
Abstract
Understanding of host pathogen interactions is important in planning strategies for effective control of the pathogen. The present study investigated the regulation of genes involved in the activation of splenic immune system in mature laying chickens challenged with T strain of infectious bronchitis virus (IBV). Among all the genes studied, the relative expression levels of Fas cell surface death receptor (FAS), interleukin 7 (IL7), IL18, proteasome subunit alpha 3 (PSMA3), major histocompatibility complex, class II (MHCII), interferon alpha (IFNα), immunoglobulin A (IgA), and nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) were significantly (p < 0.05) upregulated, while Toll-like receptor 7 (TLR7) and TLR5 were significantly downregulated in the challenge compared with the control group. Genes such as vascular cell adhesion molecule 1 (VCAM1), FK506-binding protein 1B (FKBP1B), transforming growth factor-beta 3 (TGFB3), NLR family pyrin domain containing 3 (NLRP3), TYRO3 protein tyrosine kinase (TYRO3), TNF receptor-associated factor 3 (TRAF3), C-X-C motif chemokine receptor 4 (CXCR4), macrophage inflammatory protein-3 (MIP3A), TLR2-1, TLR3, and TLR21 were not altered in mRNA expression levels between the challenge and control groups. In conclusion, the splenic immune response to IBV infection involved the regulation of cytokines, TLRs and NF-κB.
Introduction
Infectious bronchitis virus (IBV) is a gammacoronavirus with enveloped positive-sense single-stranded RNA of 27.6 kbp (6). IBV is the causative agent of a highly contagious infectious bronchitis (IB) in layers and broilers. After initial multiplication in the upper respiratory tract, the IBV quickly spreads to various organs, including the spleen (17). The most common symptoms of IB are related to the respiratory system and include nasal discharge, sneezing, gasping, and coughing (17). In layers, IB causes egg quality problems while, in broilers, in addition to the development of secondary infection, IB affects production traits. In Australia, layer chicks are routinely vaccinated for IB with one or both of A3 and Vic S vaccine strains. Both vaccinated and unvaccinated laying hens can shed IBV in feces up to 3 weeks postinfection (8).
Understanding the mechanisms of host immune system regulation in IBV infection is pivotal for IB control. The innate immune system is activated after the detection of a pathogen. The immune system and mucosal epithelial surface cells are involved in innate immune responses. The first step in innate immunity is the detection of the invading pathogen. The production of interferon (IFN) and other antimicrobial molecules is initiated once the host cells pathogen recognition receptors (PRRs) recognize pathogen-associated molecular patterns (PAMPs) (2,39). A subset of host cell PRRs can recognize viral nucleic acids exposed in the host cytoplasm. Toll-like receptors (TLRs) bounded to membrane can recognize endosomal nucleic acids and the family of RIG-I-like receptors recognizes viral RNAs (42). The primary ligands for the activation of TLRs are double stranded RNA and 5′-triphosphate containing RNA, normally absent from healthy host cells (21).
In IBV infection, an innate immune response activates when the virions bind to sialic acid receptors on the mucosal linings of the tracheal tissue (34). However, viral RNA is also recognized by PRRs (reviewed in Ref. 2). This immune response may be due to pathways in which TLRs are activated (13). Among TLRs, TLR7 and TLR3 are known for recognizing RNA virus-encoded PAMPs (1). As a result of virus multiplication, macrophage numbers are increased and natural killer cells are rapidly activated (18). T lymphocytes are actively involved in the clearance of IBV (13). To alter the host cell machinery, IBV multiplication in macrophages inhibits nitric oxide synthesis with or without affecting production of type I IFNs (22). The IFNs secreted from the infected cells in turn provide a signal for the infected and uninfected cells to induce the transcription of antiviral effector genes (IFN inducible genes) (21).
The production of IBV-induced IFNs depends upon host cell type and virus strain (16). The primary objective of the current study was to gain insights into the regulation of the splenic immune system in mature laying chickens challenged with IBV T (N1/62) strain. The outcome of the study further broadens our understanding of the regulation of the immune system-related genes by IBV infection in the spleen of adult birds.
Materials and Methods
Rearing of laying chickens and tissue collection
This study was a part of a project, in which IBV was used to understand its effects on chicken oviduct and spleen; therefore, only one time-point was selected for sampling. IBV T was selected as it is the main wild-type strain causing IB in Australia. In brief, day-old chicks vaccinated for Marek's disease were raised in isolation sheds with strict biosecurity measures. The rearing conditions were as per the standard of ISA General Management Guide 2009-10, Australia. At the age of 18-week, pullets were moved to individual cages in an isolated poultry house for monitoring individual birds. Sampling details of the chickens have been explained by Khan et al. (19).
RNA extraction from spleen tissue
RNA was extracted from an ∼100 mg of spleen tissue using TRIsure (Bioline, Australia), according to the manufacturer's instructions. The extracted RNA was purified using an RNeasy Mini Kit (Qiagen, Australia) as per the manufacturer's protocol. A DNase-I was used to remove traces of genomic DNA. The purified RNA was tested in an Agilent 2100 Bioanalyzer (Agilent Technologies, Germany) to measure its quantity and purity. All the RNA samples gave an average RNA integrity number ≥9.10.
Primer sequences and validation
The genes chosen in the current study are involved in various pathways of the host immune system. Primers for the candidate target genes were either sourced from published literature or designed (in NCBI and primer 3 software) by taking into account the exon-intron spanning (Table 1). Specific amplifications of the primers were confirmed in an Agilent 2100 Bioanalyzer gel electrophoresis using a DNA-1000 Kit. The primer amplification efficiency (%) was in a range of 90% to 105%.
Primer Detail of Candidate Target and Reference Genes
Relative gene expression analysis
Quantitative PCR on spleen RNA samples was performed as per the protocol of SensiFAST SYBR Lo-ROX One-Step RT-PCR Kit (Bioline) in Rotor Gene Q thermocycler. The two-step cycle conditions were as per protocol of the PCR master-mix kit with annealing and extension temperature according to Table 1 for 20 sec.
IBV load in spleen
Viral load in the spleen tissue was quantified using qPCR and a standard curve constructed from eight different dilutions (10-fold) of recombinant plasmid vector containing a 181 bp fragment of S1 glycoprotein gene of IBV. The individual RNA samples from spleen were converted into cDNA using the QuantiTect Reverse Transcription Kit (Qiagen). To quantify viral RNA (as cDNA copies) in the samples, qPCR was performed using the SensiFAST™ SYBR® No-ROX Kit (Bioline). Viral load per gram of spleen tissue was calculated as previously described (20).
Statistical analysis
Gene expression data were analyzed in qbase+ software version 3.0 (37) using YWHAZ and TBP as reference control genes (19). YWHAZ and TBP were used, as they were the most stable reference genes in the spleen of IBV-infected chickens (19). The normalized relative quantities from qbase+ were further analyzed in StatView software (SAS Institute, Inc., Version 5.0.1.0) by taking virus challenge as main effect. Level of significance between the treatment groups was separated by Tukey–Kramer test (p < 0.05).
Results and Discussion
Efficiency and specificity of primers
Efficiency (%) of individual primers was in the acceptable range (90% to 105%). All the primers were specific in amplification as confirmed by the Agilent 2100 Bioanalyzer gel (Fig. 1A, B). The melting curve analysis of all primer pairs showed a single peak indicating primer specificity for the target gene amplification.
Amplicon size of gene fragments amplified by PCR.
IBV load in spleen
The mean viral load of IBV T RNA (cDNA copy number) in the challenged birds sampled on days 9–10 p.i. was 2.28 × 107 copies per gram of spleen tissue. As described in our previous report, a significantly higher titer of IBV antibodies for the challenged birds (S/P value 1.443) compared with the control group (S/P value 0.122) in the ELISA test confirmed the modulation of the immune response by the virus (20). All the infected birds showed characteristic clinical signs of IB as described (17). Using qPCR, no viral RNA (or cDNA) was detected in the samples collected from the control birds. As the primary site of multiplication for IBV is the trachea, immediately after the entrance of the virus, viral antigen can be detected from day 1 p.i. in tracheal tissues (24). The virus then spreads into systemic organs, such as kidney, oviduct, and spleen. In the current study, the 2.28 × 107 viral copies per gram of spleen tissue on days 9–10 p.i. showed that the virus was still multiplying in the spleen.
IBV infection significantly affected expression of immunity related genes
Among all the immunity-related genes chosen, IBV T virus challenge significantly (p < 0.05) increased the expression of FAS, IL7, PSMA3, IL18, MHCII, NF-κB, IFNα, and IgA (Fig. 2A–H) while decreased the expression of TLR7 and TLR5, compared with the control (Fig. 2I–J). On the contrary, most of the immune system-related genes CXCR4, TYRO3, TRAF3, NLRP3, TGFB3, FKBP1B, VCAM1, TLR21, TLR3, MIP3A, and TLR2–1 were not significantly (p > 0.05) affected by the virus challenge (Table 2). IBV infection studies often use young birds whose immune system is still developing. The current study used mature laying hens to explore the host response to virus multiplication in the spleen. Generally, the infected host immune system detects and responds to viral RNA through various mechanisms in a cell-type-specific manner (31).
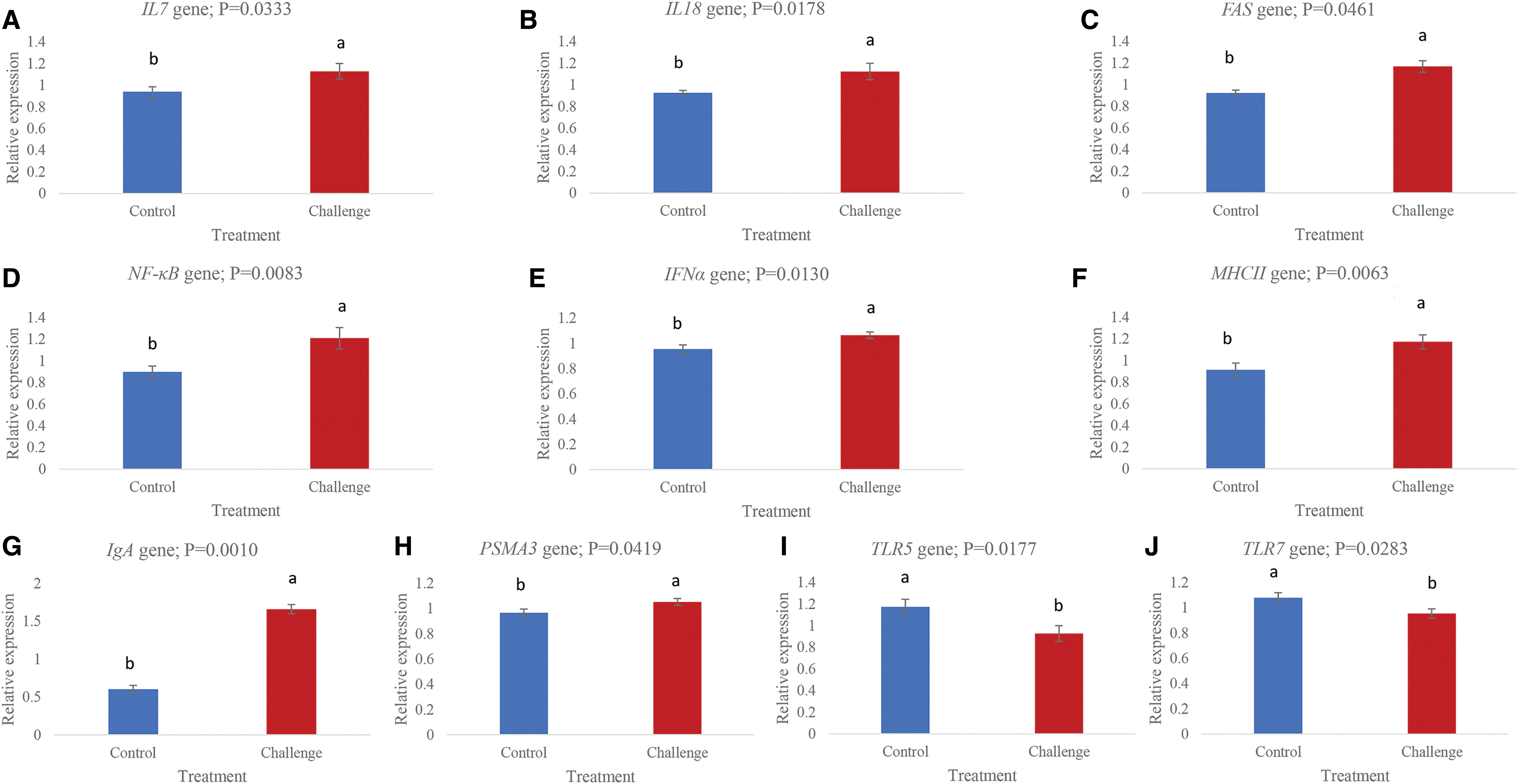
Relative expressions of genes in the spleen of control and infectious bronchitis virus challenged laying chickens.
Effect of Infectious Bronchitis Virus T Challenge on the Relative Expression Levels of Genes Involved in the Immune System Regulation
Values are mean of NRQs ± standard error. Relative quantities for individual genes are scaled to the average across all unknown samples per target gene. Cq values for individual genes were normalized in qbase+ using 2−ΔΔCq approach. Using one-way ANOVA, the NRQs were analyzed by taking virus challenge as an independent variable. There were 20 birds in each of the control and challenge groups.
NRQs, normalized relative quantities.
The data obtained in the current study have the limitations in terms of choosing one sampling time-point. In infectious study models, usually more than one sampling time-points are selected; however, we sampled the birds at days 9–10 p.i. for collecting oviduct samples to have a snapshot understanding of the virus effects on regulation of genes involved in egg and eggshell formations (19,20). Research on the immune system regulation in chickens against IBV has mainly focused on tracheal tissue in young chicks. Our findings showed that IBV T strain has the potential to regulate the spleen immune response in mature laying hens with a mean viral load of 2.28 × 107 (as cDNA copy number) per gram of tissue on days 9–10 p.i. This indicates that the infected chickens are regulating various immune system pathways in the spleen to clear the virus particles from the body. The main reason of choosing IBV T in the current study was that the virus is able to cause IB in Australian poultry industry in field conditions.
In the current study, the upregulated mRNA level of IFNα in the challenged compared with the control group indicates its role in IBV infection. A higher production of IFNα on days 9–10 p.i. indicates that the spleen was involved in pathways necessary for the clearance of the virus. A higher number of macrophages (3) and increased production of IFN (4) and IFNα mRNA level (32) in IBV challenge studies have been reported previously in chickens. The FAS gene encodes a protein called TNFRSF6/FASR/APO-1/CD95 or APT1. The FAS receptor is a death receptor on the surface of cells that leads to apoptosis induced by cytotoxic T cells (40). In the current study, a significantly higher expression of FAS in the challenge group suggests its role in apoptosis of the cells undergoing IBV multiplication. It seems that cells expressing FAS in the spleen were involved in pathways of apoptosis. The role of FAS in the apoptosis of cells undergoing IBV multiplication has been reported in chickens (9,14,15,38). Immunoglobulins play a vital role in the humoral response of host body to pathogens. Different classes of immunoglobulins respond differently to IBV infection in chickens, and their secretions depend upon tissue type, bird age, and course of infection (10,12,25,26,36). In the current study, a significantly higher mRNA level (p = 0.0010) of IgA in splenic cells undergoing IBV multiplication may explain its important role in humoral immunity. The upregulation of the IgA gene indicates that the IBV was eliciting its expression in splenic cells on days 9–10 p.i.
In the current study, both the TLR5 (cell surface TLR) and TLR7 (endosomal TLR) were downregulated by IBV challenge in the spleen of laying hens. The downregulations of TLR5 and TLR7 genes by IBV challenge may be the responses of the birds to combat viral multiplication in the splenic cells. Similarly, downregulation of TLR5 in the kidney of IBV-infected chickens (9) and downregulation of TLR7 in the spleen (14) and trachea (32) of chickens challenged with IBV have been reported. However, in the studies (32), the expression level of TLR7 significantly upregulated on day 5 p.i. It seems that the TLR5 and TLR7 were affected more in the splenic immune system by IBV compared with TLR2-1, TLR3, and TLR21, whose expression levels were not significantly different between the challenge and control groups. However, as the current study sampled the splenic tissue on days 9–10 p.i., a solid conclusion about the roles of TLR2-1, TLR3, and TLR21 in IBV infection cannot be drawn. TLR3 was significantly upregulated in tracheal tissue of IBV-infected chickens on days 1, 5, and 8 p.i. (32), which is contradictory with what we found in the current study.
In the current study, a significantly higher expression of NF-κB in the spleen of the challenged birds shows that the virus activated NF-κB pathway. The genes in NF-κB pathway might have then acted in immune response, cell survival, or cell proliferation in the spleen undergoing viral multiplication. Major histocompatibility complex II (MHCII) proteins play a pivotal role in the adaptive arm of the immune system through guiding the development and activation of CD4+ T helper cells (35). A significant upregulation of MHCII by IBV challenge indicates that the activation of T lymphocytes was enhanced. Interleukins (ILs) are types of cytokines involved in cell signaling. IL18 is a proinflammatory cytokine secreted mainly by macrophages and is involved in the induction of IFNγ and enhancement of NK cell activity. Significant upregulations of both IL7 and IL18, in the current study, show that IBV is a potent inducer of not only cytokines but also proinflammatory cytokines. It seems that both the IL7 and IL18 would have modulated a broad range of immune responses involving the proliferation and activation of macrophages, CD4+ helper T lymphocytes and regulation of transcription factors necessary for the production of other types of cytokines. A significant upregulation of proteasome subunit alpha 3 (PSMA3) by virus challenge indicates its role in triggering certain pathways (e.g., NF-κB, cytokine production) in the spleen of laying hens. In the current study, PSMA3, IL17, IL18, and NF-κB were all upregulated in the spleen by virus challenge. However, the upregulated PSMA3 does not coincide with the upregulation of cell adhesion molecules, as the expression level of vascular cell adhesion molecule 1 (VCAM1) was not significantly different between the challenge and control groups. VCAM1 seems to play an important role in the production of cell adhesion molecules (33) and is also involved in the activation of CD4+ T cells (11). However, VCAM1 was significantly upregulated in the spleen of chickens by IBV challenge reported in a previous study (14). Therefore, the VCAM1 expression in response to IBV challenge warrants further study. In conclusion, IBV T strain infection induced an immune response through the regulation of various genes in the spleen of mature laying hens that was observed on days 9–10 p.i. The significant regulation of genes showed that IBV affected the NF-κB pathway, cytokine production pathways, and the downregulation of TLRs. The possible role of the PSMA3 gene in the NF-κB pathway in the spleen immune response to IBV might be through proteasome complex formation. Further studies are required to understand the mechanisms of immune evasion strategies of IBV and downregulation of TLR5 and TLR7 in laying chickens.
Footnotes
Acknowledgments
Help from Dr. Sandra Sapats at the Australian Animal Health Laboratory, CSIRO, Geelong, Australia, for providing the IBV T (N1/62) strain is highly acknowledged.
Ethical Statement
The experimental procedures complied with the guidelines specified in the Australian Code for the Care and Use of Animals for Scientific Purposes 8th edition (2013). The experiment was approved by the Animal. Ethics Committee at the University of New England under authority no. AEC15-118.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
J.R. received funding from the Australian Egg Corporation Limited under grant no. AECL 1UN121. The funding body was not involved in experimental procedures or data analysis.