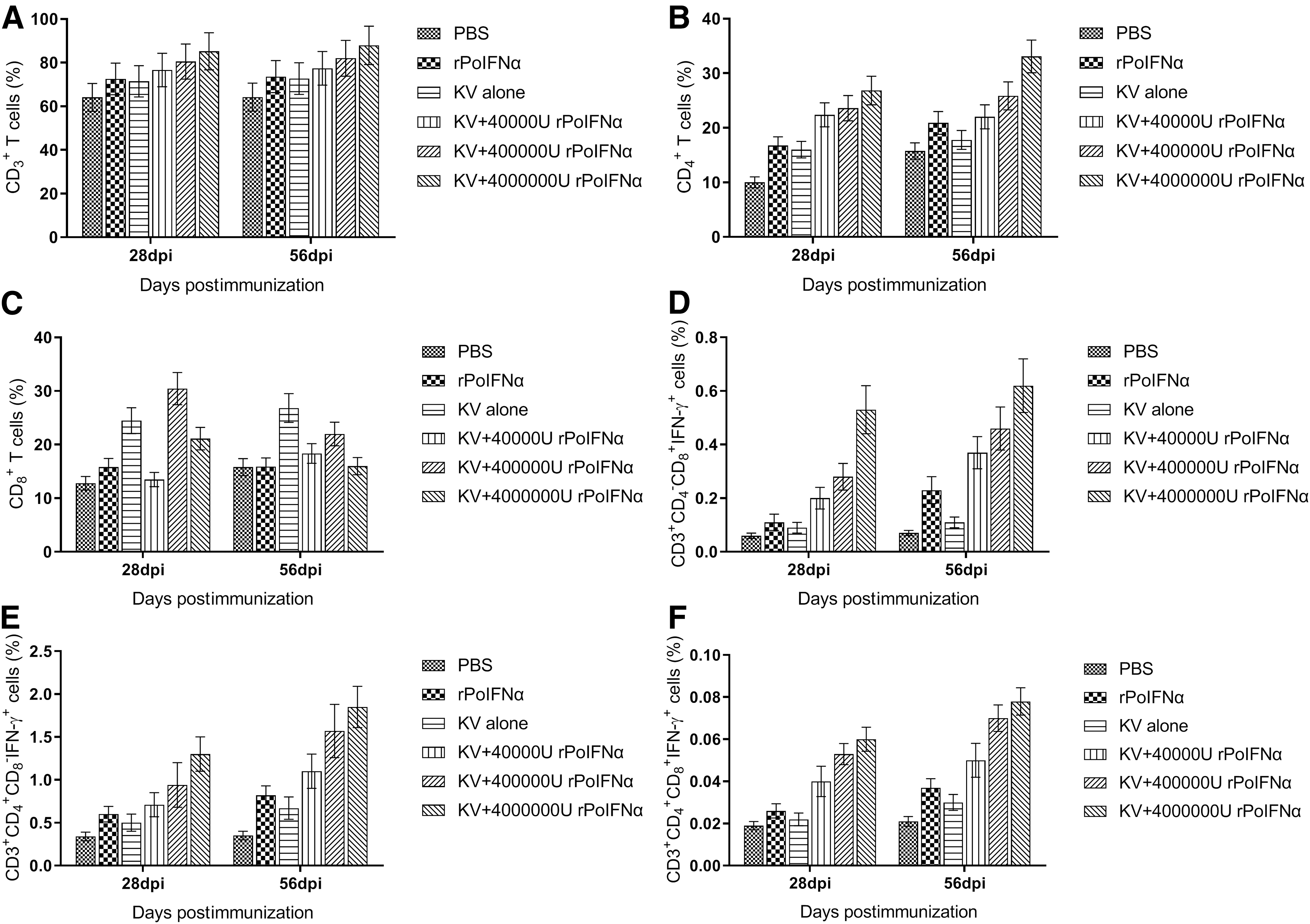
PRRSV antigen-specific activated lymphocyte subsets in the blood of the immunized pigs in each group during the period of initial immunization and booster immunization.
Discussion
PRRS is a problem for >10 years in the global pig industry, and the PRRS KV vaccine has been used clinically to inhibit the spread of the disease. Although the PRRS KV vaccine has shown the ability to induce rapid IgM and IgG responses, the antibodies generated are subneutralized. Scientific observations found that the sub-NAs enhanced viral replication in macrophages, which may play an important role in participating in the development of PRRS.
Both KV and MLV have been approved for PRRSV control. KV vaccine contains adjuvant, which is contrary to MLV vaccine. Although MLV vaccines provide better protection than KV vaccines, Its protection is only greater in homologous cases, and there is a risk of virulence recovery in terms of safety (16,26). Some western countries, such as the United Kingdom, allow only PRRSV KV vaccines when used in breeding animals. Also, the main inoculation targets of some commercial KV vaccines, including the Progressis® Vaccine produced by Merial Corporation, are sows and gilts, and they are intended to reduce reproductive disorders caused by PRRSV infection (15).
Studies have shown that multiple inoculations of KV in serum-positive sows could enhance the immunity against PRRSV (28). For example, the Promise® PRRSV KV vaccine (Intervet, Millsboro, DE) was injected twice into PRRSV-infected pigs, although it did not decrease viral shedding, it did increase the production of IFN-γ and NA titers (16-fold), which are consistent with the results of our study (27).
As part of humoral immunity, the production of NA plays a major role in the removal of PRRSV and the prevention of virus infection (23). CMI is also crucial for the host immune responses against PRRSV infection (38). However, previous IFNα studies indicated that only a solo CMI response was insufficient for cross-protection (4,22). Protective immunity to heterologous PRRSV strains infection may require both augments of CMI and antibody production (4). It strongly suggests that PRRSV NA antibody production and virus-specific cellular immune response work together to promote virus clearance, indicating that innovative formulations of noninfectious vaccine and delivery systems are needed to trigger humoral and cellular responses to effectively control PRRS (1,29,34,36,41).
Our results indicate that rPoIFNα has immunopotentiating potential when combined with the PRRSV KV vaccine. Specifically, we demonstrate that a coadministration of rPoIFNα with the killed PRRSV-induced both Th1 and Th2 immune responses in pigs. rPoIFNα was shown to have good safety. After its immunization, the clinical manifestations of pigs were not affected, including weight gain (Fig. 1) and body temperature (Fig. 2).
After vaccination, we also determined concentration changes in the PRRSV-specific antibodies, NAs, cytokines IL-4, and IFN-γ. We found that (i) for PRRSV-specific antibody titers, the production time of PRRSV-specific antibodies in those rPoIFNα+KV groups was earlier than that in the KV alone group. The production levels of PRRSV-specific antibodies in those rPoIFNα+KV groups were also higher than those in the KV alone group, suggesting that rPoIFNα increase the PRRSV-specific antibody responses triggered by KV vaccine (Fig. 3). (ii) For NA titers, pigs vaccinated with rPoIFNα+KV were significantly different from pigs vaccinated with KV alone vaccine, indicating that rPoIFNα may increase NA levels; these augments may contribute to humoral immunity and anti-PRRSV protections (Fig. 4).
(iii) For cytokines, the addition of rPoIFNα significantly upregulated the generation of IFN-γ and IL-4 and improved the cell-mediated response to the PRRSV KV vaccine (Figs. 5 and 6 ). (iv) For lymphocyte proliferation activity, our data demonstrated that the PRRSV-specific lymphocyte proliferation responses in the KV+rPoIFNα coadministration groups were significantly higher than that in the KV alone immunization group (p < 0.05) (Fig. 7). Since the lymphocyte proliferative response is usually related to CMI, it indicates that the combination of IFNα and KV can enhance the CMI of the PRRSV KV vaccine.
We also discovered that rPoIFNα at a dose of 400,000 U or 4,000,000 U significantly enhanced the porcine immune response when formulated with the PRRSV KV vaccine. Specifically, the supplementation of rPoIFNα to the PRRSV KV vaccine enhanced humoral and cellular responses compared with the PRRSV KV alone group. Our results also showed that the immunopotentiating effect of rPoIFNα was greater than the dose of 400,000 U at the dose of 4,000,000 U, which suggests that rPoIFNα can enhance the immune response in a dose-dependent manner in pigs. The best result with 4,000,000 U is more durable. Our finding is consistent with the results reported by Tovey et al., based on the observations that when IFNα is used together with inactivated influenza vaccine, the levels of IgG and IgA in body fluids increase significantly in a dose-dependent manner (35).
Conclusion
In conclusion, when rPoIFNα was prepared at doses of 400,000 U or 4,000,000 U, rPoIFNα had an adjuvant effect. The antibody production window period of the PRRSV KV+rPoIFNα group was earlier than that of KV alone group. Also, the combined use of rPoIFNα with KV vaccine can enhance the antibody titer, the lymphocyte SI, the secretion of IFN-γ and IL-4, and the lymphocyte subsets populations compared with the single use of PRRSV KV alone vaccine after immunization. To our knowledge, this is the first in vivo study of the potential of rPoIFNα in promoting immune response to PRRSV KV vaccine in pigs. The results suggest the appropriateness of rPoIFNα as an adjuvant for immune stimulation in farm animal vaccines. However, further field challenge assays are needed to test the protective effect of rPoIFNα+KV to pigs.
Compliance with Ethical Standards
All animal experimental procedures performed in this study are conformed to the policies of the National Institutes of Health Guide for the Care and Use of Laboratory Animals and were approved by the Bioethics Committee of Anhui Medical University.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported by the programs from the 2017 Wuhu Science and Technology Plan Project (Grant No. 2017yf01), Natural Science Foundation of Anhui Province (Grant No. 1808085MC75), National Key R&D Program of China (Grant No. 2017YFD0501000 and 2017YFD0500906), and Scientific research activities of postdoctoral researchers in Anhui (Grant No. 2017B194). The funders did not participate in the study design, data collection, data analysis, publish decision, and article writing.
Supplementary Material
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.