
Abstract
Yellow fever virus (YFV) is the prototype member of the genus Flavivirus, which contains more than 60 positive-sense, single-stranded RNA viruses, many of which are considered public health threats. YF disease is controlled by a live attenuated vaccine, 17D, which was generated empirically through serial passage of the wild-type (WT) strain Asibi in chicken tissue. The vaccine, which has been used for over 80 years, is considered to be one of the safest and most effective live attenuated vaccines. It has been shown that the humoral immune response is essential to a positive disease outcome during infection. As such, the neutralizing antibody response and its correlation to long-term protection are a critical measure of 17D efficacy. The primary target of these antibodies is the envelope (E) protein, which is the major component of the virion. Monoclonal antibodies can distinguish WT strain Asibi and vaccine strain 17D by many different measures, including physical binding, hemagglutination inhibition, neutralization, and passive protection. This makes the WT-vaccine pair ideal candidates to study the structure–function relationship of the E protein in the attenuation and immunogenicity of flaviviruses. In this study, we provide an overview of structure–function of YFV E protein and its involvement in protective immunity.
Flaviviruses
The genus Flavivirus contains many medically important arthropod-borne viruses (arboviruses) such as dengue virus (DENV), Japanese encephalitis virus (JEV), St. Louis encephalitis virus (SLEV), tick-borne encephalitis virus (TBEV), West Nile virus (WNV), yellow fever virus (YFV), and Zika virus (ZIKV). Flavivirus infection can be neurotropic (JEV, SLE, TBE, and WNV), causing encephalitis, meningitis, and long-term sequelae in survivors, viscerotropic, (YFV) leading to jaundice, renal failure and hemorrhagic disease, or teratogenic (ZIKV).
The Flavivirus genome consists of three structural genes (capsid [C], premembrane/membrane [prM/M], and E) that make up the virion and seven nonstructural genes (NS1–NS5) that constitute the viral replication complex. Both structural (25,29,32) and nonstructural (15,30,51,65) genes are targets for antibodies; however, neutralizing antibodies (nAbs) are directed exclusively against the prM and E proteins (5,14,56). The prM protein elicits a weak nAb response, whereas the E protein elicits strong nAb responses. In addition, the E protein has been shown to elicit antibodies with other functional properties, such as hemagglutination inhibition (HAI), hemolysis, and passive protection of mice (16,17,24).
Yellow Fever Virus
YFV, the prototype member of the genus, is endemic in sub-Saharan Africa and tropical South America. YF disease ranges from asymptomatic to severe jaundice and hemorrhagic fever. As no antiviral therapies exist, the primary disease control strategy is vaccination with the live attenuated vaccine, strain 17D. Despite the availability of a safe and effective vaccine, YFV still causes large, periodic outbreaks. 17D was derived through serial passage of the wild-type (WT) strain Asibi in chicken tissue (70). Over eight decades of use, 17D has proved to be an excellent vaccine that is highly immunogenic. Currently, three substrains of 17D are used as vaccines (17D-204, 17DD, and 17D-213) (12). As with JE and TBE vaccines, studies have shown that nAbs are the correlate of protection for the YF 17D vaccine (21,42,47). Indeed, a log neutralization index of 0.7 [i.e., neutralization titer determined by constant antibody and varying concentrations of virus (47); and a 50% plaque reduction neutralization titer (PRNT50) of around 1 in 40 (42)] (i.e., constant virus and varying dilutions of antibody) is considered to be protective; however, these assays have not been compared head to head in a validated assay.
Although the mechanism of 17D attenuation is unknown, the amino acid residues that distinguish Asibi and 17D viruses have been determined (33,63). Overall, the three 17D vaccine substrains differ from WT Asibi virus at 20 common residues, of which, 9 are in the structural proteins (Table 1) and 11 are in the nonstructural proteins. The E protein contains the most amino acid substitutions (n = 8), and is responsible for tissue tropism and receptor binding, and the major target for nAbs [reviewed in ref. (50)], suggesting that this protein plays a key role in attenuation and immunogenicity. To understand structure–function of the YFV E protein, much work has been undertaken comparing the YFV vaccine strain 17D and its WT parent, Asibi. The role of Asibi/17D residues in the E protein on neuroinvasiveness in mouse models has been well documented (4,20,53,60), but there is relatively limited information on viscerotropism.
Amino Acid Changes Between Wild-Type Parental and Vaccine Strains of Yellow Fever Virus
The WT strains of YFV, Asibi and FVV, which were used to generate the two YFV vaccine strains substrains and FNV, were isolated in the same outbreak and are therefore genetically very similar. 17D and FNV were generated through independent serial passage series with 17D being passaged primarily in chick embryo lacking neuronal tissue, whereas FNV was passaged primarily in mouse brain. Adapting WT YFV to these distinct tissues led to changes in the E protein, which can be detected by mAbs.
mAb, monoclonal antibody; YFV, yellow fever virus; WT, wild type; FVV, French viscerotropic virus; FNV, French neurotropic vaccine.
In addition to 17D, another live attenuated YFV vaccine, the French neurotropic vaccine (FNV), derived by 126 passages of WT strain French viscerotropic virus (FVV) in mouse brain, was used to combat YF until 1982 when production stopped due to unacceptable levels of postvaccinal encephalitis in children (7). WT strains Asibi and FVV were isolated from the same outbreak and only differ by two amino acids (Table 1) (13). Interestingly, despite being derived by passage in chicken and mouse tissue, respectively, 17D and FNV acquired two common amino acid substitutions during the attenuation process: M-36 and NS4B-95 (39).
Structure of YFV E Protein
During infection, the E protein binds to cell receptors (that are currently not known for YFV) and the virus is internalized by receptor-mediated endocytosis. The virus is then trafficked through the cytoplasm inside endosomes. Interestingly, 17D-204 virus has been shown to enter cells in a unique, clathrin-independent mechanism, whereas WT Asibi virus utilizes the classical, clathrin-mediated endocytosis pathway (26).
Initial structural studies of the E protein were determined by physical epitope mapping the surface of the E protein with monoclonal antibodies (mAbs), together with functional studies on the mAbs [reviewed in ref. (58)].The antigenic structure of the Flavivirus E protein was first determined by X-ray crystallography of the TBEV E protein as three domains, termed C, A, and B, which were subsequently found to map onto domains I, II, and III (EDI, EDII, and EDIII), respectively (57). The YFV E protein is 493 amino acids in length with the 400 N-terminal amino acids containing the ectodomain (EDI, EDII, and EDIII). EDI and EDII are discontinuous in sequence, while EDIII is continuous (Fig. 1A). EDI contains a nine-stranded β-barrel and is a linker between EDII and EDIII. It connects to EDII by four flexible linkers and EDIII by one. These linkages are the hinges that allow for conformational change to occur during the replication cycle (Fig. 1B, C) (48). EDII, an elongated finger domain, contains the fusion loop that interacts with target cell membranes during attachment and fusion (Fig. 1C) (2,49). EDIII is an immunoglobulin-like domain that is thought to be involved in receptor binding, but has yet to be conclusively proven (57). Amino acid substitutions in EDIII are associated with Flavivirus pathogenicity (19,35,37,41). The C-terminal 100 amino acids contain the stem-anchor region that connects the two transmembrane domains that anchor the E protein in the viral membrane and is necessary for the rearrangement of E on the surface of the virion through its interactions with prM (3,73).
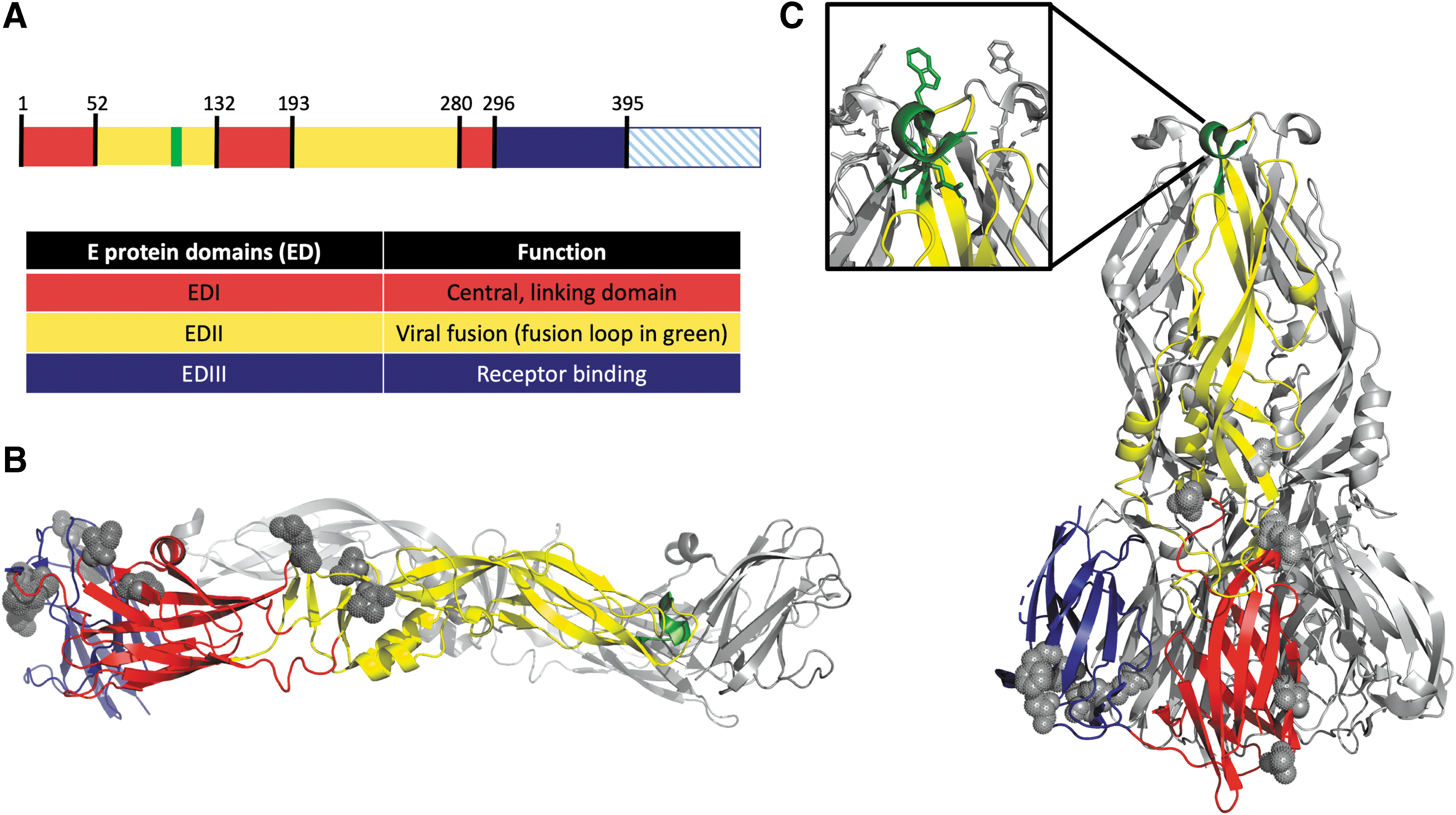
The YFV E protein is made up of three domains. Discontinuous domains I and II and continuous domain III have been shown to have important functions in dimerization and the conformational change that allows the virus to enter cells
Flaviviruses have extensive serologic cross-reactivity due to cross-reactive epitopes on EDI and EDII. In particular, the fusion loop contains a sequence of amino acids that are conserved between all flaviviruses and epitopes that elicit reasonable, but not strong, neutralizing activity. EDIII is ∼100 amino acids in length, has no glycosylation sites and one disulfide bridge, but contains epitopes that elicit high titer neutralization activity in vitro. As such, it has been an attractive vaccine immunogen because it can be easily expressed and employed on a variety of vaccine platforms. However, animal studies have shown it to be a poor immunogen in vivo, eliciting weak nAb titers (23,52). Recent studies have shown that EDIII epitopes are rarely seen by the immune system (i.e., few B cells produce antibodies to EDIII) and that most epitopes seen by the human immune response are conformational, including those that cross domains (59).
Using nuclear magnetic resonance (NMR) spectroscopy, the structure of Asibi EDIII was reported and shown to be slightly different to other flaviviruses due to the addition of a proline at E-325 that causes the BC loop of the YFV EDIII to be smaller than most mosquito-borne flaviviruses (71). Based on mouse mAbs, the EDIII BC loop contains major neutralization epitopes of JEV (E-333), WNV (E-332), and YFV 17D-204 (E-305, E-325), suggesting that this region is important antigenically (11,55,60,74). Despite changes to the BC loop, it was recently shown that the overall structure of the YFV 17D-204 vaccine virus E protein is very similar to other flaviviruses in both prefusion and postfusion states (46).The structure of the prefusion E protein state was determined by x-ray crystallography at 2.4 Å and postfusion at 3.1 Å based on 17D-204 E protein (Genbank: X03700) that was expressed in Escherichia coli BL21. Consistent with NMR studies, analysis of domain orientation showed that YFV EDIII was the most divergent from other flaviviruses with an increased rotation of EDIII during the dimer to trimer transition, bringing the C terminus of EDIII and the fusion loop 28 Å closer together (46). There is currently no high resolution cryogenic electron microscopy structure for YF virions.
E Protein Epitopes Recognized by mAbs
mAbs have been generated to many flaviviruses using live virus, inactivated virus, and recombinant E protein immunization of mice, humans, and chimpanzees [reviewed in ref. (38)]. In contrast to other flaviviruses, panels of YFV mAbs have been prepared using both live attenuated vaccines and WT viruses.
Specifically, YFV mAbs have been prepared from mice (1,8,10,16,21,43,54,66,67) and humans immunized with 17D-204 vaccine (21), mice immunized with FNV (28,30,54), and mice immunized with WT strain Asibi (31), enabling differentiation of vaccine- and WT-specific epitopes on the E protein. There are only three human mAbs and all are YF species specific (21); the remainder is derived from mice. As with other flaviviruses, nAbs are directed toward all three E protein domains. mAbs have shown that EDI and EDII contain many conformational Flavivirus cross-reactive epitopes that, in general, either elicit low levels or no neutralizing activity. In contrast, most epitopes on EDIII are linear, either strain or YFV species specific or Flavivirus serocomplex specific, and elicit high titer neutralization in vitro (16,30,64,61,44,66). Interestingly, the mAbs with the highest neutralization activity were directed against EDIII and recognized 17D-204 substrain-specific epitopes (16,21,66). Somewhat surprising has been the observation that YFV species-specific epitopes (i.e., only recognize YFV strains) elicit low levels of nAbs (21,31,45,60,61,66,67) and map to conformational epitopes on EDI and EDII (21,45,61) (Fig. 2).
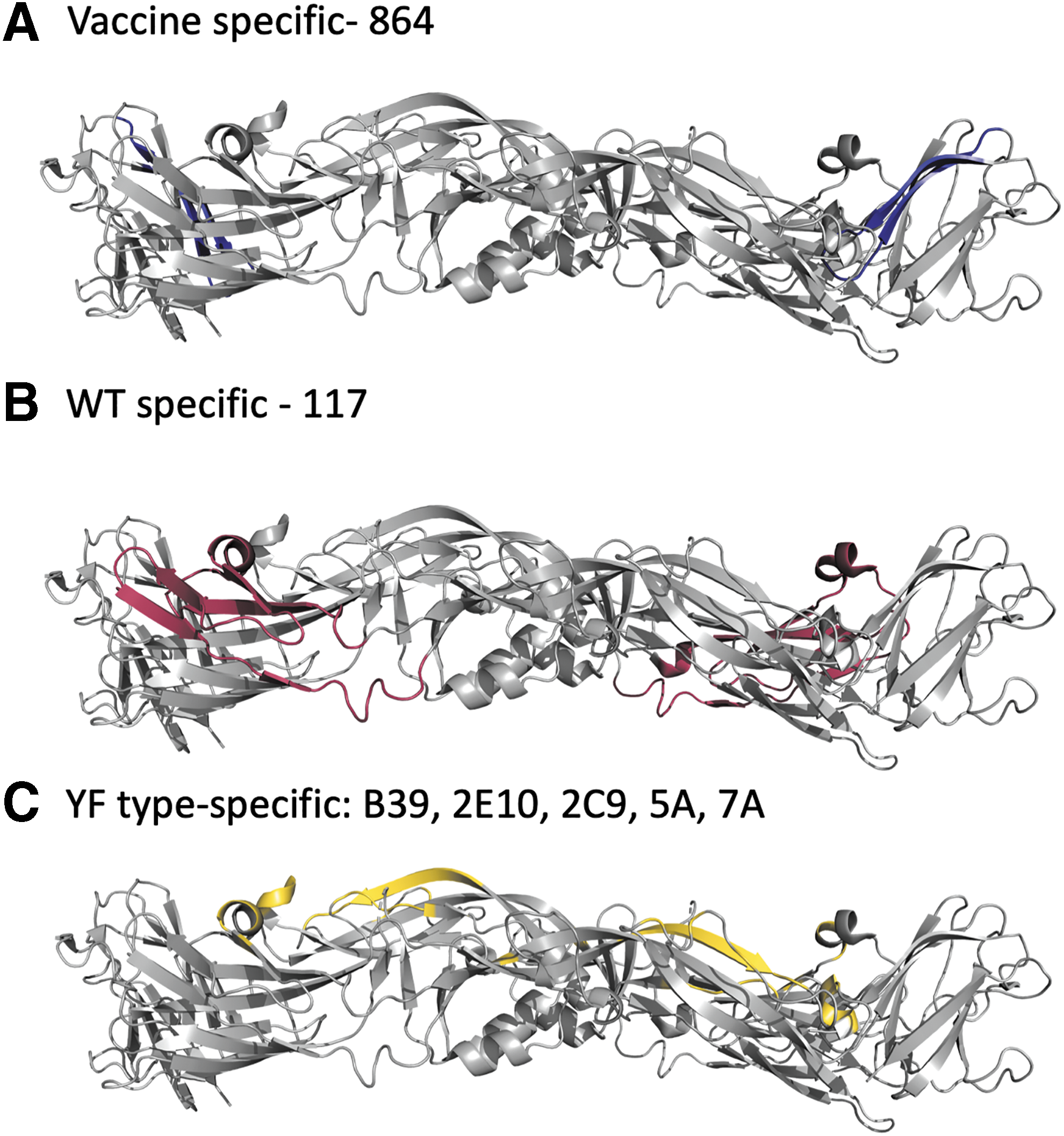
mAb specificity is mapped to distinct regions of the E protein. The 17D-204 vaccine-specific epitope of mAb 864 has been mapped to a surface-exposed region of EDIII, which is shown in blue
The mechanism of neutralization of YFV by antibodies has been poorly studied to date. Neutralization of YFV was first studied by Buckley and Gould (16) who showed that the YFV E protein was primarily responsible for eliciting nAb responses. Moreover, as with other flaviviruses, comparisons of neutralization indices and physical binding titers showed that the presence of an epitope alone was not enough to confer neutralization or provide protection (16,29). Recently, Lu et al. have used structural biology to investigate the mechanism of neutralization by a human mAb, 5A (21,46). X-ray crystallography of prefusion and postfusion forms of the E protein and mAb attachment shows that at the prefusion step, the mAb binds to highly conserved regions of the YFV EDII, while in the postfusion from, it prevents the fusion loop interaction with the endosomal membrane (46).
mAbs Used to Map Determinants of Virulence and Attenuation of YFV
Over the last four decades, a number of groups have generated mAbs against YFV, the majority of which are specific to the E protein (1,10,17,30,64,68). By comparing neutralizing capabilities, physical binding of non-neutralizing mAbs, HAI titer, and passive protection, a structure–function relationship has been elucidated for E protein epitopes. In the mid-1980’s, five antigenic domains were described on the surface of the E protein: 17D specific, vaccine (17D and FNV) specific, YFV (WT and vaccine strains) specific, intermediate reactivity with flaviviruses, and broad reactivity with flaviviruses (Table 2) (17,67).
Properties of Yellow Fever Virus-Derived Monoclonal Antibodies
The development of YFV mAbs allows strains of YFV to be distinguished by both structural and biological properties.
HAI, hemagglutination inhibition.
17D-specific mAbs can be further delineated into mAbs that recognize all substrains of 17D (i.e., 17D-204, 17D-213, and 17DD) (mAbs 861 and H6), 17D-204 substrain specific (mAbs 8A3 and 864) and 17DD substrain specific (mAb H10), which suggests that even though the substrains of 17D are biologically very similar, structurally there is some variation (30,64,68). This is supported by nucleotide sequence data that show E protein amino acid differences between substrains at residues 56, 153, and 407 (Table 1) (40). mAb 864 is a 17D-204 substrain-specific mAb whose epitope is functionally important to the virus based on very high neutralization titers (16,30). mAb 864 neutralization-resistant mutants identified spatially adjacent EDIII residues E-305 and E-325 as part of the epitope important to mAb recognition and E-305 as a YFV mouse neurovirulence determinant (Fig. 2A; Table 3) (60,62). mAb 8A3 has a similar specificity to mAb 864 and elicits high neutralization activity; however, no mAb 8A3 neutralization-resistant mutants have been characterized (66,67). Only mAb 429 has been identified as FNV specific and that recognized NS1, not the E protein (30).
Mapping of Yellow Fever Virus Monoclonal Antibodies to the E Protein
Vaccine-specific mAbs (mAbs 411 and H5) recognize all 17D substrains and FNV (30,68). mAb 411 has no recognized biological activity, whereas mAb H5 has HAI activity (16,68). In addition, mAbs have identified epitopes that exclusively recognize WT strains (mAbs 117, S17, S18, S24, and S56). Two of these mAbs, S17 and S56, had no neutralizing activity, whereas the remaining WT mAbs generated, 117, S18, and S24, had neutralizing activity. WT-specific epitope mAb 117 was mapped to E-173 using escape variants (Fig. 2B; Table 3) (62). Interestingly, neither WT-specific (mAbs 117 and S17) nor vaccine-specific epitopes (mAbs 411 and H5) identified elicit neutralizing activity, suggesting that they may contribute to the structure of the E protein (16,18,66,68). In addition to the two live attenuated vaccine strains, there is an attenuated variant of Asibi virus generated by six passages in HeLa cells (Asibi Hp6) that is characterized by loss of viscerotropism, loss of neurotropism, and loss of mosquito competence (the same phenotype as 17D vaccine) (16,18,66,68). Significantly, Asibi Hp6 virus gained the vaccine epitopes recognized by mAbs 411 and H5 (9,34). Coincidental with the gain of vaccine epitopes were the loss of WT epitopes recognized by WT mAbs 117, S17, S18, S24, and S56, suggesting vaccine epitopes are structurally involved in the attenuated phenotype.
Functional Mapping of mAbs Against YFV E Protein
A large panel of YFV species-specific antibodies (mAbs 3A3, 2C9, 2D12, 2E8, 2E10, 3E9, 4E1, 5E3, 429, 492, 825, 863, 871, 979, B26, B39, 3B6, 3F4, 4A1, 4C9 4H10, 5H2, 5B6, 8H3, 5A, R3, and 7A) has been identified as recognizing WT and vaccine strains of YFV (1,10,21,18,30,66). mAbs 2C9, 2E8, 2E10, 825 5A, and 7A and R3 neutralize virus and are active by HAI against both 17D and Asibi viruses. mAbs 2B8, 2D12, 3A3, and 4E1 neutralize Asibi virus only. mAb 5E3 only neutralizes 17D virus, but is active by HAI against both WT and vaccine strains of YFV (Table 2).
A YF type-specific mAb, B39, was mapped by neutralization escape mutants to EDI (E-155 and E-158 mutations in distinct escape mutants) (30). Interestingly, mAb B39 is only able to neutralize 17D-204 and not WT strains (61). 17D-204 viruses resistant to B39 showed no change in virulence or neurovirulence in a mouse model, suggesting that the B39 epitope does not contribute to YFV virulence. The YF type-specific mAb 2E10 (epitope located in EDII: E-71) is able to neutralize B39-resistant 17D-204 but not 17D-204 itself, suggesting that 17D-204 has multiple, topographically distinct YFV-specific epitopes or the epitopes that are part of a bigger antigenic site (27,36,61). mAb 2E10 is also active by HAI and elicited passive protection in a mouse model (45,61). Regions associated with YF type-specific mAb alter the hydrophobicity of the E protein, a property seen in a majority of mAb neutralization-resistant Flavivirus mutants (Fig. 2C; Table 3).
YFV E protein also contains intermediate Flavivirus reactive epitopes (mAbs 4E11, 5H3, 5E5, 5E6, 126, 140, 423, 427, 818, 819, 829, 869, 917, 924, 925. 992, 993, and 999) that recognize one or two flaviviruses, in addition to YFV (1,10,17,21,64). Some of these intermediate Flavivirus reactive mAbs could be broadly reactive Flavivirus mAbs; however, they have only been tested against one or two flaviviruses. Nonetheless, a number of Flavivirus broadly reactive epitopes (mAbs 110, 427, 612, 813, 843, and 868) have been identified on the YFV E protein. Significantly, mAb 813, raised against 17D-204 virus, recognizes all flaviviruses tested (>50) and has weak neutralizing and HAI activity (17,64).
Not All mAbs Elicit Passive Protection
The ability of YFV mAb to passively protect against infection has been tested for a handful of those previously mentioned. Mouse mAbs 2B8, 2C9, 2E10, 4E11, 5E3, 5H3, 813, 825, 864, and 868 and human mAb 5A provide protection against 17D-204 challenge in a mouse model (45,64). Two mAbs in this group, 2C9 (E-71, E-125) and 5A (E-65-74, E-81, E-84, E87, E101-E104, E240-244), have been mapped to EDII (Fig. 2C; Table 3). Two mAbs, 411 and H10, provided passive protection against the 17DD substrain of 17D exclusively (21,30,66). Other mAbs were tested, but did not provide passive protection against 17D-204 challenge, including mAbs 8A3, 110, 126, 140, 427, 612, and 843 (30,68). Although the majority of YFV mAbs have not been evaluated for passive protection against WT Asibi virus, to date, only mAb 117 protects mice against intracranial WT Asibi virus challenge (30,66).
Antibody-Dependent Enhancement
The phenomenon of antibody-dependent enhancement (ADE) of virus infection has been investigated for most flaviviruses [reviewed in ref. (50)]. While ADE during YF infection can be shown in cell culture (64) and mice (6) with antisera from 17D vaccinees and YFV mAbs, there is no consistent relationship between the pattern or degree of neutralization/HAI and YFV ADE (22), and no evidence that ADE takes place in humans. Indeed, over 850 million doses of 17D vaccine have been distributed with no reports of ADE. Accordingly, there are few studies on ADE with YFV.
Conclusions
Understanding the biological mechanisms behind mAb specificity and potency is extremely important in the context of next-generation Flavivirus vaccines. YFV is the only example of a Flavivirus where mAbs have been raised against both vaccine and WT strains. This provides an excellent model to study the immunogenicity and structural role of the E protein in live Flavivirus vaccines. Furthermore, it will be an important goal of any second-generation vaccine design to increase the durability of the vaccine Ab response toward accessible and conserved structures of the E protein to ensure the optimum opportunity for lifelong protective immunity.
An important consideration when undertaking future vaccine design projects are the approaches for which mAbs have been historically generated, especially YFV where most mAbs were generated over 20 years ago. Murine humoral immunity to YFV is typically directed at EDIII, whereas studies of the human response to YFV suggest that human mAbs are most often directed at EDII (1). It is worth noting that recombinant EDIII is often used as an immunogen to generate anti-Flavivirus mouse mAbs, which may skew the immune response toward an artificial immunogen. In some studies, the response to YFV prM was higher compared with EDIII, further underscoring the importance of virion maturation in the Ab response to natural infection and vaccination (21,44,72). The structural basis for this mAb direction is unclear as the fusion loop is located in a cryptic site of mature virions. Murine immune responses can be pushed toward EDII and the fusion loop if virus is treated with detergent, exposing these buried regions (72). Studies on the human mAb response also showed that there was minimal reactivity to monomeric Flavivirus E protein, but extensive reaction to overlapping, dimeric or herringbone epitopes (69). This suggests that the model of E protein maturation does not fully capture the complexity of E protein rearrangement or that fusion loop epitope is larger and more complicated than previously thought. Nonetheless, YFV mAbs generated to date are a useful tool in mapping the structural changes to the virion that distinguish WT and vaccine strains of YFV.
Studies comparing WT to vaccine virus with E protein mAbs show that the E proteins are topographically and functionally distinct, but differences tend to be largely subtle. To add to this complexity, very few mAbs generated against YFV elicit high titer neutralization. More puzzling is that the highest nAb titers come from vaccine-specific mAbs, which are unable to neutralize WT YFV. As such, it is unclear how the YF vaccine 17D E protein elicits nAbs that provide protective immunity against WT YFV infection if the major neutralization epitope(s) only recognize(s) vaccine epitopes and YF species-specific epitopes are not associated with high neutralization activity; however, there are two possible explanations. First, it has been shown that 17D vaccine virus gives higher neutralization titers to homologous 17D virus than WT strains (16,21), which is consistent with the experimental data described above. Second, the mAbs described above were made at least 15 years ago and recent studies with human mAbs raised against DENVs have shown the human nAb response is to conformational epitopes that cross domains (1). Thus, our knowledge of the YF nAb response is likely incomplete.
Overall, mAbs have been important in understanding the structure–function of both YFV WT and vaccine E proteins, but our knowledge is still lacking in terms of understanding how the live vaccine elicits long-term protective immunity.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
A.D.T.B. is supported by a grant from the Gillson Longenbaugh Foundation. E.H.D. is supported by a predoctoral fellowship from the University of Texas Medical Branch Sealy Institute for Vaccine Sciences