
Abstract
Women with persistent human papillomavirus (HPV) infections have a high risk of developing cervical cancer (CaCx). HPV-16 alone accounts for more than 60% of CaCx worldwide. Most of the HPV infections are transient and only a subset of women develop persistent HPV-16 infection. Many studies have shown associations of different human leukocyte antigen (HLA) alleles with HPV-mediated CaCx, but there are only a few studies globally that relate to persistent HPV-16 infection. Furthermore, such studies from India are sparse. Hence, we investigated the association of HLA-A, B, DRB, and DQB alleles with persistent HPV-16 infection and HPV-16-positive CaCx in south India (Tamil Nadu). HPV-16 persistent infection was observed in 7% of normal women. A total of 50 women with HPV-16-positive CaCx, 21 women with HPV-16 persistent infection, and 74 HPV-16-negative normal women were recruited for this study. Low-resolution typing of HLA-A, B, DRB, and DQB alleles was performed. HLA-B*44 and DRB1*07 showed a significant association with persistent HPV-16 infection (odds ratio, p-value = 26.3, 0.03 and 4.7, 0.01, respectively). HLA-B*27 and DRB1*12 were significantly associated with both HPV-16+ CaCx and persistent HPV-16 infection (23.8, 0.03; 52.9, 0.01; 9.8, 0.0009; and 13.8, 0.009; respectively). HLA-B*15 showed a negative association with HPV-16-positive CaCx (0.1, 0.01), whereas DRB1*04 exhibited protection to both HPV-16-positive CaCx and persistent HPV-16 infection (0.3, 0.0001 and 0.1, 0.0002, respectively). Thus, we show HLA allelic association with HPV-16 infection in Tamil Nadu. Larger studies on high-resolution HLA typing coupled with HPV-16 genome diversity will offer further insights into host/pathogen genome coevolution.
Introduction
Cervical cancer (CaCx) is the fourth frequent cancer accounting for 6.6% of all female cancers worldwide with 570,000 new cases in 2018 and ∼311,000 deaths in 2018 (68). A majority of the CaCx cases are diagnosed in women who are <50 years old and more than two-thirds of the women suffering from this disease are identified in the less developed regions of the world. It is seen that most of the CaCx cases arise in Southeast Asia, Latin America, and sub-Saharan Africa (64). About 120,000 of the cancer of cervix uteri cases occur in India, which accounts for one-fourth of the global burden of the CaCx (16). The major etiological agent for the development of CaCx is sexually acquired human papillomavirus (HPV) infection.
The motive behind screening is for detecting the presence of any precancerous lesions and if left untreated, it may progress to CaCx. It is recommended that every woman in the age group of 30–49 years undergoes HPV screening at least once in her lifetime. The screening procedures include cytology, visual inspection with acetic acid, and HPV testing (59). Papillomaviruses are small, nonenveloped, double-stranded DNA tumor viruses that infect the keratinocytes in the basal layers of the stratified squamous epithelium (19). The viral genome consists of eight major genes broadly classified into two: Early (viral replication and oncogenesis) and Late (capsid). There are more than 150 types of HPV that have been identified based on the sequence variation of L1 capsid gene (17,21). The HPV types are broadly classified into two types as follows: (1) low risk (nononcogenic): 6, 11, 42, 43, 44, and 55 that cause anogenital warts and (2) high risk (oncogenic): 16, 18, 31, 33, 35, 39, 45, 51, 52, 56, 58, 59, 66, and 68 that may lead to CaCx and other cancers.
HPV type 16 (HPV-16) has been phylogenetically classified into four main lineages, A–D, which further branch into four sublineages as follows: A1–4, B1–4, C1–4, and D1–4 (14). A study by Clifford et al. (14) has described the phylogenetic distribution of HPV-16 (sub) lineages with respect to the relative CaCx risk. It was found in the study that D lineages (specifically D2 and D3) and A3 and A4 sublineages were exhibiting increased CaCx risk rather than the widespread A1 lineage.
The high-risk types, HPV-16 and -18, account for more than 80% of CaCx and precancerous lesions. HPV-16 has a higher rate of infection and infects almost 60% women worldwide (34). In cytologically normal women, HPV-16 is seen at a frequency of 4–6% (8). Not all individuals who acquire an infection develop the disease (44). Only about 10–15% of HPV-infected individuals establish lifelong persistent infection and a subset of this progresses to invasive CaCx (67). Those with persistent HPV infection are more prone to acquire epithelial cell abnormalities and subsequently develop cancer at the site of infection. An effective immune response promotes the spontaneous clearance of the virus, while a compromised immune response often leads to the pathological process by further developing into CaCx.
The factors involved in the natural clearance of the virus, specifically in persistent HPV infection, have been a major area of interest. The host, pathogen, and environmental factors primarily dictate the outcome of an infectious disease. The classical human leukocyte antigen (HLA) system also called as “the eye of the immune system” is one such genetic marker, in which certain alleles were shown to be associated with reduced clearing of HPV infection and the subsequent progression to CaCx (56). The HLA in humans is encoded by the major histocompatibility complex (MHC) genes and these cell surface glycoproteins present the antigen to the T cell receptor activating the T cell. These highly polymorphic genes belong to three subgroups: namely, class I (A, B, and C), class II (DR, DQ, and DP), and class III (TNF-α, complement proteins). The class I HLA molecules are expressed on all nucleated cells and present viral and altered self-antigens to CD8+ T cells. The class II HLA molecules are expressed on antigen-presenting cells and present exogenous antigens to CD4+ T cells, leading to the coordination and regulation of effector cells (69).
The human MHC locus is recognized as the most polymorphic region of the human genome. This high degree of functional polymorphism in the HLA genes makes them the quintessential set of “candidate genes” in disease susceptibility among humans (22).
Previous studies have shown positive and negative associations of HLA alleles with different degrees of HPV infection in different populations. The literature survey of HLA association with HPV-mediated CaCx in India revealed only four studies from the east and north Indian population. Das Ghosh et al. found B*40 allele to be significantly overrepresented in CaCx cases than controls, whereas B*15 was significantly underrepresented among the east Indian women (15). Gokhale et al. showed HLA- B*37, B*58, DRB1*03, and DQB1*02 alleles to be significantly associated with increased risk of CaCx, while HLA- B*40, DRB1*13, DQB1*06, and DQB1*03 showed a significant resistance to CaCx in the north Indian population (23). A study by Kohaar et al. (28) from north India showed that HLA-DRB1*15 and the haplotype DRB1*15-DQB1*06 to be predisposing factors in CaCx or precancer development, whereas DRB1*04 allele exhibited susceptibility to cervical precancer lesions. DRB1*13 allele in the same population showed a strong protection to HPV infection (28). A study by Bhattacharya and Sengupta has shown HLA-DQB1*03 homozygosity and HLA-B*07 to be a predisposing factor in HPV-16/18-related CaCx in women from east India (6).
There is only one study from Tamil Nadu that associates HLA alleles with CaCx, but it did not address the association between the high-risk HPV types and HLA (51). There was no study associating HLA alleles with persistent HPV-16 infection from India. Hence, the present study aims to identify the association of HLA-A, B, DRB, and DQB genetic polymorphisms with persistent HPV-16 infection and HPV-16-mediated CaCx in Tamil Nadu, India.
Materials and Methods
Subjects
The present study investigated the association of HLA-A, B, DRB, and DQB genetic polymorphisms with persistent HPV-16 infection in a hospital-based case/control study. A total of 145 normal women were recruited for the study from the Kovai Medical Centre and Hospital, Coimbatore, Tamil Nadu.
A detailed questionnaire was provided to each individual and the response was received appropriately. Cervical swab was obtained from all the women. A second swab was collected from those who turned up positive for HPV-16 after 4 months. The time period of 4 months was followed as described in a few previous studies (12,20,26,30,32,52,65). Those who were HPV-16-positive in both the swabs were considered to have persistent HPV-16 infection and the rest were treated as controls. It was taken care that the control samples used in the study were age, region, and ethnicity matched. The inclusion criterion for the cervical swab collection was normal reproductively active females, while the exclusion criteria were pregnant and menstruating females and those with any other clinical disease. In addition, 70 formalin-fixed paraffin-embedded (FFPE) tissue blocks of CaCx cases were obtained from the GVN Cancer Hospital, Trichy, Tamil Nadu. The present study was carried out in accordance with the principles of the Helsinki Declaration and the ethical clearance for the study was obtained from the respective tertiary care hospitals.
Genomic DNA extraction
The cervical swabs were added to a tube containing 2 mL cell lysis buffer (100 mM Tris [pH 7.6], 40 mM ethylenediaminetetraacetic acid [pH 8.0], 50 mM NaCl, 0.2% sodium dodecyl sulfate, and 0.05% sodium azide) during the time of sample collection. Genomic DNA from cervical swab was extracted using a standard salting-out method (39). DNA from FFPE tissue blocks was extracted using the QIAamp DNA FFPE Tissue Kit (cat no. 56404) following the manufacturer's protocol.
Screening of HPV
The extracted DNA samples were screened for the presence of HPV-16 by a custom-made TaqMan assay targeting the L1 gene. The following set of primers and probes were used for the real-time polymerase chain reaction (PCR) assay (27).
Forward primer: 5′-TTGTTGGGGTAACCAACTATTTGTTACTG-3′
Reverse primer: 5′-CTCCCCATGTCTGAGGTACTCCTTAA-3′
Probe: 5′-FAM-GTCATTATGTGCTGCCATATCTACT-MGB-3′
Thermocycling was performed in ABI7900 real-time PCR system. The cycling condition was initial denaturation of 96°C for 5 min, followed by 40 cycles of 96°C for 15 sec and 60°C for 30 sec (annealing and extension combined). Fluorescence data for each cycle were collected at 60°C. Known HPV-16-positive and known HPV-16-negative samples were used as controls during the reaction.
HLA typing
HLA- class I (A and B) and class II (DRB and DQB) alleles were genotyped by low-resolution PCR/sequence-specific priming (PCR-SSP) for the study samples (9,10,41,63). The alleles were identified by running the PCR amplicons along with a DNA marker.
Statistical analyses
The association of HLA-class I and II alleles with HPV-16-positive CaCx and persistent HPV-16 infection was determined by calculating the odds ratio (OR) with Bonferroni correction for multiple comparisons. A p-value of <0.05 was considered to be statistically significant. All statistical analyses were performed using EpiTools package in R v3.4.1 (62). HLA haplotypes were estimated from the frequency data using the maximum likelihood method in Arlequin v3.5 (1).
Results
The present study describes the association of HLA-A, B, DRB, and DQB alleles with HPV-16 infection and HPV-16-mediated CaCx. A total of 145 women with normal cervical cytology were enrolled, of whom 21 had persistent HPV infection as evidenced from the presence of HPV-16 after a 4-month follow-up. Women who turned out negative in the follow-up and those who did not show HPV-16 in the initial screening were considered controls. The time period to define persistent infection varied considerably between studies ranging from 3 to 24 months. Studies such as Byun et al. (12), Hoffman et al. (26), Kudoh et al. (32), Dutta et al. (20), Wang et al. (65), Koshiol et al. (30), and Rodríguez et al. (52) have used 3 months of follow-up to define persistent infection. Of the 70 FFPE CaCx blocks, 50 were positive for HPV-16. Thus, a total of 74 normal women, 50 HPV-16-positive CaCx cases, and 21 women with persistent HPV-16 infection were enrolled for the study. The patients (HPV-16 persistent infection and CaCx) had a mean age of 36.1 ± 14.9 years and the controls were 38.1 ± 10.6 years.
Distribution of HLA-class I alleles in the study samples
We identified a total of 14 different HLA-B alleles in the study samples. HLA-B*35 was the commonest allele in the study samples (Fig. 1, Supplementary Table S1). HLA-B*27 showed the highest association with both HPV-16-positive CaCx and persistent HPV-16 infection (OR, p-value = 23.8, 0.03 and 52.9, 0.01, respectively) (Table 1). Similarly, HLA-B*44 showed a significant association with persistent HPV-16 infection (OR, p-value = 26.3, 0.03). This allele showed an OR of 13.8 in HPV-16-positive CaCx but was not statistically significant as the lower bound of the 95% CI was below 1 (OR, p-value = 13.8, 0.08, 95% CI = 0.74–260.16). These two alleles (HLA-B*27 and HLA-B*44) were completely absent in the control samples. HLA-B*57 showed a significant association with HPV16-positive CaCx (OR, p-value = 5.2, 0.03). HLA-B*15 showed a significant negative association with HPV-16-positive CaCx (OR, p-value = 0.1, 0.01).
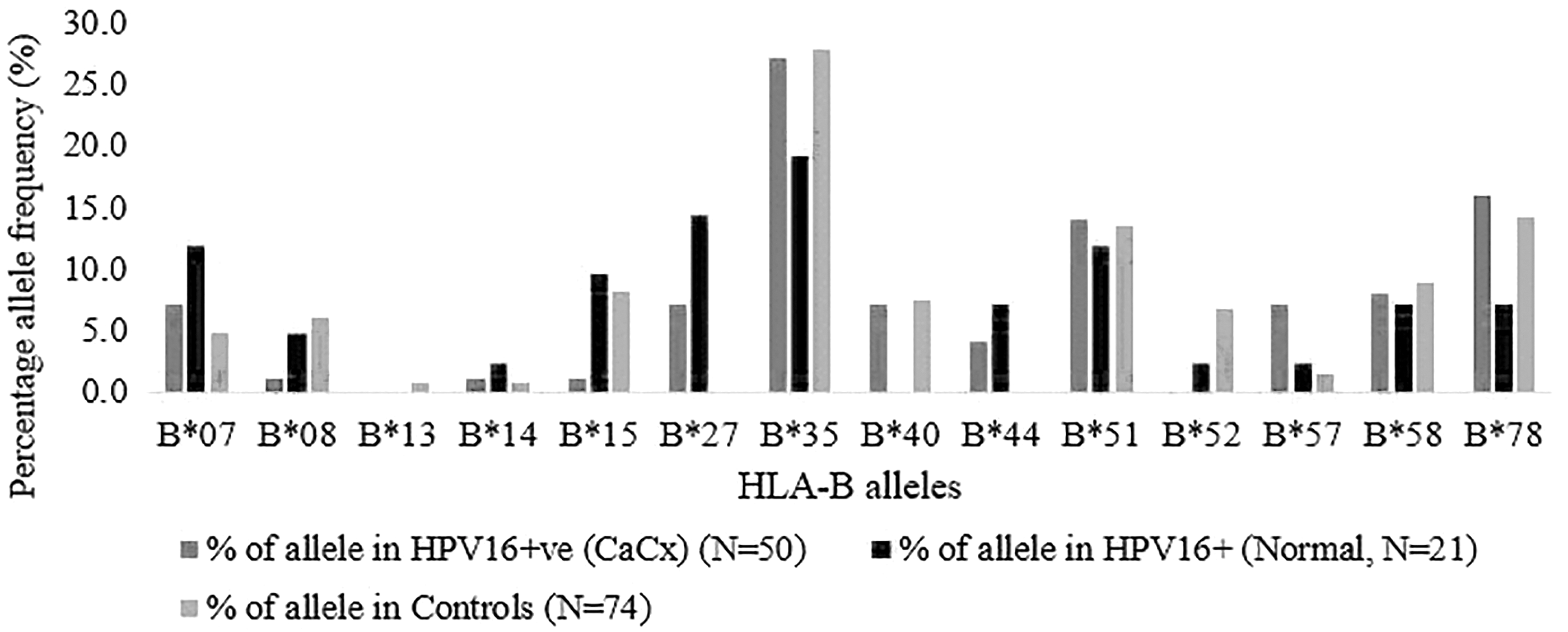
Percentage of allele frequency of HLA-B alleles in HPV-16-positive CaCx, persistent HPV-16 infection (normal), and controls. CaCx, cervical cancer; HLA, human leukocyte antigen; HPV, human papillomavirus.
HLA-B Allele Frequencies of HPV 16-Positive Cervical Cancer, Persistent HPV 16 Infection, and Controls
CI, confidence interval; HLA, human leukocyte antigen; HPV, human papillomavirus; OR, odds ratio; p-value, probability value of Fisher's exact test.
Seventeen major HLA-A alleles were genotyped in all the samples, of which we identified 14 HLA-A alleles in the study samples. HLA-A*02, A*01, A*24, and A*11 were the commonest alleles (22.8%, 16.9%, 13.1%, and 12.8% respectively) in the study samples (Supplementary Table S2). HLA-A*11 showed an association with HPV-16-positive CaCx cases (OR, p-value = 2.3, 0.04) (Table 2). On the contrary, this allele did not show a significant association with persistent HPV-16 infection. HLA-A*69 and A*32 were found at a frequency of 4.8% and 2.4% in persistent HPV-16-positive women, while these alleles were absent in the controls (OR and p-value = 18.3, 0.06 and 10.7, 0.15, respectively). HLA-A*01 was present in 20.3% of the controls and 7.1% of persistent HPV-16-positive women (OR = 0.3, p-value = 0.04, 95% CI = 0.1–0.96). This has to be interpreted with caution as the upper bound of the 95% CI borders with no association, that is, OR of 1.
HLA-A Allele Frequencies of HPV 16-Positive Cervical Cancer, Persistent HPV 16 Infection, and Controls
Distribution of HLA-class II alleles in the study samples
In the present study, we identified 13 HLA-DRB1 alleles and 4 HLA-DQB1 alleles. HLA-DRB1*15 and DRB1*04 were the commonest alleles (Fig. 2, Supplementary Table S3). DRB1*12 showed a very significant association with both HPV-16-positive CaCx and persistent HPV-16 infection (OR, p-value = 9.8, 0.0009 and 13.8, 0.009, respectively) (Table 3). HLA-DRB1*15 showed a significant association with HPV-16-positive CaCx but not with persistent HPV-16 infection (OR, p-value = 1.9, 0.03 and 1.2, 0.59, respectively). DRB1*07 showed a significant association with persistent HPV-16 infection but not with HPV16-positive CaCx (OR, p-value = 4.7, 0.01 and 2.6, 0.07, respectively). The lower bound of 95% CI of the OR of HPV-16-positive CaCx was 0.92, which is <1. When both the groups of cases were combined as HPV-16-positive, the association was significant (OR, p-value = 3.15, 0.02) (Supplementary Table S3). On the contrary, HLA-DRB1*04 showed a very significant negative association with both HPV-16-positive CaCx and persistent HPV16 infection (OR, p-value = 0.3, 0.0001 and 0.1, 0.0002, respectively) (Table 3). This allele was observed in 31.1% of the controls, whereas it was observed in 10% of HPV-16-positive CaCx cases and 4.8% of persistent HPV-16 infection.
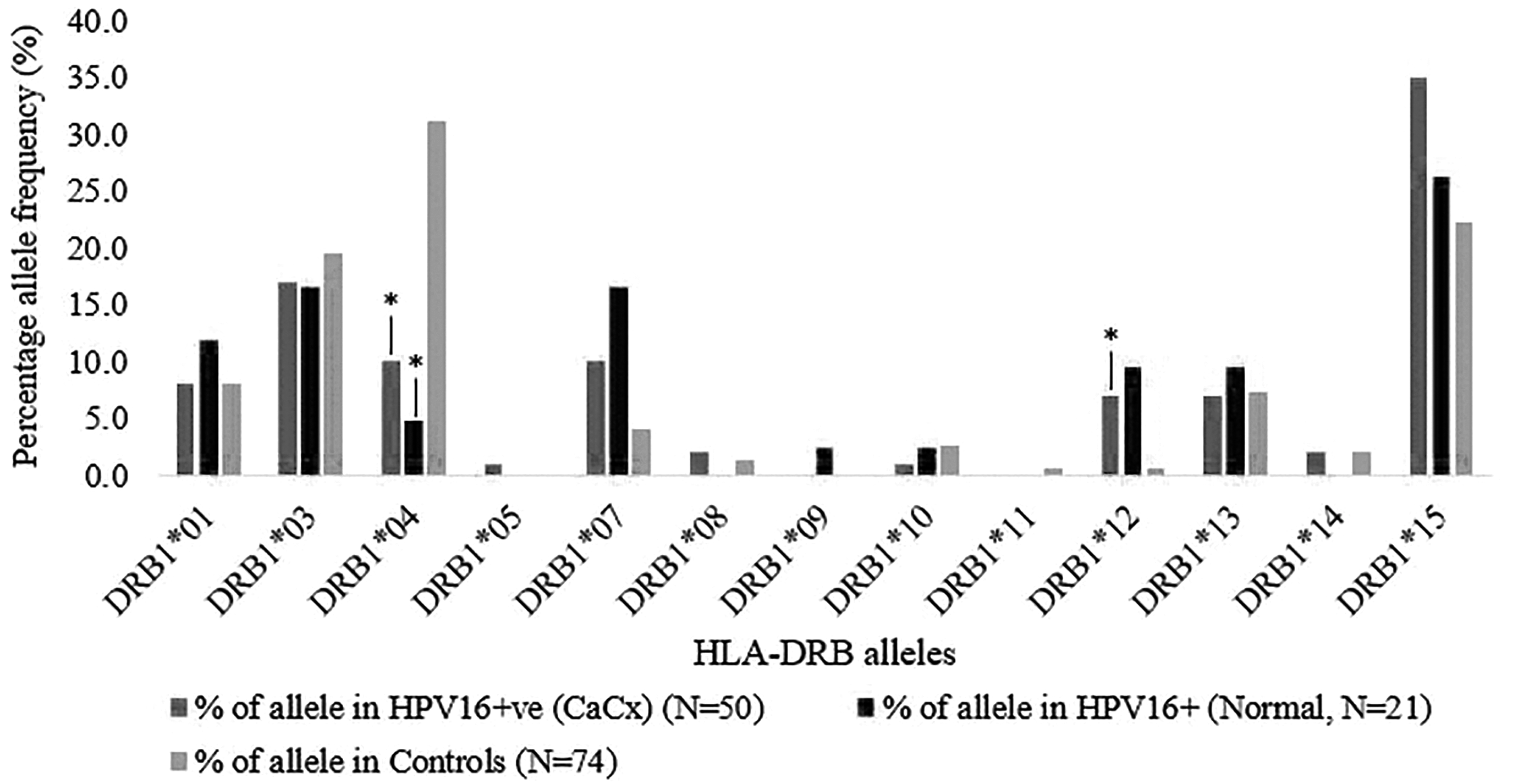
Percentage of allele frequency of HLA-DRB alleles in HPV-16-positive CaCx, persistent HPV-16 infection (normal), and controls. *Indicates odds ratio p < 0.05.
HLA-DRB and DQB Allele Frequencies of HPV 16-Positive Cervical Cancer, Persistent HPV 16 Infection, and Controls
The p-value of DRB1*12 was statistically significant after Bonferroni correction (p c = 0.0117, respectively) in HPV 16-positive CaCx, while DRB1*04 showed a significant negative association with both HPV 16-positive CaCx and persistent HPV 16 infection (p c = 0.0013, 0.0026).
CaCx, cervical cancer.
Four major HLA-DQB alleles were identified in the study samples and no major association was identified. Sequence-based typing of the HLA-DQB alleles may further identify subtypes that may have associations with the HPV-16 infection.
Association of HLA-class I and class II haplotypes with HPV-16 infection
HLA haplotypes were estimated from the frequency data using the maximum likelihood method. We estimated two 2-loci haplotypes namely HLA A-B & HLA DRB1-DQB1 and one 3-loci haplotype namely HLA A-B-DRB. A total of 84 HLA A-B haplotypes were identified. HLA A*01-B*35 and A*02-B*35 were the commonest haplotypes (9% and 7.6%, respectively) in the study samples (Supplementary Table S4). The haplotype A*02-B*27 showed a significant association with persistent HPV-16 infection (OR, p-value = 26.32, 0.03) (Table 4). The haplotype A*26-B*08 showed a negative association with total HPV-16 infection (HPV-16-positive CaCx and persistent HPV-16 infection combined) (OR, p-value = 0.14, 0.02) (Supplementary Table S4).
HLA-Class I and II Haplotype Frequencies of HPV 16-Positive Cervical Cancer, Persistent HPV 16 Infection, and Controls
We identified 33 MHC class II DRB1-DQB1 haplotypes. The haplotypes DRB1*15-DQB1*03, DRB1*04-DQB1*03, and DRB1*03-DQB1*03 were the commonest haplotypes in the study samples (13.8%, 10.7%, and 9%, respectively) (Supplementary Table S4). The haplotypes DRB1*07-DQB1*03 showed significant association with total HPV-16 infection (OR, p-value = 4.1, 0.05) (Supplementary Table S4). The haplotype DRB1*15-DQB1*03 showed significant association with HPV-16-positive CaCx (OR, p-value = 2.1, 0.05) (Table 4). The haplotype DRB1*04-DQB1*03 showed a significant negative association with both HPV-16-positive CaCx and persistent HPV-16 infection (OR, p-value = 0.34, 0.01 and 0.14, 0.01, respectively). Among the 3-loci haplotypes, A*02-B*27-DRB1*01 alone showed a significant association with persistent HPV-16 infection (OR, p-value = 26.3, 0.03).
Discussion
Persistent infection with HPV-16 is the primary etiological factor of cervical carcinogenesis. HPV-16 infects basal epidermal layers of the mucosal and cutaneous skin via microaberrations. In the basal layers of the epithelia, the viral genome is maintained in a low copy number in the keratinocytes. Further infection of these cells in the basal layer leads to the activation of a cascade of viral gene expression, thereby resulting in the amplification of the viral genome (36,71). The time of development of an initial infection to the progression of the disease varies from several days to months. Various innate and adaptive immune responses play a crucial role in the spontaneous clearance of the virus. CD4+ T cell response to HPV E2 and E6 antigens are important in clearing the low-grade HPV infection, whereas CD4+ T cell response against E7 antigen results in the prevention of neoplasia (18,60). The extent and type of T cell activation depend critically on the type and expression of HLA (40).
HLA promotes the activation of an effective immune response against the cancer cells by means of selecting the peptides that are generated from the mechanism of processing of foreign antigens and thereby displaying them on the cell surface for T cell recognition. HLA class I molecules are present on all nucleated cells, whereas class II molecules are expressed on antigen-presenting cells. The class I molecules present the intracellular antigens to cytotoxic T cells, while the class II molecules present extracellular antigens to T helper cells. Based on the stimulus of T cell activation, the T helper lymphocytes divide into three subtypes and each of them produces its unique pattern of cytokines, namely T helper 1 (Th1), T helper 2 (Th2), and T helper 17 (Th17). Cytokines play a vital role in the regulation of viral transcription, in inflecting viral replication and in safeguarding the host against HPV infection. One of the possible reasons of the variation in the positive and negative associations of HLA with HPV-16 infection is due to the increased levels of Th1 (T helper cells) cytokines, and studies across the globe have shown an association of Th1 cells with persistent HPV infection, CaCx, and metastasis (3,46).
The present study showed significant positive associations (HLA-A*11, B*27, B*44, B*57, DRB1*12, DRB1*15, and DRB1*07) and negative associations (HLA-B*15 and DRB1*04) of HLA alleles with HPV-16-positive CaCx and persistent HPV-16 infection (Tables 1–3). Of the various class I HLA alleles, HLA-B*07 has remained the commonest allele globally and has been reported to be a high-risk allele in familial CaCx (47) and in high-grade squamous intraepithelial lesions from the Costa-Rican population (66). In a study consisting of British women with HPV-16-positive vulval intraepithelial neoplasia (VIN), HLA-B*07 was found to show similar increased frequencies in both HPV-16-positive CaCx and VIN (15). A meta-analysis study showed the same allele to play a protective role against chronic hepatitis B infection (55). In India, B*07 was observed to be significantly overrepresented in cytologically normal subjects and was 20.5% significantly higher in HPV-16-positive CaCx when compared with the controls (6). In the present study, B*07 was found to show a positive association with both HPV-16-positive CaCx and persistent HPV-16 infections but was not statistically significant (OR, p-value = 1.5, 0.5 and 2.7, 0.12, respectively).
The present study showed the highest and significant association of HLA-B*27 with HPV-16-positive CaCx and persistent HPV-16 infection (Table 1). The p-values in both the groups were statistically significant (0.03 and 0.01, respectively). Positive associations of HLA-B*27 with HPV-16-positive CaCx were observed in southern Chinese (13) and Japanese women (53). HLA-B*44 and B*57 were found to be significantly associated with persistent HPV-16 infection and HPV-16-positive CaCx in the present study. HLA-B*44 also showed susceptibility to HPV-16-positive CaCx in Swedish and Dutch populations (31,71). A significantly positive association of HLA-B*57 in the present study corroborates with that of HLA-CaCx association studies on Swedish, Mexican, and Dutch populations (7,25,71). HLA-B*57 allele was also found to be in 11% of HIV-infected children in the Indian population and is a widely accepted molecular marker for hypersensitivity to abacavir (38).
In the present study, HLA B*15 showed a significant negative association with HPV-16 infection (Table 1). This observation is in line with the study by Das Ghosh et al. that HLA B*15 in the east Indian population was underrepresented among HPV-16-positive women thereby playing a potential role in the spontaneous regression of HPV infection (15). This allele also showed resistance toward HPV-16-positive CaCx in the southern Chinese and Swedish populations (13,71). It is interesting to note that the positive and negative associations of HLA-B alleles (B*44, B*57, and B*15) with HPV-16-positive CaCx and persistent HPV-16 infection in the present study correlated with our previous meta-analyses study (5).
In the present study, only two HLA-A alleles (A*11 and A*01) were associated with HPV-16-positive CaCx and persistent HPV-16 infection. HLA-A*11 showed a significant association with HPV-16-positive CaCx. Only a marginal positive association of A*11 was observed in a Dutch population (31) with CIN grade III. In the present study, it was observed that HLA-A*01 showed a significant negative association with persistent HPV-16 infection (OR, p-value = 0.3, 0.04) (Table 2), and a similar protective association of the same allele was seen in Dutch and Finnish populations (31,61). HLA-A*69 and A*32 alleles showed susceptibility toward persistent HPV-16 infection but were not statistically significant in the present study. HLA-A*69 is a split antigen that belongs to the serotype A28 and this serotype showed a negative association with HPV-16-positive women of Swedish and Dutch origin (31,71).
A study by Rathika et al. (51) was the only study available in literature that associated HLA alleles with CaCx in women from Tamil Nadu. However, this study did not look into the HPV genotype in the cases. This study showed HLA alleles B*15, B*07, DRB1*04, DRB1*15, DQB1*0301, and DQB1*0601 to be associated with CaCx. We found that our results were in contrast with Rathika et al. We found no association of B*15, a nonsignificant association of B*07, and a negative association of DRB1*04. To our surprise, we found the overall frequency of the HLA alleles to be markedly different between the two studies. The overall HLA-A*01 frequency in our study samples was found to be 16.9% and this frequency was similar to previous studies from the same region that showed frequency estimates of 13.1%, 14.1%, and 17% (5,32,36). However, Rathika et al. reported this allele at a frequency of only 6.3%. Similar differences in the frequency of A*03 allele between our study (6.5%) and that of Rathika et al. (21.5%) were observed. This allele in previous studies was observed to be 6.3% (45), 6.9% (49), and 7% (4). Similar drastic variations in the frequency of HLA-A*11, A*02, and A9 serotypes (A*23/A*24) were observed between our study and that of Rathika et al. In all these alleles, the overall frequency estimates of the present study were similar to that of the previous three studies from the same region (4,45,49). This may be the reason for the contrasting associations identified between the present study and that of Rathika et al.
Among the HLA-class II alleles identified in the present study, HLA-DRB1*12, DRB1*15, and DRB1*07 showed significant associations with HPV-16-positive CaCx and persistent HPV infection (Table 3). The significant positive associations of HLA-DRB1*12 in the present study corroborated with Japanese (53) and French (54) women with HPV-16-positive CaCx. HLA-DRB1*15 showed a susceptibility risk to invasive CaCx in the north Indian population (28) that corroborates with our present study. Significant associations of HLA-class II alleles with respect to HPV-16-positive CaCx in the present study correlated with our meta-analyses data (5). HLA-DRB1*07 was identified as a predisposing factor for persistent infection in the present study and in the American (37) and Dutch (7) populations. We also identified HLA-class I and II haplotypes to be associated with HPV-16-positive CaCx and persistent HPV-16 infection in the present study (Table 4). The 2-loci haplotype HLA A*02-B*27 has shown a significant association with persistent HPV-16 infection in the present study. The same haplotype was typical of patients with progressive keratoconus disease (24). The class II HLA haplotype (DRB1*04-DQB1*03) showed a negative association with both the groups (HPV-16-positive CaCx and persistent HPV-16 infection) of the present study, whereas the same haplotype exhibited susceptibility to cervical precancerous lesions in north Indian women (28). This discrepancy could be due to variation in allele frequency between north and south India.
The high-risk HPV type-16 is a pandemic pathogen and poses a major threat by causing almost 60% of cervical infections in women worldwide (34). There are five different clades of HPV-16 being phylogenetically classified as A1–A3, A4, B, C, and D and they exhibit distinct geographic distribution: European lineage “A,” which is predominant in Europe, Asia, and South America (A1–A3); East Asian lineage A4; non-European lineages “B, C, and D,” which are more heterogeneous in distribution. The lineages B and C are predominant in Africa, and D is predominant in North and South America (11,42). Clade-specific variation in the pathogenesis of HPV-16 has been reported (70). Unpublished data from our laboratory showed that the HPV-16 isolates in the study samples belonged to the lineages A and D. The diversity of HPV-16 thus may be a contributing factor for the observation of multiple HLA alleles being associated with the infection. Furthermore, it has been shown that HPV-16 infected the ancestral human populations and the viral phylogeny exhibits splits as that of Neanderthal/Denisovan and ancestral modern human populations suggesting that the viral lineages codiverged in parallel with the hosts with the split between archaic (42). This coupled with the global diversity of HLA alleles suggests a dynamic and a continuous process of host/pathogen genome coevolution.
In the present study, we performed low-resolution HLA typing using the PCR-SSP method for the HLA class I and II alleles, which pertains information to the first 2 digits of HLA alleles. We believe that our study is the first of its kind to identify the association of HPV-16 infection and HPV-16-positive CaCx with HLA alleles in women from the south Indian state of Tamil Nadu. It has been previously established that Tamil Nadu populations are genetically very diverse and ancient (2). The diversity of HLA alleles in this region is one of the highest in the world (43). Thus, the identification of HLA allelic associations in this population may still be informative even at this resolution. Performing high-resolution typing or sequence-based HLA typing may provide further insights on the observed associations of the present study. In addition, the discrepancy between the present and previous studies reported in the same population can also be attributed to the differences in the resolution level of genotyping method adopted in each study. We used a total of 71 cases (50 HPV-16-positive cases and 21 women with persistent HPV-16 infection) and 74 controls for the present study. Considering the possibility of type II error, we calculated the statistical power for the present sample size and we achieved a statistical power of about 84%. However, for detecting a single allele of statistical significant association, the present sample size may not be sufficiently powerful.
It is to be noted that the present study used a time frame of 4 months to define persistent HPV infection. Previous studies have used time frames ranging from 3 to 24 months to define persistence. Byun et al., Hoffman et al., Kudoh et al., and Dutta et al. had a category of the cohort that has persistent infection for 3 months (12,20,26,32). On the contrary, Liu et al., So et al., Radley et al., Skinner et al., Zhu et al., Ramanakumar et al., Konno et al., and La Torre et al. had used 6 and 12 months as separate time points for determining persistence (29,33,35,48,50,57,58,72). However, none of these studies correlated persistence with HLA genotypes. Thus, the present study, which has used a 4-month period, may be only an indicator to persistent HPV-16 infection. Further studies with a longer follow-up period may be essential for confirmation.
To summarize, the association of HLA class I and II alleles with HPV-16-mediated CaCx and persistent HPV-16 infection was studied in the Tamil Nadu population. Among the HLA-class I alleles, A*11 and B*57 showed a positive association with HPV-16-positive CaCx, while B*44 showed a significantly positive association with persistent HPV-16 infection. HLA-B*27 allele showed the highest association with both HPV-16-positive CaCx and persistent HPV-16 infection. A significant negative association was observed in HLA-B*15 with HPV-16-positive CaCx. Similarly, HLA-DRB1*12 exhibited a very significant association with both HPV-16-positive CaCx and persistent HPV-16 infection, while DRB1*15 showed a significant association with HPV-16-positive CaCx, and DRB1*07 was significantly associated with persistent HPV-16 infection. Future studies in different populations with a larger sample size, high-resolution HLA typing, and comparison with HPV-16 genetic diversity shall further provide insights on the exact role of HLA alleles in the development of HPV-16-mediated CaCx.
Footnotes
Acknowledgments
We thank all the volunteers who participated in the present study.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
We thank DST-SERB for providing financial assistance to perform this work and project assistant fellowship for the first author of this article under grant order YSS/2014/000136. We thank SASTRA Deemed University for providing reagents under the R&M fund. We also thank CSIR for providing Senior Research Fellowship for the first author of this article under the fellowship order 09/1095(0042)/19-EMR-I.
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.