
Abstract
Immunization with hepatitis B vaccine is an effective measure for prevention and control of hepatitis B Virus (HBV) infection. Although lots of efforts to improve the effect of hepatitis B vaccine have been made, the function of human beta defensin 2 (hBD2) on hepatitis B vaccine keeps unclear. In this article, we report that hBD2 not only promoted the activation and maturation of immature dendritic cells (iDCs) by increasing MHC II and CD86 expression, but it also significantly upregulated the mRNA level of IL-6 and IL-12B in mouse bone marrow-derived dendritic cells. The serum concentrations of IFN-γ in mice stimulated with 300 ng hBD2 increased from 25.21 to 42.04 pg/mL, with a time extension from 4 to 12 h post-injection. During the process of three times immunization (1, 14, 28 days) with 3 μg hepatitis B vaccine combined with or without 300 ng hBD2 with a 2 week interval in BALB/c mice, the antibody against HBsAg (HBsAb) concentration in serum at every time point of observation in the combined group was statistically higher than the hepatitis B vaccine group. The serum concentration of IgG2a subclass HBsAb on the 14th day post last injection in the combined group was significantly higher than the hepatitis B vaccine group. Further, the splenic cells from the mice treated with both hBD2 and hepatitis B vaccine possessed a greater ability to produce a surface antigen of hepatitis B virus (HBsAg) specific IFN-γ than those treated with hepatitis B vaccine alone. The percentages of CD3+/CD4+ T cells and CD3+/CD8+ T lymphocytes in spleens from the mice treated with 300 ng hBD2 were statistically higher than the phosphate buffered saline group. These data suggest that hBD2 improves iDC maturation and the immune efficiency of hepatitis B vaccine in BALB/c mice.
Introduction
Hepatitis B virus (HBV) has been a major cause of human hepatitis (32). Although hepatitis B could be prevented by hepatitis B vaccine, hepatitis B resulted in 27 million infected people in 2016 and about 887,000 deaths in 2015 in the world (12,55). hepatitis B prevalence maintains the highest level in the Western Pacific Region and African Region, where 6.2% and 6.1% of the adult population is infected by HBV, respectively (55).
Since 1982, aluminum salt has been available as an immune adjuvant in hepatitis B vaccine (33). Although the immunogenicity of the surface antigen of hepatitis B virus (HBsAg) in hepatitis B vaccine has been widely accepted, the defects of alum adjuvant in the vaccine and problems of hepatitis B vaccine related with alum adjuvant have been a matter of global concern for a long time (10). First, the alum adjuvant in the vaccine may induce inflammation and local reactions at the site of injection (1,24) and increase the production of IgE antibody as part of the overall Th2 profile response (1,5,18). Second, from the view of immune efficiency, hepatitis B vaccine with alum elicits weak or even no response under the regular three-dose immunization in about 5–15% of the health population (2). Third, this kind of hepatitis B vaccine results in low sera reactions in patients with renal insufficiency (13,16), inflammatory bowel disease (15,48), and chronic HCV infection (8) and it works unsuccessfully in immunosuppressive or immunodeficient patients (33).
However, at last, the content of aluminum in the human body comes mainly from drinking water (47), foods such as infant formulae (9), and even bad behaviors such as smoking. A great majority of aluminum in the bloodstream is able to be cleared by the renal system (37). More attention should be paid to the safety of various vaccines containing alum, such as mixed diphtheria pertussis tetanus (DPT) vaccine and hepatitis B vaccine, because the absorption rate of aluminum from the vaccine through injection is nearly 100%, and excess aluminum may be deposited in the spleen, thymus, lungs, and bones, and lead to many local or systemic toxic effects, including immunotoxicity and the inhibition of lymphocyte and macrophage functions (27,43,57,63). The known side effects after an intramuscular injection of alum-containing vaccines include local granulomatous inflammation at the vaccination site and constellation of neurologic symptoms, such as myalgia, arthralgia, chronic fatigue, weakness, and cognitive issues (10,36,42). Children are usually prone to be vulnerable to aluminum toxicity, because the programmed vaccinations, including DPT vaccine and hepatitis B vaccine, have been performed to a very great extent in infant to child stages and lots of toys for children to play are made of aluminum. So, the alum salt in vaccines could sometimes turn into poison if the concentration of alum in blood or organs reaches over the limit (14). Reasonably, the total acceptable amount of alum dose for human vaccines (1.0–1.5 mg/vaccination) would limit the alum adjuvant to potentially carry several antigens (5). Therefore, people believe that it is well worth to find a safer and more effective substitute for aluminum used in the hepatitis B vaccine in future. A typical example is that a novelty adjuvant named CpG 1018 containing repeating synthetic oligodexynucleotide sequences of cytosine phosphoguanosine (CpG) has been successfully invented and used to develop a new type of hepatitis B vaccine named HEPLISAV-B, which was licensed for adults by the US Food and Drug Administration in November 2017. According to the report, this 2-dose hepatitis B vaccine provided a higher level of seroprotection (85.8%) against HBV than a 3-dose vaccine with alum adjuvant (58.5%) in patients aged 60–70 years with type 2 diabetes mellitus (19).
Defensins, first discovered in 1985, belong to the family of antimicrobial peptides and could be classified into three categories (alpha-, beta-, and theta-defensins) (3). Among the human β-defensins (hBDs) subfamily, human β-defensin 2 (hBD2) can not only function as a stimulator to make keratinocyte proliferation, migration, and wound healing (27), but it is also highlighted due to its immune activation ability in dendritic cells (DCs) and thus contributes to the innate and adaptive immune response. During the past few decades, lots of papers reported the immunity improvement or immune adjuvant effect of beta-defensin 2 from human or murine cells. In 2013, Vemula et al. (49,50) reported that murine beta-defensin 2 showed a better adjuvant effect in the DNA vaccine of H5N1 influenza virus. In 2018, Kim et al. (25) reported that hBD2 improved the immunogenicity of spike protein from Middle East respiratory syndrome-coronavirus (MERS-CoV) and the antiviral immunity by increasing the IFN-β, IFN-γ and TNF-α, IL-1β, and IL-6 expression as an adjuvant, both in vitro and in vivo. In 2019, Sun et al. (46) reported that hBD2 enhanced the immune therapy effect against melanoma in mice. However, the immune function of hBD2 on HBsAg as an adjuvant is unclear. So this research was performed to investigate whether hBD2 enhances the immunity of hepatitis B vaccine and found for the first time that hBD2 increased the production of serum antibody against HBsAg (HBsAb), especially IgG2a subclass, and IFN-γ in BALB/c mice. These results imply that hBD2 could be a good candidate as a natural adjuvant to improve the immune efficiency of hepatitis B vaccine.
Materials and Methods
Animals
Six- to eight-week-old female BALB/c mice were purchased from the animal center of Southern Medical University and fed with a standard laboratory diet at 20°C–22°C. All animals involved were in compliance with the Guide for the Care and Use of Laboratory Animals, which was issued by the National Institutes of Health in 1996. The animal procedures were approved by the Institutional Animal Care and Use Committee of Nanchang University, and the number of the experiment permit is SYDWLL2018–136.
Generation, differentiation, and initial identification of bone marrow-derived DCs from Balb/C mouse
According to the classical method reported by Son et al. (44), 2 × 106 bone marrow-derived dendritic cells (BMDCs) from the femurs and tibias of the Balb/C mice were cultured in 10 mL complete RPIM 1640 (Invitrogen) supplemented with 10% heat-inactivated FCS, 100 U/mL penicillin, and 100 μg/m streptomycin, in the absence or presence of 40 ng/mL recombinant granulocyte macrophage colony-stimulating factor (rmGM-CSF) and 20 ng/mL rmIL-4 (PeproTech, Suzhou, Jiangsu, China) for 3 days. Then, the adherent cells were dislodged by using a cell scraper and cultured in 10 mL complete RPMI 1640 containing 20 ng/mL rmGM-CSF and 10 ng/mL rmIL-4 for another 3 days. During the incubation, the cell morphology was observed daily under an inverted microscope. At day 6, the levels of CD11c, MHC-II, CD86, and CD80 on the membrane of BMDCs were tested by flow cytometry.
BMDCs maturation and cytokine(s) expression under different concentrations of hBD2 treatments ex vivo
The recombinant hBD2 peptide, which was composed by a sequence of GIGDPVTCLK SGAICHPVFC PRRYKOIGTC GLPGTKCCKKP, was purchased from Protein Specialists (ProSpec). The concentration of lipopolysaccharide (LPS) in hBD2 peptide powder that was verified was less than 0.01% according to the instructions, and the correctness of cyst connection was also verified by the producer. They were resolved with phosphate-buffered saline (PBS) before use. Two × 105 BMDCs, which were separated and cultured for the first 6 days as described earlier, were continuously cultured in 1 mL complete RPMI 1640 medium containing 20 ng/mL rmGM-CSF, 10 ng/mL rmIL-4, and different final concentrations of hBD2 from 0, 12.5, 25, and 50 to 100 ng/mL for another 24 h. Then, the membrane expression of CD11c, MHC-II, CD86, and CD80 was tested by flow cytometry (FCM) assay. On the other hand, relative quantities of cytokines, including IL-6, IL-12B, and IFN-γ, were determined by quantitative reverse transcriptase polymerase chain reaction (qRT-PCR) on the Applied Biosystem 7500 Real-Time PCR System, and they were normalized based on β-actin expression for each sample. The primer information and the conditions of qRT-PCR for IL-2, IL-6, IL-12B, and IFN-γ are detailed in Supplementary Data S1.
The patterns of IL-6, IL-12, and IFN-γ expression with time extension after hBD2 treatment in Balb/C mice
Thirty-six mice, aged 6–8 weeks, were randomly divided into control and experimental groups after a 1-week adaptive husbandry. The mice in the control group intramuscularly received 200 μL PBS, whereas the mice in the experimental group received an equal volume of PBS containing 300 ng hBD2. Orbital bleeding in the circumstance of ether anesthetization was performed at 4, 8, and 12 h post-injection. After that, the mice were immediately sacrificed by cervical dislocation and spleens were collected in a clean bench after the disinfecting action of 75% alcohol immersion at the same time. The sera were used to detect the concentration of cytokines IL-12p70 (ab119531; Abcam) and IFN-γ (ab100689; Abcam) by an enzyme-linked immunosorbent assay (ELISA). The spleens were dissected into small pieces to extract total RNA by the TRIZOL method to test mRNA levels of IL-6, IL-12B, and IFN-γ to β-actin by real-time RT-PCR.
The immune efficiency evaluation of hBD2 on hepatitis B vaccine in Balb/C mice
Forty mice were randomly grouped into four categories after a 1-week adaptive husbandry. Each mouse was intramuscularly immunized at week 0 as follows: Group 1: immunization with 200 μL PBS; Group 2: immunization with 200 μL PBS containing 300 ng hBD2; Group 3: immunization with 200 μL PBS containing 3 μg HBsAg in recombinant hepatitis B vaccine (Genetech Biotechnology, Huabei Pharmaceutical Co., Ltd., Shijiazhuang, China); and Group 4 (named combined group): first immunization with 200 μL PBS containing 300 ng hBD2, then 200 μL PBS with 3 μg HBsAg in hepatitis B vaccine by another injection at the same site after 4 h. At weeks 2 and 4, boost immunizations were given to mice by using the same reagent and dose. According to the introduction of the recombinant hepatitis B vaccine from the producer, this kind of vaccine was produced by HBsAg DNA expression in Chinese hamster ovary cells and contained some Al(OH)3 and NaCl solution, no AlPO4; this has been one of the most popular vaccines used in China for a long time (38,61,62). The sera that were collected by orbital bleeding in the circumstance of ether anesthetization at days 11, 22, 33, and 42 post-1st immunizations were used for the total HBsAb test by ELISA to value the change of HBsAb concentration. On day 42 after the first immunization, all mice were sacrificed and HBsAg-specific IFN-γ valuation in splenic monocytes was tested by ELISA. For sera collected on the last day, the concentrations of IL-2, IL-12p70, and IFN-γ and the levels of IgG1 and IgG2a were also tested by ELISA, respectively. The total plan of the experiment in this stage is shown next as a scheme figure (Fig. 1).
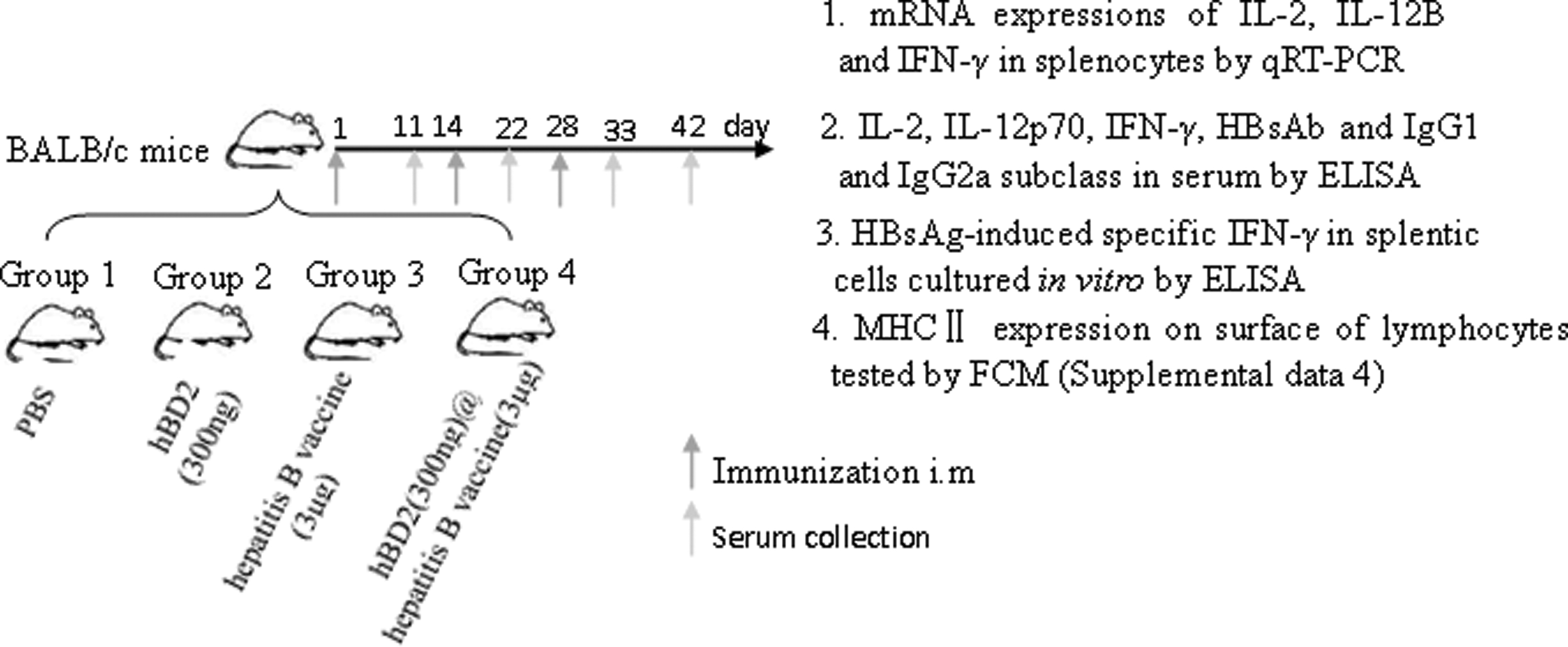
Experimental scheme of immunization with HBsAG and hBD2 in mice. HBsAG, surface antigen of hepatitis B virus; hBD2, human beta-defensin 2.
Analysis of IL-6, IL-12B, and IFN-γ mRNA levels by quantitative real-time RT-PCR
First, total RNA was extracted by using the High Pure RNA isolation kit (Roche Diagnostics, Mannheim, Germany). Thereafter, the absorbance of the resulting total RNA was measured at 260 nm, and the concentration was calculated. Then, cDNA was synthesized by using High-Capacity RNA-to-cDNA Master Mix® (Applied Biosystems, Foster City, CA). Treatment was performed according to the manufacturer's protocol, and adjustments were made to yield a total volume of 20 μL. The following conditions were used with a GeneAmp® PCR System 9700 (Applied Biosystems) to synthesize cDNA: 25°C for 10 min, 42°C for 60 min, 85°C for 5 min, and a 4°C hold. Then, to 1 μL of the resulting cDNA sample, 5 μL TaqMan® Universal Master Mix II (Applied Biosystems), 0.5 μL primer, and 3.5 μL RNAse-free water (Applied Biosystems) were added to yield a total volume of 10 μL. Primers for mice β-actin, IL-6, IL-12B, and IFN-γ were purchased from Applied Biosystems. Reactions were performed by using an ABI PRISM 7700 sequence detection system (Applied Biosystems) with 30 cycles of 94°C for 1 min, 60°C for 1 min, 72°C for 1 min, and a final 5-min extension at 72°C.
Double antibody sandwich ELISA for the tests of IL-2, IL-12p70, and IFN-γ concentration
According to the ELISA kit instructions for the quantitative measurement of Mouse IL-12 p-70, IL-2, and IFN-γ concentrations in serum, the standards with different concentrations and the serum samples were diluted at first. Three replicate wells for each sample, one well for each concentration of the standard, and one well for blank control were set. Then, 100 μL standards and 50 μL test samples were added to the corresponding wells and 50 μL of no protein control was added to the blank control well. Along with 50 μL of biotin-conjugated anti-mouse specific detection antibodies into all wells, the microplate was then incubated at room temperature for 2 h. After washing with wash buffer, 100 μL of diluted streptavidin-horseradish peroxidase (HRP) conjugate was added into each well, and the plate was incubated at room temperature for 2 h. After emptying wells and washing three times, 100 μL tetramethylbenzidine (TMB) substrate solution was added into all wells and the microplate was incubated in a dark room at room temperature under 25°C for 10 min. After adding 100 μL of stop solution into each well, the absorbance value of each well on a spectrophotometer was read and recorded by using 450 nm. Then, IL-2, IL-12p70, and IFN-γ concentrations of the samples were calculated according to their standard curves, which were made by the concentration and absorbance values of the corresponding standards, respectively.
Double antigen sandwich ELISA to test serum total HBsAb levels
HBsAb level was detected by double antigen sandwich ELISA with the HBsAb diagnostic kit (Yingke, Xiamen, China) as per the manufacturer's instructions. Briefly, blank, negative, and positive controls were designed in the same plate. Fifty microliters of sera sample and an equal volume of negative and positive controls were added into the corresponding wells. Then, 50 μL of biotin-conjugated HBsAg was added into all wells, and the microplate was incubated at 37°C for 1 h. After washing three times with PBST, 100 μL of diluted streptavidin-HRP conjugate was added into each well and the plate was incubated at room temperature for 2 h. After empting wells and washing three times, 100 μL of TMB substrate solutions was added into all wells. The microplate was incubated on a shaker set at 400 rpm at 37°C for 20 min. After adding 100 μL of stop solution into each well, the OD450 value for each sample was assayed in triplicate. The results of the test were expressed as the ratio between the optical density (OD) value of the signal detected on the sample and the cutoff value (S/CO). According to the kit instruction, the cutoff value in this test was calculated by the multiple between the average value of OD450 and two times of standard deviation of the negative controls. This is a semi-quantitative assay in which the S/CO value is directly proportional to the quantity of the antibody (23). In our experiment, it is directly proportional to the concentration of HBsAb, and the value of S/CO equal or greater than 1.00 was considered reactive.
Indirect ELISA to test the concentration of IgG1 and IgG2a subclasses against HBsAg in serum
The concentrations of IgG1 and IgG2a in mouse were tested by indirect ELISA according to the kit instructions. Six wells were set for each sample and positive control. Briefly, 50 μL samples were added into the well and incubated at 37°C for 1 h. The plate was washed and taped on absorbent paper. One hundred microliters of HRP conjugated Rabbit-anti mouse IgG1 or Rabbit-anti mouse IgG2a antibody was added into each well, and the plate was incubated at 37°C for another 2 h. Then, wells were washed and 100 μL TMB substrate solutions were added into all wells. After appropriate treatment, including incubation and washing, the stop solution was added and the value of OD450 was tested on a machine. Then, the cutoff value was made by double average values of the PBS group, and the ratio value of sample OD450 to cutoff (S/CO) for all samples was calculated for statistical analysis.
The test of HBsAg-specific IFN-γ expression in splenic cells from the mice with different schedule immunizations
On day 42 after the first immunization, all mice were sacrificed by cervical dislocation. The spleen was separated in a clean bench and gently ground into the monocytes on the mesh 200. Then the splenic monocyte suspension was prepared by the peripheral mononuclear blood cell preparation method. To test the levels of HBsAg-induced specific IFN-γ, 2 × 105 splenic mononuclear cells were collected and co-cultured in 1 mL complete RPIM 1640 culture medium with 0.2 μg/mL HBsAg for 18 h. The supernatant of cell culture was collected and used to detect IFN-γ secretion by double antibody sandwich ELISA assay, as previously described.
Flowcytometry assay for BMDCs maturation
To measure whether hBD2 had the ability to aid in BMDCs maturation, the BMDCs cultured with rmGM-CSF and rmIL-4 for 6 days, and the cells cultured with different concentrations of hBD2 treatment for another 24 h were collected as mentioned earlier. To remove the unspecific staining that was produced by the interaction between IgG Fc fragment on the fluorochrome-conjugated antibody and CD16 (IgG FcRIII) or CD32 (IgG FcR II) on the surface of BMDCs, 2 × 104/100 μL mouse antibody against CD16/CD32 (eBioscience, San Diego, CA) were used to block Fc receptors on BMDCs for 20 min at first. Then, each sample was divided into three subgroups in parallel and the membrane staining for each of the three subgroup samples was performed with 0.25 μL PE-labeled anti-mouse CD11c and 0.3 μL FITC-labeled anti-mouse MHC class II (MHC-II), 0.3 μL PE-labeled anti-mouse CD80, or 0.3 μL FITC-labeled anti-mouse CD86 (all from eBioscience, San Diego, CA), separately. During this period, 1 tube negative control that was not stained with any fluorochrome-conjugated antibody was set, and a single positive sample for four fluorescence-labeled antibodies was separately and individually stained at the same time. After incubation for 30 min at room temperature in a dark room, all tubes were centrifuged at 1,500 rpm for 5 min and the suspension was removed. Then, cells were washed three times with PBS, and 200 μL PBS for each tube was added and resuspended before FCM test. During the operation of FCM, the voltage of photomultiplier tube (PMT) and the gate for cells entering into analysis was set according to the distribution of the cells on scatter dot diagram by side scatter (SSC) and forward scatter (FSC) at first. Then PE-labeled CD11c was used as a gate to discriminate DC from other cells after adjustment of fluoresce compensation between PE and FITC by using single positive staining sample. At last, double parameters analysis between PE-labeled CD11c and FITC-labeled CD80, FITC-labeled CD86 and FITC-labeled MHC-II was performed, and the medians fluorescence intensity for MHC-II, CD11c, CD80 and CD86 was tested (Supplementary Data S2B). Cell events were counted and calculated through FACScalibur and CellQuest™ software (BD Biosciences, San Jose, CA) according the usage instruction. The conditions including the value of PMT voltage and the gate were remained consistently during whole test. The reanalysis of some data was performed on Flowjo software (BD Biosciences).
Flowcytometry assay for splenic CD4 and CD8 T cell subsets
After the splenic cell suspension was made and adjusted into 1 × 107/mL by PBS, and the liquid of 100 μL was added into the flow tube. Then FITC-Hamster Anti-Mouse CD3ɛ (Biolegend), PE-Hamster Anti-Mouse CD4 (eBioscience, San Diego, CA) and PerCP-Hamster Anti-Mouse CD8a (Biolegend) were mixed in a volume ratio of 1:1.25:1.25 and 3.5 μL mixed antibodies were added into every tube, and the tubes were incubated in dark room under room temperature for 30 min. After centrifugation with 1,500 rpm for 5 min and washing three times with PBS, 300 μL PBS was added into the tube and the cells were suspended again. During these procedures, one tube of blank control without any antibody and three tubes of single positive control with different fluorochrome-conjugated antibodies were separately set at the same time. During the FCM operation, the gate and value of PMT voltage that was used to separate the lymphocytes group from other cells was set by the scatter properties of the cell plots scaled under FSC and SSC. Then, the cells with FITC and PE fluorochromes at the same time were calculated as CD3+/CD4+ T lymphocytes, and the cells with FITC and PerCP fluorochromes at the same time were calculated as CD3+/CD8+ T lymphocytes (Supplementary Data S4B and Fig. 4A). All of these parameters remained in the same station during the complete FCM operation. The count percentages of CD3+CD4+ and CD3+CD8+ T lymphocytes in different groups were tested and recorded at last.
Statistical analysis
For most figures, aggregated data are presented by using mean values to represent the central tendency, and standard error of the mean (SEM) to represent variability. All data were expressed as the mean ± standard error and analyzed by using SPSS 19 software (IBM, New York, NY). The procedures of one-way ANOVA or two-way ANOVA were performed for the difference analysis within multiple groups for corresponding data, and LSD was performed for the difference analysis between every two groups. The p-values obtained by these methods were considered as statistical significance if they were ≤0.05.
Results
hBD2 promotes BMDCs maturation by increasing MHC II and CD86 expression
To determine the effect of hBD2 on the differentiation and maturation of BMDCs, the growth of BMDCs cultured with or without hBD2 was observed daily under an inverted microscope. The round bone marrow leukocytes were densely seeded in a glass petri dish at day 1. A quantity of dot-shaped cell colonies sparsely distributed along the surface was found at day 2. Till day 5, the cell colonies became clusters and gradually dislodged from the bottom of the glass petri dish. Many typical DCs were floating in the culture medium and exhibited increased cell size, and cell nucleuses were visible at day 6 (Supplementary Data S2A). The spike-shaped projections were also seen clearly around the edges of cells. The large cell clusters at day 6 were collected for identifying the surface markers by FCM. High expression of CD11c (89.1%) and CD80 (95.7%) was exhibited on the DCs surface, whereas low expression of MHC II (54.2%) and CD86 (35.1%) classified the isolated cells into immature DCs (iDCs) (Fig. 2A).
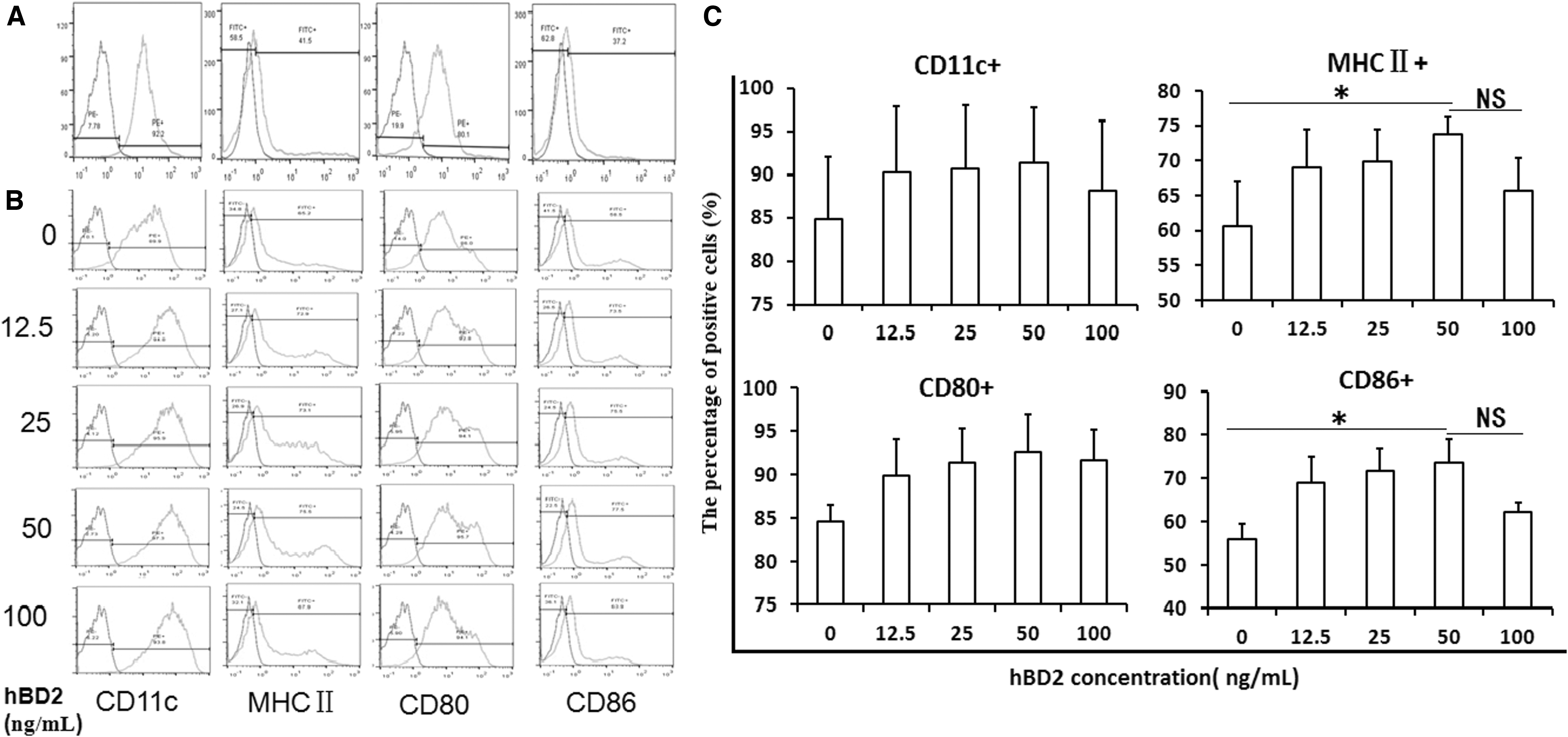
hBD2 promotes BMDCs maturation by increasing MHC II and CD86 expression. BMDCs were treated with rmGM-CSF and rmIL-4, and the intensity of CD11c, CD80, MHC II, and CD86 on the surface of BMDCs was tested by FCM. The antibodies against CD16(IgG FcR III)/CD32(IgG FcR II) were used to remove the possible nonspecific fluorescence given by an interaction between the Fc fragment of fluorescence-labeled antibodies and Fcγ receptor on the surface of BMDCs. Blank control was used to determine background levels of staining and to adjust PMT voltage value as a gating strategy.
After BMDCs were maintained with different concentrations, including 0, 12.5, 25, 50, and 100 ng/mL of hBD2 for another 24 h, the cellular percentages of CD11c+, MHC II +, CD80+, and CD86+ were obviously increased. Among the different concentrations of hBD2, 50 ng/mL was considered the optimum concentration for implementation. At this concentration, the highest expression signals of CD11c+ (91.32% ± 6.45%), MHC II+ (73.75% ± 2.47%), CD80+ (92.65% ± 4.31%), and CD86+ (73.81% ± 5.20%) were recorded; especially the percentages of CD86+, MHC II+ had significant meaning compared with the samples without hBD2 (p < 0.05). At 100 ng/mL hBD2, most of the marker signals were reduced (Fig. 2B, C). The gate scheme for FCM analysis of CD11c+, MHC II +, CD80+, and CD86+ is presented in Supplementary Data S2B.
hBD2 increases IL-6, IL-12B, and IFN-γ expression in BMDCs in vitro and in Balb/c mice
At the same time of the measure, the typical molecules related with BMDCs maturation on the surface of BMDCs, mRNA level of IL-6 and IL-12 were also tested by real-time PCR to measure whether the function of cytokines secretion of BMDCs was improved by hBD2. The results disclosed that gene expression of IL-6 was steadily upregulated as hBD2 concentration increased. The relative quantitation (RQ) of IL-6 mRNA culminated in 3.16 ± 0.1 at 100 ng/mL, which showed statistically significant differences when opposed to 0 ng/mL (p < 0.01) (Fig. 3A). Fifty nanograms/mL was the optimum concentration for the gene expression of IL-12B, and at this concentration RQ (2.99 ± 0.18) showed statistically significant differences when compared with 0 ng/mL (p < 0.01) (Fig. 3A).
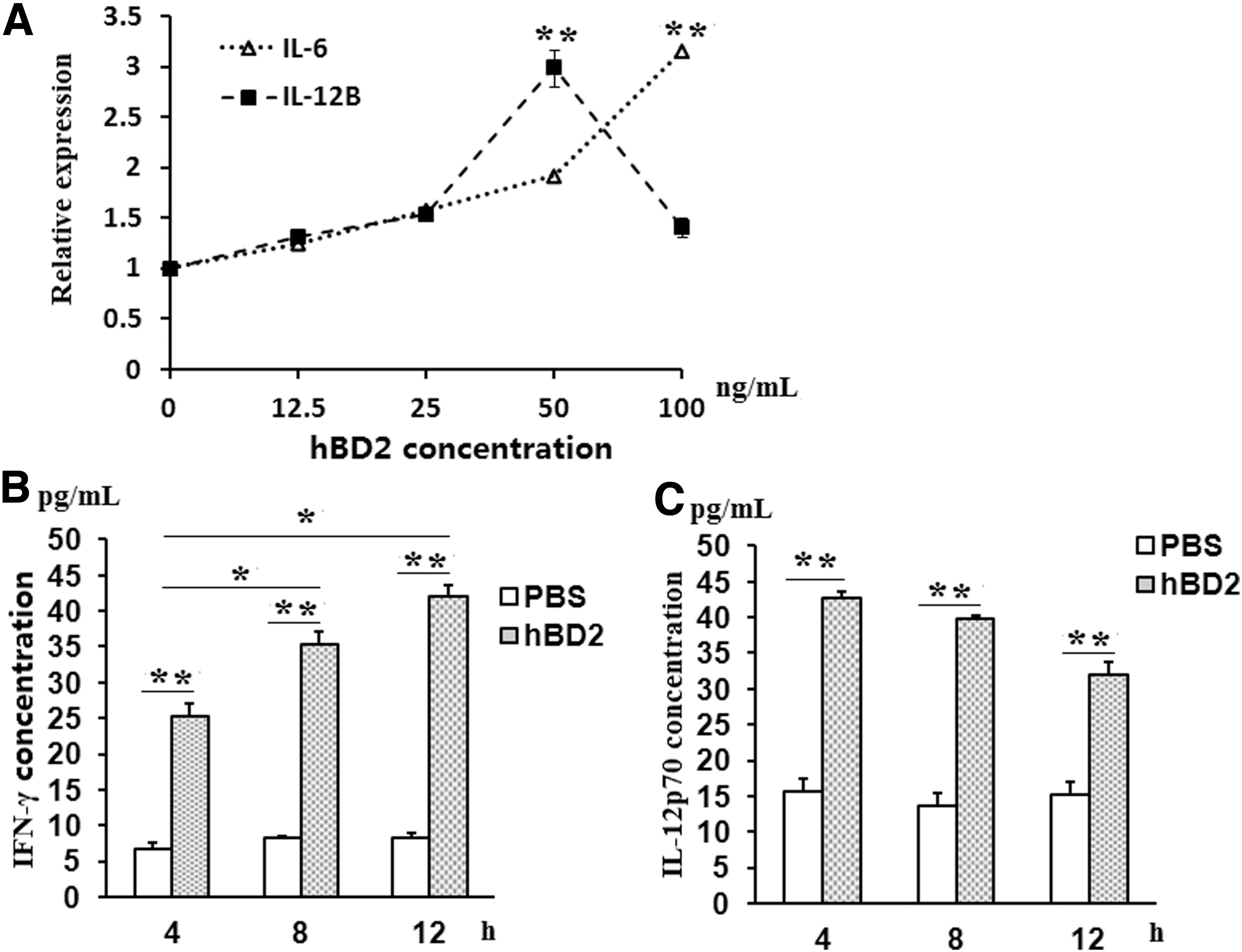
hBD2 upregulates the gene expression of IL-6, IL-12, and IFN-γ in Balb/c mice.
To determine the effect of hBD2 on cytokine expression in the body, IL-12p70 and IFN-γ expression in serum at 4, 8, and 12 h was tested by ELISA in Balb/c mice treated with 300 ng hBD2. The results showed that with the time extension at a 4 h interval, the serum concentrations of IFN-γ in the hBD2-treated group were increasing, and its value at 12 h was significantly higher than that at 4 and 8 h (p < 0.05), respectively (Fig. 3B). Although the concentrations of IL-12p70 in the hBD2-treated group were decreasing with the time extension, no significant differences were found among the time points of 4, 8, and 12 h (p > 0.05) (Fig. 3C). However, the concentrations of IFN-γ and IL-12p70 in PBS groups remained at similar levels at different time points and were statistically lower than hBD2 groups at every corresponding time point (p < 0.05) (Fig. 3B, C and Supplementary Data S3).
hBD2 enhances hepatitis B vaccine-induced total immunoglobulin and IgG2a level of HBsAb
With the aim of testing the function of hBD2 on humoral immunity, 2 weeks after the final immunization the sera levels of HBsAb and its levels of IgG1 and IgG2a subclasses were tested by ELISA. The HBsAb titers increased after immunization and they reached a peak at the 42nd day in the hepatitis B vaccine group and the combined group (hepatitis B vaccine + hBD2); the values of HBsAb titer (S/CO) from the mice in the combined group at days 11, 22, 33, and 42 were significantly higher than that from the hepatitis B vaccine group at the same day post-immunizations, respectively (p < 0.01) (Fig. 4A and Supplementary Data S5).
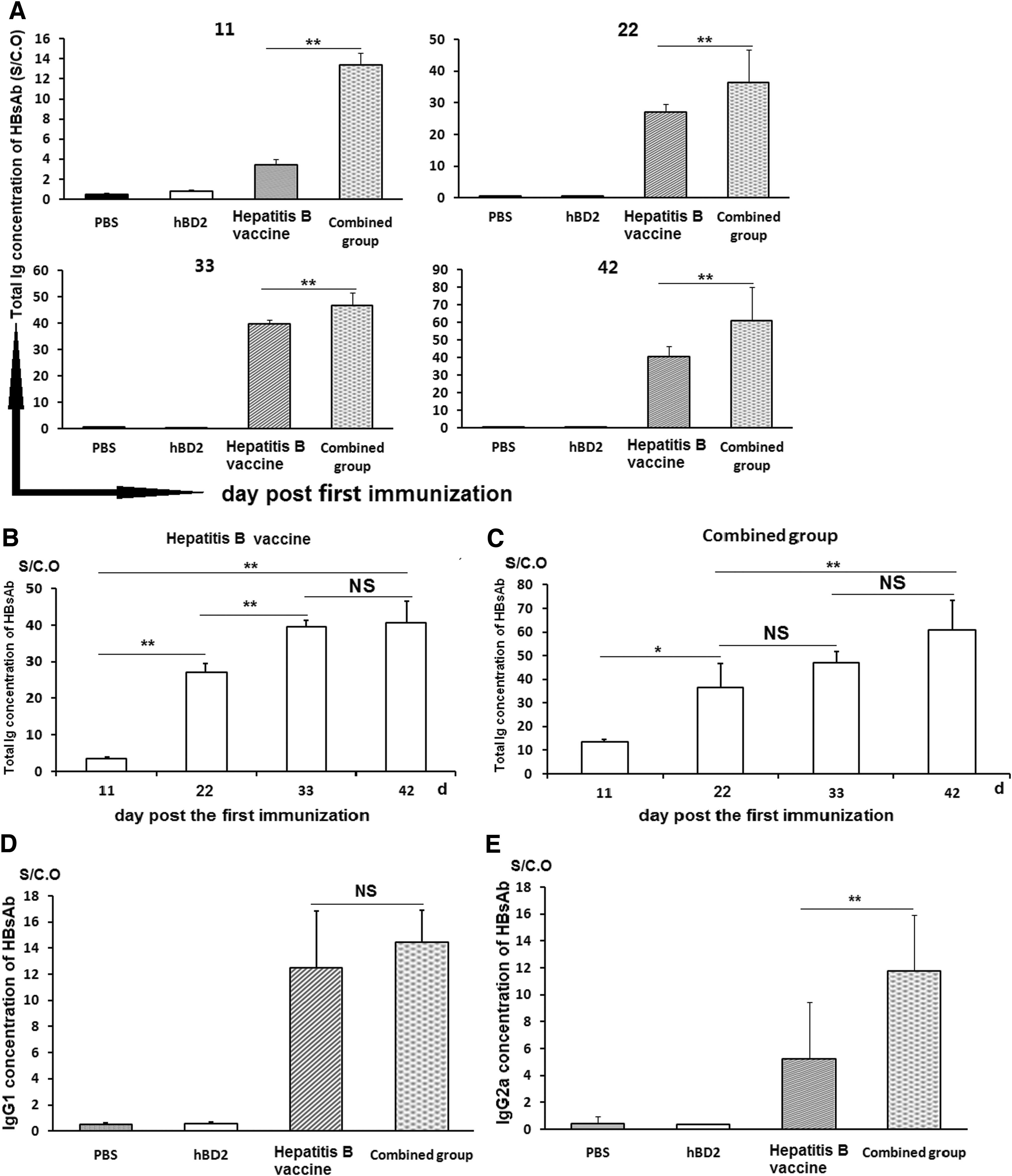
hBD2 shows higher efficiency in assisting hepatitis B vaccine to generate HBsAb immunoglobulin and IgG2a subclass. After immunization, total HBsAb in serum in each group was tested by double antigen sandwich ELISA, and the levels of IgG1 and IgG2a subclass were checked by indirect ELISA.
Moreover, the results showed that HBsAb level in the sera from the combined group increased more quickly than the hepatitis B vaccine group (Fig. 4A). For HBsAb titers in the mice from the hepatitis B vaccine group, except that there was no significant difference between the values measured on the 33rd and 42nd day after the first immunization (p > 0.05), there was a constant significant difference between any other two time points (p < 0.01) (Fig. 4B). However, for the combined group, there were no statistically significant differences between the data from the 33rd and 42nd day (p > 0.05), and the data from the 33rd and 22nd day post the first immunization as well (p > 0.05) (Fig. 4C). Significant differences were found among the time points on the 11th, 22nd, and 42nd days, post the first immunization (p < 0.01) (Fig. 4C).
Because different subclasses of immunoglobulin have different immunological functions, the quantities of IgG1 and IgG2a subclasses of HBsAb in serum from the hepatitis B vaccine group and the combined group at the 42nd day were tested by indirect ELISA. IgG2a concentration in the combined group was significantly higher than that from the hepatitis B vaccine group (p < 0.01) (Fig. 4E). However, IgG1 values between the hepatitis B vaccine group and the combined group had no statistically significant difference (p > 0.05) (Fig. 4D).
hBD2 increased the count percentage of CD3+CD4+ and CD3+CD8+ T cell subclasses in mice spleen
To measure the ability of hBD2 to improve T cell proliferation and differentiation, the percentages of spleen CD3+CD4+ and CD3+CD4+ T lymphocytes were tested by FCM in hepatitis B vaccine immunized Balb/c mice with or without hBD2 injection as described earlier. The results showed that the percentages of spleen CD3+CD4+ T lymphocytes in the hBD2 group, hepatitis B vaccine group, and combined group were close to each other according to the FCM assay and showed no striking difference among them (p > 0.05), but they were significantly higher than that from the PBS group (p < 0.05) (Fig. 5A, B). However, the percentage of CD3+CD8+ in the hBD2 group was significantly higher than those in the other groups (p < 0.05), and there was no significant difference between the hepatitis B vaccine group and the combined group (p > 0.05) (Fig. 5A, C). These results have some implications that hBD2 could increase the percentage of CD3+CD8+ and CD4+/CD8+ T lymphocytes in Balb/c mice. But this possibility needs more research to be verified in future.
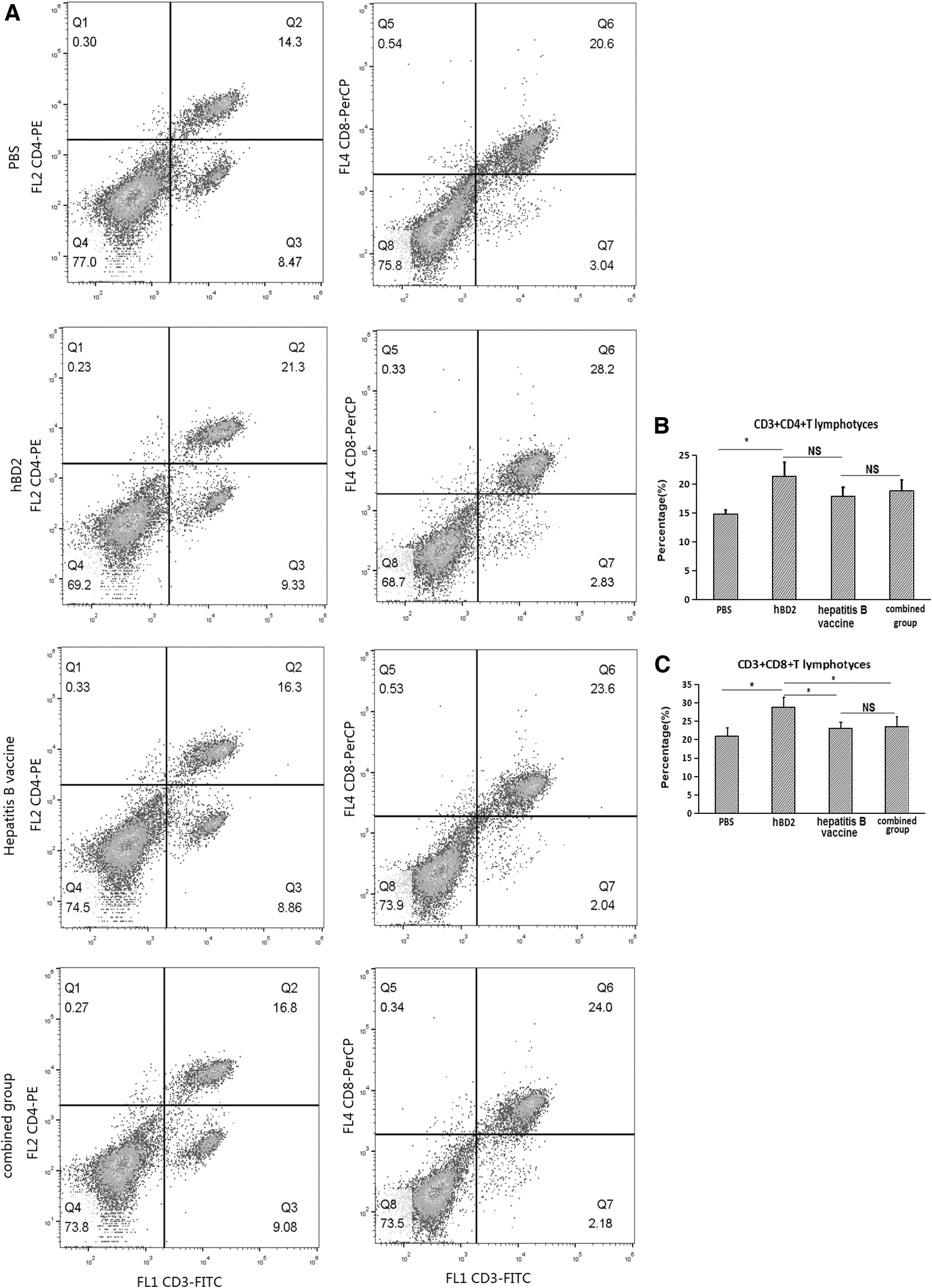
hBD2 improves the percentage of CD3+CD8+ and CD4+/CD8+ T lymphocytes in Balb/c mice. After three times immunization with PBS, hBD2, hepatitis B vaccine, or hepatitis B vaccine and hBD2 (combined group) with a 2 week interval, all mice were sacrificed on the 14th day post last injection as described earlier according to experimental design. The spleen was removed, splenocytes suspension was prepared in a clean bench, and the splenetic monocyte suspension was prepared by the PBMC method. Then, a monocyte suspension was used to test the T cell subsets assay on flow cytometery.
hBD2 enhances IL-2, IL-12p70, and IFN-γ expression in Balb/c mice with hepatitis B vaccine immunization
To measure whether the effect of hBD2 on hepatitis B vaccine improves the biological function of cellular immunity, mRNA levels of IL-2, IL-12B, and IFN-γ in splenocytes were tested by real-time PCR and the concentrations of IL-2, IL-12p70, and IFN-γ in serum by ELISA were tested in the mice with different schedules of immunization as described earlier. The RQs of IL-12B (Fig. 6B) and IL-2 (Fig. 6C) in the combined group were higher than in any other groups (p < 0.05). The RQ of IFN-γ in the combined group was higher than in the PBS and hepatitis B vaccine groups (p < 0.05) but near to the hBD2 group (p > 0.05) (Fig. 6A). Moreover, RQs of IFN-γ, IL-12B, and IL-2 in the hBD2 group were higher than in the hepatitis B vaccine groups (p < 0.05) (Fig. 6A–C). Surprisingly, there were no differences in the RQs of IFN-γ, IL-12B, and IL-2 between the hepatitis B vaccine group and the PBS group (p > 0.05) (Fig. 6A–C).
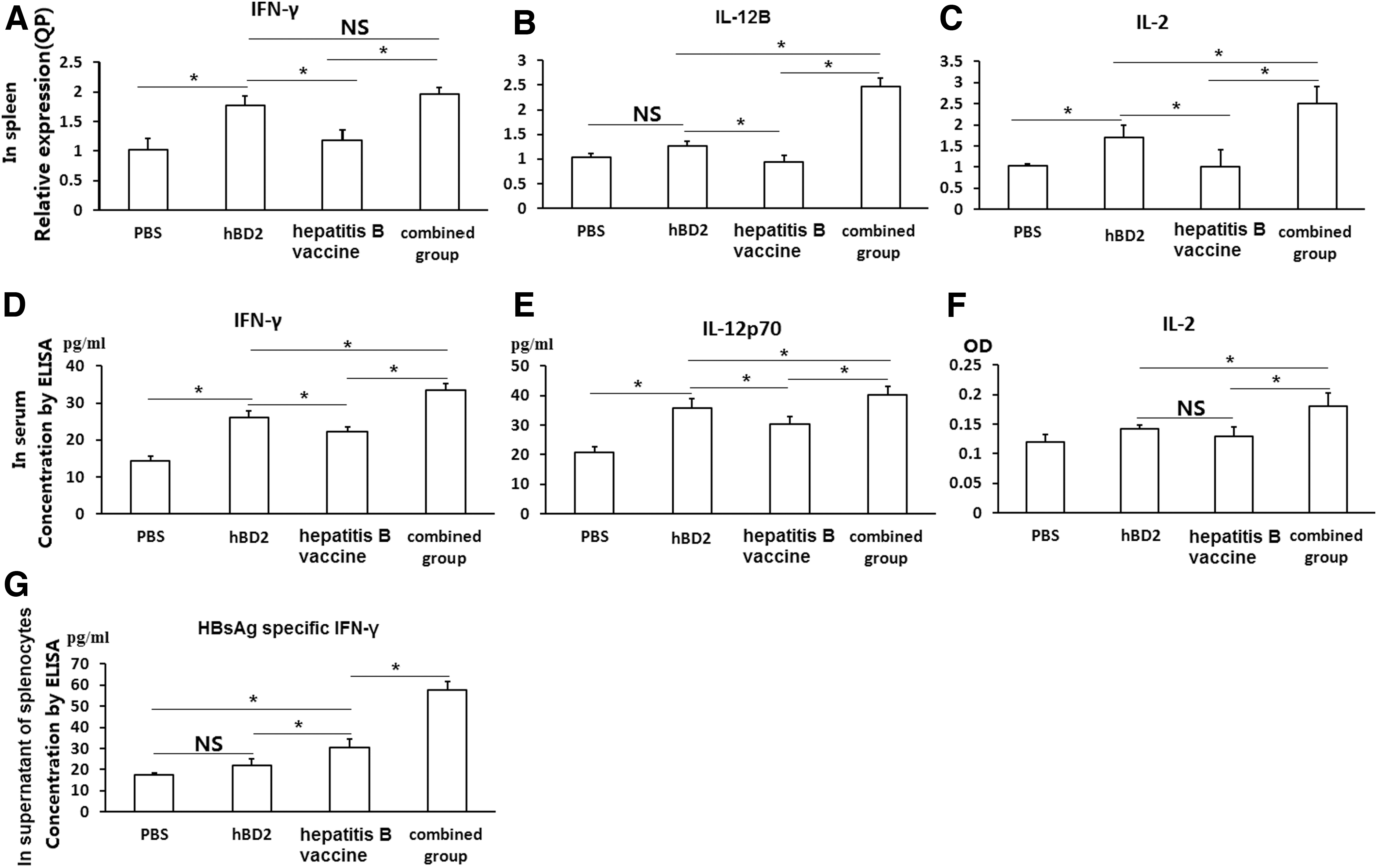
hBD2 enhances IL-2, IL-12p70, and IFN-γ expression in Balb/c mice with hepatitis B vaccine immunization. Two weeks after three times immunization with PBS, hBD2, hepatitis B vaccine, and combined group (hepatitis B vaccine + hBD2), as described earlier, respectively, the serum and spleen cells were collected. The mRNA levels of IL-2, IL-12B, and IFN-γ in spleen cells, and IL-2, IL-12p70, and IFN-γ in serum were tested; hepatitis B vaccine-induced specific IFN-γ in supernatants of the splenetic cells was stimulated with HBsAg for another 18h in vitro, as well.
In addition, the sera levels of IL-2, IL-12p70, and IFN-γ in the PBS group were significantly lower than the levels in the other three groups (p < 0.05), rather than IL-2 in the hepatitis B vaccine group (Fig. 6D–F). Further, the combined group was the most productive group, and the hepatitis B vaccine group produced less IL-12p70 and IFN-γ than the hBD2 group (p < 0.05) (Fig. 6D–F).
hBD2 enhanced HBsAg-specific IFN-γ expression in splenic cells
Because IFN-γ is a very important cytokine that enhances Th1 type immunity, and to explore the effect of hBD2 in the stimulation of antigen-specific cytokine expression, hepatitis B vaccine-induced specific IFN-γ in splenic cells from the mice with three times different immunization plans as described earlier was further tested by double antigen sandwich ELISA. After the splenic cells harvested from the mice with different schemes of immunization were stimulated with 0.2 μg/mL HBsAg for 18h in vitro, the IFN-γ concentration in splenic mononuclear cell supernatants in the PBS group (17.57 ± 0.93 pg/mL) was statistically lower than that in the hepatitis B vaccine group (30.24 ± 4.03 pg/mL) and the combined group (57.74 ± 3.86 pg/mL) (p < 0.05), but not the hBD2 group (21.92 ± 2.97 pg/mL) (p > 0.05). Further, IFN-γ concentration in the combined group was also significantly higher than in the hepatitis B vaccine group (p < 0.05), which exhibited statistically significant difference compared with the hBD2 group and the PBS group (p < 0.05) (Fig. 6G).
Discussion
This study reported for the first time that hBD2 attempted to improve the immune efficiency of the hepatitis B vaccine in Balb/c mice, and contributes to BMDCs maturation, which implies that hBD2 could be a better alternative for the hepatitis B vaccine containing an alum adjuvant that often induces Th2 type immune responses (5). Usually, mature DCs enhance the immune response and iDCs induce immune tolerance, so the situation of mature DC is an important influential factor for immune response. This article reported that aluminum could not promote classical DC maturation (41). On the contrary, hBD2, a naturally biological and antimicrobial peptide, is confirmed to have the function of interacting with iDCs via CCR6 (29) and iDCs migrate from human bone marrow to the antigen location. So, the effect of hBD2 on BMDC maturation from Balb/c mice was tested by FCM at first in this article.
CD11c is a typical marker for both human and mice DCs (52). Although co-stimulatory molecules (CD80, CD86, and CD40) and antigen-presenting molecule (MHC-II) are already expressed in iDCs, their expression intensities are greatly enhanced after the DC maturation and activation (31). In our study, the cell percentage of CD11c+, MHC II+, CD80+, and CD86+ of BMDCs was upgraded to 91.32%, 73.75%, 92.65%, and 73.81% at the treatment of 50 ng/mL hBD2, respectively (Fig. 2B, C). And the spike-shaped projections around the edges of DCs were obvious. The increased CD86 on BMDCs by hBD2 in this article agreed with the document (26), which reported that hBD2 showed the same effect on THP-1 cells. These evidences indicate that hBD2 has the ability to promote BMDCs activation and maturation. A similar result was also reported by other researches (25,46) and mouse β-defensin-14 (59).
The cytokine pattern is an important sign for evaluating the DC activation. Cytokines such as IL-6, IL-12, and IFN-γ are not only helpful for the main switch of immune type from Th0 to Th1, but also beneficial for the symbols of activation effectors for innate and adaptive immunities. To explore the function of hBD2 in cytokine expression by DC, the levels of mRNA and protein for IL-6 and IL-12 in BMDC stimulated by hBD2 were examined by qPCR and ELISA in this article. The results showed that the gene expression of IL-6 and IL-12B in BMDCs ascended with addition of hBD2 (Fig. 3A). Beyond the promotion of cytokine gene expression in BMDCs in vitro, IL-12p70 concentration in sera reached the topmost expression at 4 h and then declined with the incubation time extension, but their levels were still higher than in the PBS-treated group (Fig. 3C). However, the level of IFN-γ in sera was elevated with the incubation time extension (Fig. 3B). IL-12p70, composed by IL-12p35 and IL-12p40, is the only active form of IL-12 (11). More IL-12p70 secretion is not only characteristic of mature DCs, but it also contributes to inducing the development of Th0 to Th1 and initiating specific cellular immunity (11). So, the increase of IL-12p70 in this article verified that hBD2 promotes murine DC maturation. A similar result is also found in human DCs (17).
To further explore the immune effect of hBD2 on humoral and cellular immune responses as adjuvant and compare the immune effects of hBD2 and aluminum adjuvant, which has been widely used in many vaccines, including the hepatitis B vaccine, hBD2 was conjugated with the hepatitis B vaccine and deployed in Balb/c mice. The HBsAb concentration and cytokines, including IL-2, IL-12p70, and IFN-γ in serum, and HBsAg-specific IFN-γ in splenic cells stimulated by hepatitis B vaccine in vivo were tested by ELISA. The results showed that mRNA levels of IL-2, IL-12B, and IFN-γ in splenocytes and the concentrations of IL-2, IL-12p70, and IFN-γ in sera in the combined group were consistently and significantly higher than in the hepatitis B vaccine group (Fig. 6A–F). However, the alum has reported the inhibitory effect of IL-2 and TNF-a in cultured human T lymphocytes (54,63) and chicken T lymphocytes (53,63) treated by AlCl3 in vivo. It is well known that IL-2 is a T cell-derived cytokine that serves as a cofactor for T lymphocytes growth and natural killer cells differentiation (40). The increased IL-2 in the combined group implied that hBD2 might interleave into the stage of T cell proliferation and innate immunity. In addition, IL-12 and IFN-γ expression also confirmed the enhancement of cellular immune response as a better adjuvant for hepatitis B vaccine. Except for these, the higher concentrations of HBsAg-specific IFN-γ were found in the combined group than the hepatitis B vaccine group (Fig. 6G). The result that hBD2 increases IFN-γ expression was also supported by the research by using hBD2 as an adjuvant to improve the immune protective effect of the receptor-binding domain of spike protein from Middle East respiratory syndrome-coronavirus (MERS-CoV) in mice (25). These findings might imply that hBD2 could greatly strengthen and magnify the specific cellular immune response to hepatitis B vaccine to some extent.
Except the aforementioned cellular immunity provoked by hBD2, our results presented that the percentages of CD3+CD4+ T lymphocytes in the hBD2 group, hepatitis B vaccine group, and combined group were significantly higher than that in the PBS group, but there were no statistically significant differences among the first three groups. On the other hand, the percentage of CD3+CD8+ T cells in the hBD2 group was statistically higher than values from the PBS group, hepatitis B vaccine group, and combined group, respectively, but there was no statistically significant differences between the last three groups (Fig. 5). In general, an exogenous antigen would activate CD3+CD4+ T lymphocytes by the MHC II antigen presentation pathway and an endogenous antigen would activate CD3+CD8+ T lymphocytes by the MHC I antigen presentation pathway. This could explain well that the percentage of CD3+CD4+ T lymphocytes, not CD3+CD8+ T lymphocytes in the hepatitis B vaccine group and the combined group was significantly increased, compared with the PBS group, because both hepatitis B vaccine and hBD2 were administered in an exogenous way in our experiment. And it might also be the main reason for non-significant difference in the percentage of CD3+CD8+ T cells between the combined group and the hepatitis B vaccine group.
Regarding the result of meaningless difference in the percentage of CD3+CD4+ T cells between the combined group and the hepatitis B vaccine group (Fig. 5), there are variable possibilities. First, the hepatitis B vaccine used in our experiment is a commercial product and contains both HBsAg and alum adjuvant, and the latter is widely believed to be lightly toxic to lymphocytes in some circumstances, which could decrease the number of lymphocytes (20,43,57,58,63) and does not elevate the proportion of CD3+CD4+ T lymphocytes and CD3+CD8+ T lymphocytes in Balb/c mice (5). Compared with the higher percentages of CD3+CD4+ T lymphocytes and CD3+CD8+ T lymphocytes in mice treated with hBD2 alone, the real improvement effect of hBD2 in the combined group in our experiment might be partially discounted by alum contained in the hepatitis B vaccine. Second, it is well known that the immune response of immunization may be influenced by many factors, including the characteristics and the dosage of immunogenic substances and adjuvant, inherent in the subject, the manner and the schedule of immunization, and so on. However, the imperfect results of the percentages of CD3+CD4+ T lymphocytes and CD3+CD8+ T lymphocytes in spleen in the combined group do not contradict the opinion that hBD2 exhibits a stronger function by pushing the cellular immune response as an adjuvant. Further, a low percentage may not exactly be equal to the smaller group and poorer immune function. Because the hepatitis B vaccine and hBD2 have different structures and biological functions, hBD2 has a very good function of increasing the proportion of lymphocytes in this study, but due to the lack of specific antigen stimulation, these lymphocytes may not be exactly functionally activated. This speculation may be verified from this study that the mice in the hBD2 group have the highest proportion of lymphocytes, whereas the mice in the combined group have the strongest ability of HBsAb production (Fig. 4) and cytokine secretion, including IL-2, IL-12, IFN-γ, and HBsAg-specific IFN-γ (Fig. 6G), and the highest expression density of MHC II molecules on the PBMC (Supplementary Data S4) in our experiment. These novel increasing patterns in mice treated by hepatitis B vaccine plus hBD2 convinced us to some extent that hBD2 has the exact potency of an immune enhancer on the hepatitis B vaccine. The view that hBD2 has good immune adjuvant function is also supported by a similar effect of mouse β-defensin-2 on influenza virus DNA vaccine (49).
In line with the cellular responses, our study suggests that hBD2 greatly enhanced robust humoral immunity in hepatitis B vaccine immunized Balb/c mice. It is noticeable that hBD2 in the combined group increased HBsAb titer about 20% and especially IgG2a subclass, compared with the hepatitis B vaccine group (Fig. 4A, D and E). IgG1/IgG2a ratio in the combined group and the hepatitis B vaccine group in our study (1.84 and 3.61, respectively) showed that hBD2 was more beneficial to promoting Th1 type immunity than Th2-biased isotype profile, which was reported to be elicited usually by both Al(OH)3 and AlPO4 (IgG1/IgG2a ratio of >50) (28). Interestingly, the immune dosage for mice in the hepatitis B vaccine group by using the volume of hepatitis B vaccine containing an equivalent to 3 μg HBsAg in our article was very close to that of 2 μg HBsAg plus 100 μg aluminum salt reported in the literature, and their results of ratios of IgG2a to IgG1 were also very close (5). These data confirmed that hBD2 promotes the Th1-biased isotype profile of immunoglobulin.
Although the recombinant hBD2 used in our experiment was expressed in engineered Escherichia coli, this might possibly bring some contaminated LPS into the experiment and cause some biases in the conclusion. The result of higher IgG2a rejected that suspicion to some extent, because LPS in mice usually leads to more IgM or IgG2b expression. So, on the contrary, our result holds that the immune effects in our experiment come from hBD2, not the contaminant LPS. A similar conclusion, that hBD2 strengthens humoral immunity, was also verified by other publications (15,25,33). Further, the serum concentration of HBsAb on the 11th day post the first vaccination by hepatitis B vaccine along with hBD2 was significantly higher than that with hepatitis B vaccine immunization alone; the value of HBsAb concentration on the 7th day post the second immunization in the combined group was almost equal to the titer of HBsAb on the 5th day post the third immunization in the hepatitis B vaccine group (Fig. 4A). These data prove that hBD2 strongly improves the humoral immune response of hepatitis B vaccine in Balb/c mice.
HBD-2 expression is remarkably inducible in keratinocytes or macrophages in skin, oral mucosa, lung, and other sites, in response to LPS or inflammatory cytokines, including IL-1β, IFN-γ, and TNF-α (21). Although great controversy still exists as to the real receptors of hBD2 in humans, and its mechanism of immune regulation is not totally clear, it is widely accepted that hBD2 may chemoattract iDCs and memory T lymphocytes, as well as TNF-α activated neutrophils (35) and epithelial cells (51), and trigger a robust cytokine expression from PBMCs (7) by binding with CCR6 (56) or CCR2 (26,39). By acting as an endogenous ligand for the Toll-like receptor 4 (TLR4), hBD2 may induce the expression of costimulatory molecules, including CD80/CD86, and lead to the activation and maturation of iDCs, resulting in an enhancement in antigen-specific immune responses (6).
HBD-2 is also believed to have the function of T cell activation by MEK/ERK, JNK, and PI3K/Akt pathways (22). And the activation of these pathways finally leads to the activation of transcription factors, including NF-κB, AP-1, NF-AT, Ets, C/EBP, CREB, or Sp1 in T cells; increases the expression of cytokines, including IFN-γ, IL-1β, TNF-α, IL-6, and IL-22; and inhibits IL-17 production via SOCS3 or by improving IL-10 generation (22). hBD2 owns the ability to enhance the primary innate and effective adaptive immunity by the CCR2-mediated Nod2 signaling pathway under the stimulation of hBD2-conjugated antigen (21). On the other hand, hBD2 also (30) has the ability to act rapidly and directly on APCs and lead to differential binding and cellular uptake. And sub-cytotoxic concentrations of hBD2 may also trigger a rapid release of cytoplasmic vesicle content in iDCs. However, the inhibition activity of hBD2 on the classical pathway of the complement system suggests a protective role against its uncontrolled activation (4). hBD2 is also reported to have the function to activate mast cells and lead to degranulation and release of histamine and prostaglandins (34,45), thus favoring the recruitment of neutrophils to the inflammatory site. These effects of hBD2, undoubtedly, characterize its better features as a good adjuvant and immune regulatory agent by being beneficial for antigen presentation and immune activation.
It seems unreasonable that hBD2 is used as an adjuvant lining with hepatitis B vaccine used in BALB/c mice. It is a really difficult task to fully explain the effects and mechanism of the immune regulatory role of hBD2 in immune regulation in mice, but some preliminary analysis and reported related studies might give us more confidence on this conclusion. First, the protein sequence of human CCR6 (query ID: NP_004358.2), which is believed to be a receptor of hBD2 and expressed on human DC and lots of other immune cells, shares 74.59% of percent identity value and 98% of query coverage number with mouse CCR6 (sequence ID: NP_001177267.1), according to a report by BLAST on NCBI net. These data imply that both human CCR6 and mouse CCR6 have high similarity to the sequence and even the structure and they possess similar immune functions by binding with hBD2. An exactly similar chemotactic activity of hBD2 and its mouse ortholog, named β-defensin 4, in a CCR6- and CCR2-dependent manner in C57BL/6 mice was reported by Röhrl et al. (39), in 2010. In line with this conclusion, a similar effect of mBD4 on immune regulation in mice was reported in 2000 (21). Similarly, Zhao et al. reported that the liposome-mediated hBD2 plasmid DNA transfection system was beyond the species and appeared to be a promising method for treating urinary tract infection in a rat (60). Enhanced immune therapy effect of melanoma in mice by hBD2 was reported by Sun et al. (46) in 2019. All of these imply that hBD2 not only plays a very important immune regulatory function in humans, but it also plays a vital function in these broad pattern effects in many other species.
Undoubtedly, there were some limitations to this article. Because a limited volume of serum could be extracted from one mouse, lots of typical cytokines, including IL-10, TGF-β, and IL-17, and other subclasses of immunoglobulin, including IgM, IgG2b, IgG3, and IgG4, were not included in our experiment. And for evaluation of antigen-specific T cell function, the article just focused on the expression of HBsAg-specific IFN-γ in the spleen, and no cytotoxic function test of CD8+ T lymphocytes was emphasized. The limitations of this study also include that the neutralization effect of an antibody and anti-HBV infection ability were not evaluated. These shortcomings may have a certain impact on the evaluation of the whole functions of hBD2, but all of these will not negate its overall effect absolutely.
In summary, our results suggested that hBD2 triggered specific cellular and humoral immunities by the induction of DC maturation in Balb/c mice when it was used as an adjuvant in hepatitis B vaccine.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The work was supported by grants from the National Natural Science Foundation of China (NO.30760280, 81760472), and the Science and Technology Support Program of Jiangxi Province, China (No.2007BS12802, 20142BBG70044). Funding was also provided by the Natural Science Funding of Jiangxi province, China (No. 20181BAB205064), and the Educational Department Foundation of Jiangxi Province, China (No. GJJ08126, GJJ10060).
Supplementary Material
Supplementary Data S1
Supplementary Data S2
Supplementary Data S3
Supplementary Data S4
Supplementary Data S5
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.