
Abstract
Chronic hepatitis C virus (HCV) infection leads to variable outcomes, ranging from prolonged slow hepatic damage leading to cirrhosis, and hepatocellular carcinoma (HCC). Polymorphism in cytokines IL-10 and IL-12 that impact the immune response to HCV infection may play a role in determining this outcome. This study was aimed to determine if polymorphisms in IL-10 and IL-12B contribute to HCV susceptibility and the risk of developing HCC in patients from Northeast India. IL-10 − 1082, −819, −592 polymorphisms and IL-12B −1188 polymorphisms were genotyped by polymerase chain reaction–restriction fragment length polymorphism in a total of 266 HCV-infected patients and 100 age- and sex-matched controls. In the HCV-infected subjects, 110 patients had chronic hepatitis C (CHC), 96 with liver cirrhosis, and 60 with HCC. Serum levels of IL-10 were also measured and correlated with disease severity. Haplotype analysis for IL-10 polymorphisms was carried out. Statistical data were analyzed using SPSS ver. 22.0. The frequency of IL-10 − 592 AA genotype/A allele was significantly higher in HCC patients than in CHC patients. The intermediate IL-10-producing ACC haplotype was significantly more frequent in HCC and cirrhotic patients than in CHC patients. No significant association was found for IL-10 − 819, −592 and IL-12B −1188 polymorphisms with the susceptibility to HCV infection or occurrence of HCC in HCV-infected patients. IL-10 − 592 CA polymorphism and IL-10 ACC haplotype are significant biomarkers of HCC in HCV-infected patients from Northeast India. Higher serum levels of IL-10 were also linked to higher disease severity.
Introduction
Hepatocellular carcinoma (HCC) is the most common primary liver carcinoma and around 80% of all liver cancers are classified as HCC (36). Overall, it is the fifth most common cancer and the second leading cause of cancer-related death worldwide (6,21). It is estimated that around on average 2.5% of patients who have chronic hepatitis C (CHC) develop HCC and the rate increases up to 7% after the establishment of cirrhosis in these patients (23,36). In India, 0.5–1.5% of the population is believed to be afflicted with hepatitis C virus (HCV)-induced chronic liver disease, with an increasing trend and is more frequent, particularly in areas such as Northeast India, Tribal populations, and in Punjab (2,35). The development of HCC in these individuals has been attributed to host factors (age, gender, ethnicity), viral (HCV genotype, viral load), and environmental factors (smoking, alcohol consumption) (25). Along with these factors, host genetics particularly those that affect immune response to viral infection are important considerations (48).
IL-10 is an anti-inflammatory interleukin produced mainly by monocytes, T cells, B cells, a small fraction of natural killer cells, macrophages, and dendritic cells (4). It is thought that IL-10 is directly involved in HCV-related liver disease progression, and the level of this cytokine has been found to be disease stage-specific. Serum levels of IL-10 are elevated in HCV-related cirrhosis and HCC (11,42,50). IL-10 levels are strongly regulated by IL-10 genetic variability by influencing their nuclear binding ability (15). The promoter region of the IL-10 gene contains three biallelic single nucleotide polymorphisms (SNPs) at positions −1082 (rs1800896), −819 (rs1800871), and −592 (rs1800872) bp from the transcription start site, which have been shown to have functional effects on the cytokine and also influence gene expression leading to differential IL-10 production (8,16).
The possible contribution of IL-10 promoter polymorphisms to HCV outcomes including HCV persistence and occurrence of HCC has been addressed in several studies with varied results. The IL-10 − 1082 GG and AG genotypes have been shown to be more common in patients who had persistent infection in multiple studies but were not found associated with HCC occurrence in a meta-analysis (3,46,47). Similarly, although IL-10 − 819 C allele and IL-10 − 592 A were observed higher in HCV patients compared with the control group, other studies have found no such associations (3,26). Variation in IL-10 serum levels among individuals has also been associated with three different haplotypes GCC, ACC, and ATA. The GCC haplotype, which is considered to be a high producing haplotype, has been found to be abundant in Western populations and rarer in Asian populations (42). Few studies have also reported GCC haplotype to be higher in HCV-infected patients than in controls (13). Conversely, the intermediate IL-10-producing ACC haplotype was reported in patients with HCC (42).
One of the key roles played by IL-12 is maintenance of the balance between T helper 1 (Th1) differentiation and T helper 2 (Th2) response from its precursor during viral infections (41). IL-12B is a proinflammatory cytokine, which triggers a Th1 response including IL-10 production (49). IL-12 production has been shown to be influenced by genetic variations in the gene. The C allele of the −1188 AC polymorphism in the 3′ untranslated region (UTR) of the IL-12B gene (rs3212227) is associated with enhanced IL-12B production (38). Homozygosity for the A allele at position −1188 of the IL-12 gene has been associated with HCV persistence in Chinese, Caucasian, and Egyptian HCV-infected patients (53). Furthermore, multiple studies have also reported IL-12B −1188 polymorphism to represent a significant risk factor in the occurrence of multiple cancer types including HCC (18,41).
These IL-10 and IL-12 SNPs are heavily influenced by ethnic and regional factors (1). Thus, the objective of our study was to validate the role of these polymorphisms and the resultant haplotypes as markers of HCV susceptibility and disease severity in patients with HCV infection from a Northeast Indian perspective.
Materials and Methods
Patients
Patients were recruited on the basis of anti-HCV antibody and HCV RNA positivity from the Outpatient Department of the Department of Gastroenterology, Gauhati Medical College and Hospital. Two hundred sixty-six HCV-infected patients were included in this study. Inclusion criteria of HCV-infected patients were the absence of concurrent HBV, HCV, or HIV infection, pregnancy, drug-induced liver damage, heart disease, muscle disorders, and autoimmune or hematological diseases. Healthy controls also were selected who had the same inclusion criteria along with anti-HCV antibody and HCV RNA-negative results. The presence of liver cirrhosis was ascertained by the occurrence of ascites, esophageal varices and/or bleeding, splenomegaly, jaundice, and imaging. HCC was diagnosed by four-phase multidetector computed tomography scan and dynamic contrast-enhanced magnetic resonance imaging as mentioned elsewhere (27). Ninety-six patients had liver cirrhosis and 60 had HCC. Rests of the 110 patients were categorized as chronic hepatitis patients. Informed consent was obtained from all individual participants of the study. The study was approved by the institutional ethical committee.
Detection of IL-10 and IL-12B polymorphisms
SNPs in IL-10 promoter region polymorphisms (−1082 [rs1800896], −819 [rs1800871], and −592 [rs1800872]) and IL-12B −1188 [rs3212227] were genotyped by polymerase chain reaction–restriction fragment length polymorphism (PCR-RFLP). DNA was isolated from whole blood using Qiagen DNA isolation kit (Qiagen, Germany). The primers used, PCR products obtained, and restriction enzymes used in the study are shown in Table 1. PCR amplification was carried out in total volume of 20 μL by the following protocol: denaturation at 95°C for 3 min, followed by 35 cycles of denaturation at 95°C for 30 sec, annealing at different temperatures (Table 1) for 35 sec, and extension at 72°C for 50 sec. This was followed by a final extension at 72°C for 10 min.
The Primers Used, Product Sizes, and Restriction Enzymes Used for Different Single Nucleotide Polymorphisms Studied
SNP, single nucleotide polymorphism.
RFLP was performed by incubating the PCR product with specific restriction enzyme (MnlI for IL-10 − 1082 A/G, MaeIII for IL-10 − 819 C/T, RsaI for IL-10 − 592 C/A) according to the manufacturer's instructions. RsaI cuts IL-10 − 592 A allele to generate two bands (252/178 bp), MaeIII cuts IL-10 − 819 C allele into two fragments (272/171 bp), and MnlI cuts IL-10 − 1082 G allele into three fragments (84/23/23 bp). RFLP for IL-12B −1188 polymorphism was carried out with TaqI restriction enzyme according to the manufacturer's instructions. TaqI cuts IL-12B C allele into two fragments (269/100 bp). The digested products were run on a 2.5% agarose gel and visualized under ultraviolet light using ethidium bromide as dye.
Multiplex cytokine assay
The analysis of cytokine expression in serum was performed by multiplex enzyme-linked immunosorbent assay by following the manufacturer's instruction of MILLIPLEX®MAP kit named Human Cytokine/Chemokine Magnetic Bead Panel, catalog number HCYTOMAG-60K (Merck, Millipore). The instrument 40-072 MAGPIX® System was used to run HCYTOMAG-60K kit. Serum cytokines including IL-10 were analyzed (others not in the scope of this work). Twenty random samples were selected taken from each disease group and the healthy control group. The kits were used according to the manufacturer's recommendations. Sera were divided into two 25 μL aliquots, with both aliquots of serum from each subject placed in adjacent wells in the same plate. Each plate contained sera from each group of subjects. Each plate also contained two replicates of a “control” serum (taken from a middle-aged male), calibration samples to aid in converting mean fluorescent intensity values to units of concentration, and custom assay control beads designed to assist in detecting experimental failures.
Briefly, samples were mixed with anti-cytokine antibody-linked polystyrene beads on 96-well filter-bottom plates and incubated at room temperature for 2 h followed by overnight incubation at 4°C with continuous shaking at 500–600 rpm. The plate was washed twice with wash buffer and biotinylated detection antibodies were added for 2 h at room temperature. Samples were then filtered and washed twice and resuspended in streptavidin–phycoerythrin for 30 min at room temperature. Two additional washes were provided, and samples were resuspended in Reading Buffer. Each sample was measured in duplicate. The plate was run on MAGPIX with xPONENT® software. Median fluorescent intensity data were analyzed using a five-parameter logistic or spline curve-fitting method for calculating cytokine concentrations in pg/mL in samples.
Statistical analysis
Qualitative data are presented as numbers and percentages, and continuous variables are presented as means ± standard deviations. Genotypes and allele frequencies of the SNPs were compared by using Pearson's chi-square test or Fisher's exact test. The potential factors different from a p < 0.05 in the univariate analysis were included in the multivariate analysis based on a stepwise logistic regression model. SHEsis software was used for haplotype analyses. The assumption of the Hardy–Weinberg equilibrium was assessed for all SNPs using chi-square test. The Pearson correlation analysis was performed to find correlation within specific variables. For all statistical studies, the level of significance was taken at p < 0.05. The Bonferroni correction was used in multiple testing (p < 0.05, divided by the number of SNPs/haplotypes analyzed). The statistical analysis of the data was performed using SPSS software version 22.0 (IBM, New York City, NY).
Results
Patient characteristics
The general characteristics of the patients enrolled in the study are shown in Table 2. Patients who were diagnosed to have HCC had a statistically significant higher mean age than those of healthy controls and chronic hepatitis patients (p < 0.01) but not significantly higher than cirrhotic patients. The number of males among HCC patients was significantly higher than that among healthy controls but not with chronic hepatitis and cirrhotic patients. A significantly lower platelet count, hemoglobin, and serum albumin were observed in HCC patients than in healthy controls and chronic hepatitis patients (p < 0.001). Total bilirubin was higher in cirrhotic patients than in HCC patients but statistically not significant. Alanine transaminase (ALT) and aspartate aminotransferase (AST) were significantly elevated in all HCC in comparison with all other groups (p < 0.001). The number of patients with diabetes was considerably higher in HCC patients than in healthy controls and chronic hepatitis patients but not in cirrhotic patients.
Baseline Characteristics of the Study Population
Data are expressed as mean ± standard deviation or number.
Significant differences with control.
Significant differences with CHC.
Significant differences with CIRR.
ALT, alanine transaminase; AST, aspartate aminotransferase; CHC, chronic hepatitis C; CIRR, liver cirrhosis; HCC, hepatocellular carcinoma.
IL-10 − 1082, −819, −592 genotype distribution in HCV-infected patients
The IL-10 − 1082, −819, and −592 SNP genotype distribution and allelic frequencies among healthy controls and HCV-infected patients are shown in Table 3. No deviation from the Hardy–Weinberg equilibrium was seen for any of the SNPs. Genotype and allele frequency distributions were compared using chi-square test. The AA genotype of IL-10 − 592 and the CC genotype of IL-10 − 819 were significantly lower in HCV-infected patients than in healthy controls (24.0% vs. 14.7%, p < 0.05 and 26.0% vs. 17.7%, p < 0.01, respectively). Heterozygous mutant genotype, mutant allele carrier genotypes, and mutant alleles, however, were not significantly different in HCV-infected patients and healthy controls. For IL-10 − 1082 genotype, there was no significant variation between any of the genotypes or alleles. Table 4 shows the genotype and allele frequency distributions of IL-10 SNPs in the studied disease cases, which were compared using Fisher's exact test. IL-10 − 592 genotypes were significantly different in HCC cases compared with chronic hepatitis and cirrhosis cases. The mutant AA and CA genotypes were significantly lower in HCC cases than in chronic hepatitis cases (8.4% vs. 18.2%, p < 0.05 and 18.3% vs. 35.4%, p < 0.01, respectively). The frequency of mutant allele carrier genotypes CA and AA together was also lower in HCC patients compared with both chronic hepatitis and cirrhotic patients (26.7% vs. 53.6%, p < 0.01 and 26.7% vs. 43.7%, p < 0.05, respectively). The A allele was also significantly less in HCC patients than in both chronic hepatitis and cirrhotic patients (17.5% vs. 35.9%, p < 0.01and 26.7% vs. 29.2%, p < 0.05, respectively). We failed to observe any significant variation for any other polymorphic site in HCC patients.
Genotype Distributions of IL-10 −592, −1082, −819 and IL-12B −1188 Polymorphisms in Controls and Hepatitis C Virus-Infected Patients
Chi-square (χ 2) test was used. Significant values are shown in bold.
CI, confidence interval; HCV, hepatitis C virus; OR, odds ratio.
Genotype Distributions of IL-10 −592, −1082, −819 and IL-12B −1188 Polymorphisms Among the Three Study Groups and Association Analyses of These Single Nucleotide Polymorphisms and Hepatitis C Virus Infection Outcomes
Fisher's exact test was used. Significant values are shown in bold.
p: Probability value, p1: compares CHC and CIRR; p2: compares CHC and HCC; p3: compares CIRR and HCC.
IL-10 − 1082, −819, −592 haplotype distribution in HCV-infected patients
Haplotype analysis was performed in the IL-10 genes between all HCV patients and healthy controls as well as between HCC and other categories of HCV-infected patients using the SHEsis software. The haplotype frequency distribution between healthy controls and HCV-infected patients is shown in Table 5. Among the IL-10 − 1082, −819, and −592 haplotypes, ATC was most abundant in both healthy controls and HCV-infected patients (29.5% vs. 36.6%, p = 0.069). GCC (11.5% vs. 6.5%, p < 0.05), ATA (11.5% vs. 5.4%, p < 0.01), and ACA (10% vs. 4.8%, p < 0.05) haplotype frequencies were observed to be differently expressed in healthy controls and HCV-infected patients. When taken together, GCA, ATA, and ACA haplotypes were seen to be significantly lower in HCV-infected patients than in healthy control subjects (16.9% vs. 33.0%, p < 0.001). No other haplotypes had different frequencies between the controls and patients. Haplotype frequency distribution of IL-10 SNPs was analyzed in different disease cases of HCV patients (Table 6). In HCC patients, ACC haplotype (27.6% vs. 12.3%, p < 0.01) was significantly higher and GCA haplotype (6.7% vs. 15%, p < 0.05) was lower than that in chronic hepatitis patients. In cirrhosis cases, ACC haplotype (25.0% vs. 12.3%, p < 0.01) was significantly higher and ACA haplotype was significantly lesser than that in chronic hepatitis cases (2.6% vs. 6.3%, p < 0.05). Other haplotype frequencies did not show significant variations among the HCV-infected disease cases. GCA, ATA, and ACA haplotypes, when considered together, were seen to be significantly lower in cirrhotic (10.9% vs. 22.3%, p < 0.01) and HCC (12.5% vs. 22.3%, p < 0.05) patients than in chronic hepatitis patients.
Distribution Frequencies of IL-10 (−1082, −819, −592) Haplotypes in Controls and Hepatitis C Virus-Infected Patients
Fisher's exact test was used. Significant values are shown in bold.
Distribution Frequencies of IL-10 (−1082, −819, −592) Haplotypes in Studied Disease Groups
Fisher's exact test was used. Significant values are shown in bold.
p: Probability value; p1: compares CHC and CIRR; p2: compares CHC and HCC; p3: compares CIRR and HCC.
IL-12B −1188 genotype distribution in HCV-infected patients
Results of the IL-12B −1188 SNP analysis in healthy controls and HCV-infected patients are depicted in Table 3. No significant variation of the IL-12B −1188 genotypes or alleles was observed between healthy controls and HCV-infected patients in this population (p > 0.05). Similarly, while comparing IL-12B −1188 genotype and allele frequency distributions among the HCV-infected disease groups, no significant variation was observable in our study (p > 0.05) (Table 4).
Correlation of HCV viral load with IL-10 and IL-12B SNPs
HCV viral loads in all categories of patients were correlated with IL-10 − 1082, −819, −592 and IL-10 −1188 polymorphisms. The mean HCV viral load was only marginally higher in G allele of IL-10 − 1082 and A allele of IL-10 − 592 in CHC cases. In case of IL-10 − 819, significantly lower viral load was seen in patients with C allele (p < 0.05). No significant difference was also seen in the mean viral loads when comparing IL-12B −1188 alleles in CHC patients. In cirrhotic and HCC patients, no significant differences were observed in the mean viral loads for any of the SNPs studied (Supplementary Fig. S1).
Serum concentrations of IL-10 in healthy controls and HCV-infected patients
The serum expression of IL-10 was categorized according to the pathological diagnosis of HCV-infected patients (Fig. 1). The mean serum expression in controls was 12.24 ± 9.52 pg/mL. IL-10 concentrations in chronic hepatitis patients were marginally higher at 17.01 ± 13.83 pg/mL (p = 0.502). In comparison, the serum IL-10 level was significantly higher in cirrhotic (41.24 ± 13.83 pg/mL) and HCC patients (55.78 ± 25.89 pg/mL) than in controls and chronic hepatitis patients (p = 0.072, p < 0.001, for controls and p < 0.05, p < 0.001, for chronic hepatitis, respectively).
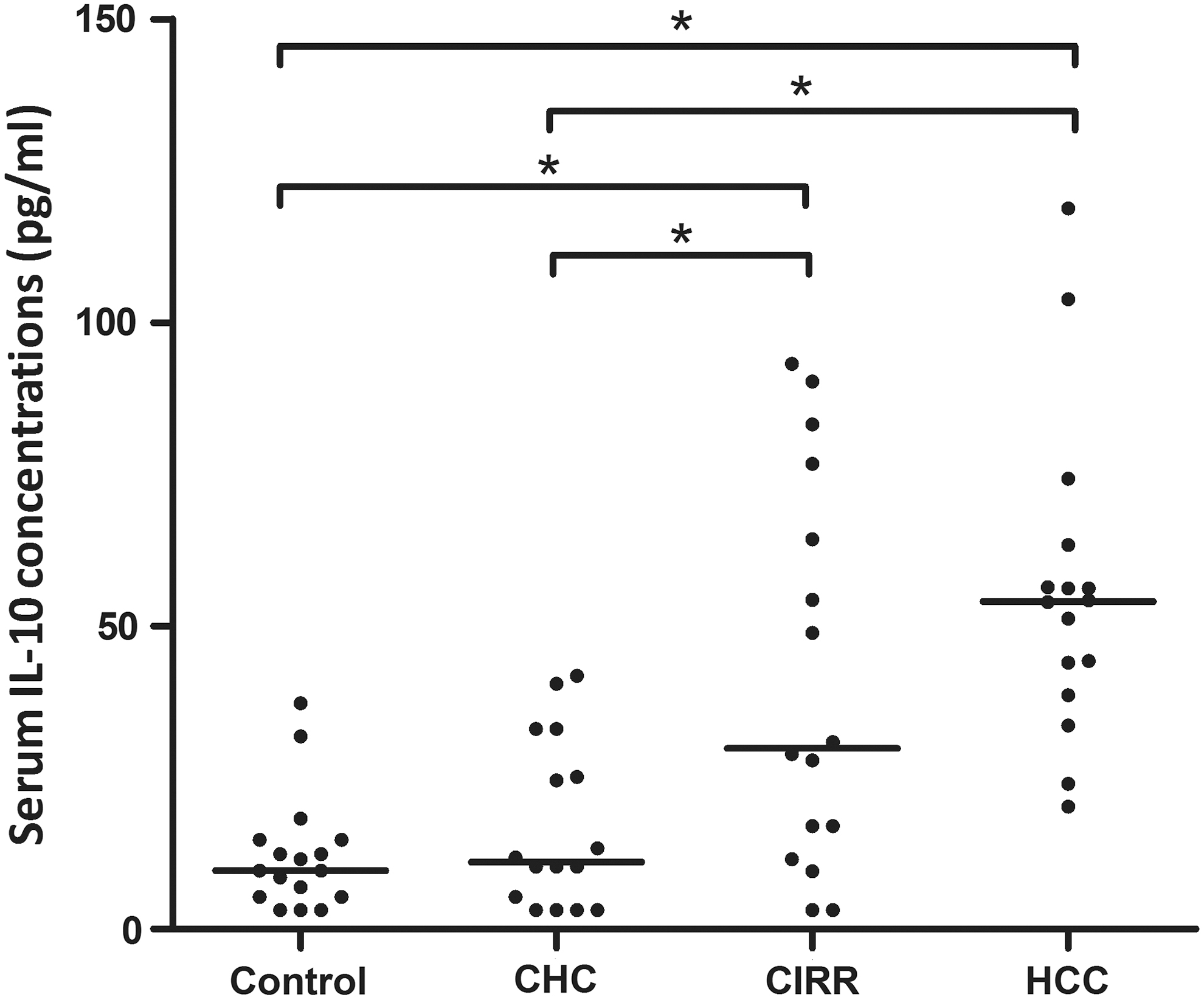
Dot plot representation of serum IL-10 concentrations in disease groups and controls. *p < 0.05, CHC, chronic hepatitis C; CIRR, liver cirrhosis; HCC, hepatocellular carcinoma.
IL-10 levels in subjects with different IL-10 SNPs
The mean serum IL-10 levels in studied disease groups and controls were calculated according to IL-10 − 1082, −819, −592 alleles and haplotypes. We observed that although there was a marginal increase in serum IL-10 concentrations with IL-10 − 1082 G allele, it was not statistically different in any of the disease stages or controls (p > 0.05). Similarly, for IL-10 − 819 SNP, the serum IL-10 level was marginally higher C allele carriers in all groups. For IL-10 − 592, A allele carriers had slightly higher serum IL-10 levels. No statistically significant differences were seen. We also compared serum cytokine concentrations in patients with IL-10 − 1082, −819, −592 ACC haplotypes and GCC/ATA/ACA haplotypes with those who did not carry any of these haplotypes. We found that there was a significant reduction in serum concentration of IL-10 in cirrhotic patients (p < 0.01) with GCC/ATA/ACA haplotypes. In HCC patients, serum IL-10 levels were also reduced in those carrying GCC/ATA/ACA haplotypes, although the relationship was not significant (p > 0.05). Serum levels of IL-10 were reduced in all groups with ACC haplotypes compared with other haplotypes (Table 7).
Mean Serum IL-10 Levels in Studied Disease Groups and Controls Segregated According to IL-10 −1082, −819, −592 Alleles and Haplotypes
p < 0.05 (Mann–Whitney U test was used).
Correlation of HCV viral load with serum IL-10 concentrations
In each disease category, the HCV viral load was correlated with serum concentrations of IL-10. No significant correlation was observed between HCV viral load and IL-10 concentrations in CHC and cirrhosis cases (p > 0.05). In patients with CHC and cirrhosis, the Pearson correlation coefficients (r) was 0.301 and 0.231, respectively. The serum IL-10 level was significantly correlated with HCV viral load in cases of HCV-infected patients with HCC, with the Pearson correlation coefficient (r) equals to 0.665 (p < 0.05) (Fig. 2).
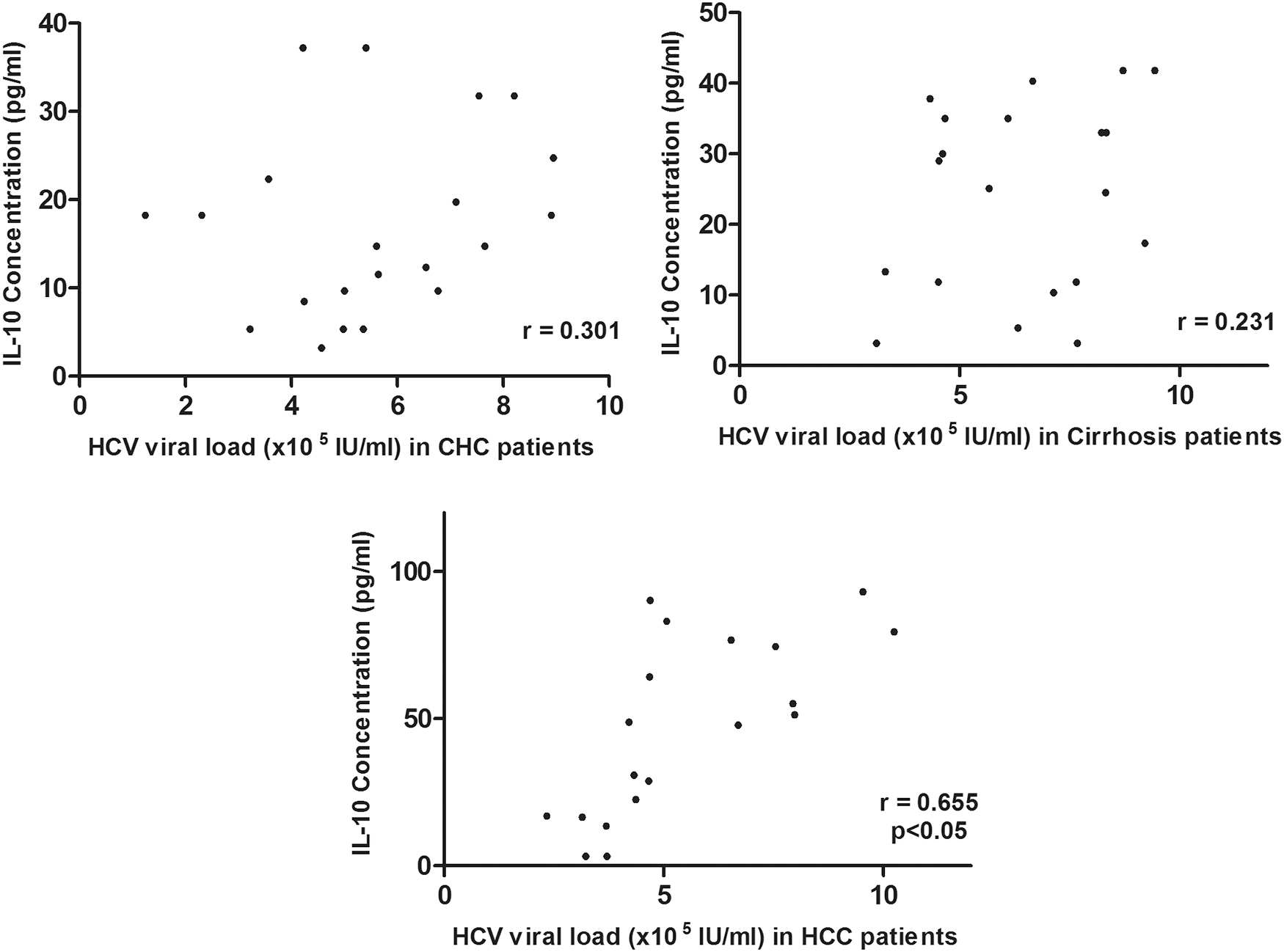
Correlation of HCV viral load with serum IL-10 concentrations in various disease stages. HCV, hepatitis C virus.
Factors associated with HCC development
To identify factors associated with HCC development, univariate analysis was carried out as shown in Table 8. Old age, increased AST and ALT levels, serum albumin, viral load, IL-10 − 592 CC genotype, IL-10 − 1082, −819, −592 ACC haplotype, absence of GCA/ATA/ACA haplotypes, and heightened serum IL-10 levels were significantly associated. Stepwise regression analysis showed that ALT (odds ratio [OR]: 3.119, 95% confidence interval [CI]: 2.597–4.088, p < 0.05), IL-10 − 592 CC genotype (OR: 3.572, 95% CI: 2.514–4.127, p < 0.01), IL-10 ACC haplotype (OR: 9.447, 95% CI: 4.561–15.364, p < 0.001), and serum IL-10 concentrations (OR: 3.541, 95% CI: 2.314–5.628, p < 0.01) were the independent predictors of HCC development. Conversely, there were no significant relationships of other SNPs studied and the risk of HCC development (p > 0.05).
Univariate Analyses and Stepwise Multivariate Analyses of Factors Associated with the Risk of Hepatocellular Carcinoma in Patients with Chronic Hepatitis C Virus Infection
Bonferroni-corrected p-value. Significant values are shown in bold.
Discussion
Polymorphisms in the promoter region of IL-10 and the haplotypes have been linked with variations in IL-10 production in each individual, and IL-10 genotype frequency has been shown to be highly variable in people with different genetic backgrounds (1,15). The IL-12B 3′UTR 1188 polymorphism has been associated with increased cancer risks in Asians in previous studies (10,55). The present study was aimed at analyzing the possible associations of polymorphism in IL-10 − 1082, −819, −592 and IL-12B −1188 and development of liver cirrhosis and HCC in the Northeast Indian population. Association of the IL-10 SNPs on cytokine concentrations in serum of HCV-infected patients was also analyzed.
Studies have reported an increased frequency of IL-10 − 1082 GG genotype and G allele in the HCV-infected patients and higher frequency of A allele in the healthy group (13). This suggests that individuals who have this high IL-10 producer G allele or GG genotype are more susceptible to chronic HCV infection, whereas the presence of the A allele or −1082 AG genotype confers protection (19,39). Although serum levels of IL-10 were elevated in patients with G allele, we failed to show any significant variation in IL-10 − 1082 GG genotype distribution between healthy controls and HCV-infected patients as also demonstrated by others (27).
IL-10 − 1082 A allele was associated with higher rates of HCC (5). In study subjects from Argentina, HCV-infected patients with IL-10 − 1082 G allele had a lower risk of progression to cirrhosis (32). On the contrary, studies have indicated no significant influence of the high IL-10-producing IL-10 − 1082 GG genotype/G allele degree of liver fibrosis in Australian patients or Brazilian patients or other higher severity liver affection such as liver cirrhosis and HCC in HCV genotype 4-infected Egyptian patients (7,20,28,34). No association was observed between IL-10 − 1082 polymorphisms and HCC in two independent meta-analyses (47,51). In accordance, we failed to observe any association of IL-10 − 1082 A/G polymorphism with increased disease severity.
For IL-10 − 819 T/C and −592 C/A SNPs, no significant difference in distribution was observed among HCV-infected patients and healthy controls in multiple studies (9,12,13,29,39). Also, a meta-analysis revealed a weak association of IL-10 − 592A allele with HCV infection in Asian population and but not for alleles at positions −1082 and −819 (26). A higher occurrence of IL-10 − 819 C allele was observed in HCV patients in a Pakistani population (3). We found IL-10 − 819 CC genotype to be more abundant in controls than in HCV-infected patients. No significant variation was observed for IL-10 − 592 C/A polymorphism.
No association was found between HCC susceptibility and IL-10 − 819 T/C polymorphism, most studies pertaining to Asian populations (29,47,54). But IL-10 − 819 TC genotype was shown to confer fibrosis protection in another study in Brazilian patients (28). Nevertheless, it was observed that the −819/592 TT genotypes are significantly more common among patients with multiple nodules and in advanced stages of HCC (5). Our results showed no significant association of IL-10 − 819 T/C SNP with liver disease severity in HCV infection.
Studies have shown that the IL-10 − 592 polymorphism contributes to cancer susceptibility, especially in smoking-related cancer and Asians (14). The heterozygous CA variant in the IL-10 − 592 has been associated with a major risk of progression of viral hepatitis (30). At the same time, two independent meta-analyses have clearly suggested that the IL-10 − 592 C allele, which is a low IL-10 producer allele and the CC genotype, increases HCC susceptibility in Asian populations (47,54). In our study, the AA genotype and A allele were significantly reduced in patients with HCC than in patients with chronic hepatitis and cirrhosis. The A allele at IL-10 − 592 showed reduced IL-10 cytokine levels in all patient groups.
Frequencies of IL-10 promoter haplotypes in different populations significantly vary (42). Haplotype analysis revealed that ATA haplotype was more frequent in the control group, and GTA and GCC haplotypes were more frequent in HCV patients (3,13). Conflicting reports were generated in a study conducted by Sepahi et al. They found that the ATA haplotype was more prevalent in HCV patients in comparison with controls. In our study, we found that GCC, ATA, and ACA haplotypes were significantly higher in control subjects when compared with HCV-infected patients. At the same time, few studies have also reported the IL-10 promoter haplotype frequencies to be not significantly different between these two groups (16,24,31).
IL-10 ACCT haplotype consisting of IL-10 − 1082, −819, −592, +117 was associated with increased occurrence of HCC in HBV-infected patients (40). Similarly, Tseng et al. also have reported ACC haplotype to be associated with HCC risk in Taiwanese patients (45). In HCV-infected Brazilian patients, no impact of ACC haplotype was observed in the risk of HCC occurrence (5). Interestingly, in a study on Polish patients, ACC haplotype was associated with a reduced risk of nonresponse and relapse in antiviral therapy (43). In our study, we found the ACC haplotype of IL-10 to be significantly higher in patients with cirrhosis or HCC than in patients with chronic hepatitis. Thus, ACC haplotype, which is an intermediate IL-10-producing haplotype, may represent a potential biomarker for cirrhosis or HCC in this population. Also, the GCC, ATA, and ACA haplotype frequencies when observed together were found to be lesser in cirrhotic and HCC patients than in chronic hepatitis patients.
Homozygosity for the A allele at position −1188 of the IL-12B gene has been associated with HCV persistence in multiple populations (22,52,53). This polymorphism has been associated with increased cancer risks in the subgroups of Asians in a previous meta-analysis (10,55). The same was also found in HCC, where IL-12B 3′UTR 1188 polymorphism was observed to be a significant risk factor for HCC development, especially among Asians (33). In a cohort of Indian patients with HBV infection, IL-12B −1188AC had a suspicious role in HCC occurrence (37). In HCV-induced HCC patients, no statistically significant difference was seen between the alleles or between the genotypes with the cancer-free chronic HCV patients, which is supported by results of the current study (17,44). One recent study in Egyptian patients has reported AA genotype to be significantly correlated with cirrhosis and HCC (18).
Conclusions
The IL-10 − 592 genotype is a marker for HCV susceptibility as well as HCC and cirrhosis occurrence in CHC patients from Northeast India. IL-10 ACC haplotype also may predispose CHC patients to higher HCC risk, whereas GCC, ATA, and ACA haplotypes may confer protection from HCV infection. Furthermore, IL-10 − 1082 and −819 along with IL-12B −1188 SNPs do not qualify as markers of HCC. With the increasing burden of HCV-infected liver disease patients in Northeast India, these potential biomarkers may have prognostic value by helping patients with chronic HCV infection and nonfavorable genetic makeup in deciding to undergo a more aggressive therapy to reduce risk of evolving more severe forms of the disease. The sample size included in the present study is one of the limitations of the study. A more detailed study involving a larger number of patients is required to shed further light into the potential of IL-10 and IL-12B SNPs as biomarkers to determine risk of cirrhosis and HCC in HCV-infected patients from Northeast India.
Footnotes
Acknowledgments
We would like to acknowledge the help provided by all members of Gastroenterology Outpatient Department, Gauhati Medical College, in the collection of samples, and members of the Department of Bioengineering and Technology, Gauhati University, for their technical inputs.
Ethical Approval
All procedures performed in studies involving human participants were in accordance with the ethical standards of the institutional and/or national research committee and with the 1964 Declaration of Helsinki and its later amendments or comparable ethical standards.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was funded by the Department of Biotechnology, Government of India (Grant no. HC-181, BCIL/NER-BPMC/2012/1549).
Supplementary Material
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.