
Abstract
Bovine herpesvirus-1 (BoHV-1) uses many mechanisms to elude the immune system; one of them is spreading intracellularly, even in the presence of specific antiviral antibodies. Cytotoxic T lymphocytes (CTLs) are necessary to eliminate the virus. The main preventive strategy is vaccination based on inactivated virus. These vaccines are poor inducers of cellular immune responses, and complicate serological diagnosis and determination of the real prevalence of infection. DNA vaccines are a good option because of the capacity of Differentiating Infected from Vaccinated Animals—(DIVA vaccine)—and may be the best way to induce cytotoxic responses. Although this type of vaccines leads to only weak “in vivo” expression and poor immune responses, incorporation of molecular and/or chemical adjuvants can improve the latter, both in magnitude and in direction. In this study, we have investigated the specific immune responses elicited in mice by DNA vaccines based on the BoHV-1 glycoprotein D (pCIgD) with and without two different adjuvants: a plasmid encoding for murine CD40L (pCD40L) or Montanide™ 1113101PR (101). Mice vaccinated with pCIgD+CD40L, pCIgD+101, and pCIgD+CD40L+101 developed significantly higher specific antibody titers against BoHV-1 than the pCIgD group (p < 0.01). The animals vaccinated with pCgD+pCD40L+101 raised significantly higher levels of IgG2a and IgG2b (p < 0.01 and p < 0.001, respectively) than mice vaccinated with pCIgD alone. On the contrary, when the activity of CTL against cells infected with BoHV-1 was measured, the vaccine pCgD+pCD40L+101 induced significantly higher levels of cytotoxicity activity (p < 0.001) than pCIgD alone. A significant increase in the CD4+ populations in the group receiving pCIgD+CD40L+101 in comparison with the pCIgD group was observed and, also, interferon gamma, interleukin (IL)-6, and IL-17A levels were higher. Considering the results obtained from this study for humoral and cellular responses in mice, the inclusion of pCD40L and 101 as adjuvants in a BoHV-1 DNA vaccine for cattle is highly recommendable.
Introduction
Bovine herpesvirus-1 (BoHV-1) is an important viral pathogen of cattle, and it produces a variety of diseases, including vulvovaginitis/balanoposthitis, infectious bovine rhinotracheitis, systemic infection in the newborn, and abortion in cows (35,73). The economic impact of this infection related to weight loss, decreased milk production, and abortion. More importantly, infected animals that are immunosuppressed may suffer secondary bacterial infections increasing morbidity and mortality (21,36).
However, if infection or reactivation of the virus occur during pregnancy, abortions can occur (54,59,70). Six European countries became BoHV-1 free after the introduction of expensive eradication programs, consisting of seropositive animals culling (62,71).
A more common way to prevent BoHV-1-related diseases is the use of inactivated or attenuated live vaccines. However, they cannot control latent infection. The ability to differentiate infected from vaccinated animals (DIVA) is an important issue (65). Live-attenuated vaccines may cause abortion, latency with reactivation and shedding (79,80). Also, they may downregulate major histocompatibility complex (MHC) class I molecules (31,55), which compromise the subsequent cellular immune response. On the contrary, inactivated viral vaccines have a short duration of immunity and are poor inducers of cellular responses (17). Importantly, BoHV-1 can persist in the presence of antiviral-specific antibodies (24,38,52). So, cytotoxic T lymphocytes (CTLs) are necessary for controlling the virus (73). Novel vaccines against BoHV-1 that do not have the problems associated with current vaccines are thus necessary. They need to contain viral epitopes capable of inducing protective immune responses, and should ideally be suitable for DIVA.
DNA vaccines have potential to induce cell-mediated and humoral immune responses with the advantages of having good thermostability, the capability of coexpression of multiple antigens and reduced production costs since they avoid the need of working with viral particles (4,10,28). Licensed DNA vaccines exist for hematopoietic necrosis virus in salmon (Apex-IHN, Novartis) (26), West Nile virus in horses (West Nile Innovator, Fort Dodge) (15), and melanoma in dogs (Canine Melanoma Vaccine, Merial) (6).
Although genic vaccines have the capacity to induce cytotoxic responses (40), they have limited potency because they do not amplify or propagate “in vivo.” For this reason, it is necessary to increase the magnitude and direction of the immune response with the incorporation of molecular and chemical adjuvants.
Viral glycoproteins are involved in several steps of BoHV-1 replication (7,25,43,57). Glycoprotein D (gD) is responsible for the penetration of the BoHV-1 in cells with participation in the membrane fusion and viral adsorption (74). It induces neutralizing antibodies (3,13,17) and has cytotoxic epitopes (16,17). Notably, vaccination with a plasmid encoding the secreted form of the gD encoding in a vector produced a different immune response (type 1/type 2) than vaccination with one encoding full-length gD (41,42).
Injection of a naked plasmid vector into animal muscle or skin can induce an immune response against a transgene-encoded antigen (Ag) (60), but in general this response is weak and needs to be enhanced to provide protective immunity (23,49,61). The interaction between the CD40 receptor and its ligand, CD40L, regulates humoral and cellular immune responses. CD40L (or CD154) is a 39-kDa type II membrane glycoprotein with homology to TNF-α (2). This molecule is expressed transiently on CD4+ T cells within hours after ligation of the TCCRD 3 complex and then is rapidly downmodulated (66). CD40L allows the T lymphocyte to interact with B lymphocytes, monocytes, and dendritic cells (DCs), inducing their proliferation, differentiation, and expressing accessory molecules that facilitate the presentation of antigens to T lymphocytes (22). Given the ability of CD40L to enhance the development and proficiency of antigen-presenting cells (APCs), we hypothesized that coinjection of a DNA vaccine with a plasmid encoding
Mice have been used as an experimental model to select optimal formulations for BoHV-1 vaccines before carrying out experiments in large animal hosts (67).
We have previously shown in the mouse model that incorporation of adjuvants into a genetic vaccine against BoHV-1 is immunologically beneficial (18,40). Recently, we demonstrated that bovines vaccinated with a DNA vaccine coding BoHV-1 gD, adjuvanted with Montanide 1113101PR (101), induced better protection upon challenge than animals vaccinated with a genic vaccine alone. Although this vaccine was able to diminish clinical symptoms and viral excretion, our results showed that it is still necessary to induce a higher immune response against the virus (61).The 101 adjuvant is composed of non-crosslinked charged polymers (10–500 nm) dispersed in water with an immunostimulating compound. It has the capacity to improve DNA binding and transfection potency. Also, it can recruit cells to the site of injection (78).
In this study, we evaluate the immune response induced in mice by a DNA vaccine containing the truncated, secreted version of BoHV-1 gD (18), in combination with the adjuvants CD40L and 101, to extend our previous studies in the murine model.
Materials and Methods
Plasmid construction
Polymerase chain reaction (PCR) product encoding the secreted version of the BoHV-1 glycoprotein D (gD) was inserted into EcoRI-digested pCIneo vector (Promega) to construct plasmid pCIgD, prepared as described before (40). Plasmid pDNA3.1 contained the gene for murine CD40L (pCD40L) and was kindly provided by Dr. Kipps, University of California. Nonrecombinant plasmid pCIneo was used as control. All plasmid preparations had A260/280 ratios >1.7. After purification, plasmids were diluted with phosphate-buffered saline (PBS).
Adjuvant
Montanide 1113101PR (101) was provided by Seppic Inc., France, and used according to the manufacturer's instructions.
Toxicological tests made on Montanide adjuvants (derml irritation test, Oral LD 50, Berlin test, IP LD 50, pyrogenicity, ocular irritation test) can be considered nontoxic and well tolerated. The Committee for Veterinary Medical Products (CVMP) considered these adjuvants safe for use in immunological products. These substances have been authorized in the annex of the European Council Regulation n° 470/2009 (previously 2377/90/EC).
Generation of bone marrow-derived DCs
Bone marrow-derived DCs were obtained as previously described (39). In brief, the marrows of femurs and tibiae of naïve BALB/c mice (n = 3) were flushed out with RPMI 1640 medium. After washing, cells were diluted at 1 × 106 cells/mL in RPMI 1640 medium (Invitrogen Life Technologies) with 10% fetal calf serum (FCS), 5.5 × 10 − 5 of 2-mercaptoethanol (Sigma-Aldrich), and 30% conditioned medium from GM-CSF-producing NIH-3T3 cells. The cells were cultured for 9 days with 50% of the medium replaced every 2 days. On day 9 of culture, >85% of the harvested cells expressed CD40, CD11c, MHC class II, and CD80 but no Gr-1 (data not shown).
Endocytosis assay
DCs (produced from naïve mice) were incubated with medium, pCIneo (1 μg/mL), pCIgD (1 μg/mL), 101 adjuvant (ratio 24% adjuvant and 76% PBS), and pCIgD+101 (ratio 24% adjuvant and 76% DNA). Then, 100 μg/mL FITC-labeled OVA was added to RPMI 1640 medium and incubated for 40 min at 37°C. After incubation, the cells were washed with cold PBS three times, and evaluation of uptake was tested by flow cytometry. The fluorescence background in cells incubated with FITC-OVA at 4°C was determined and subtracted. The results are expressed as mean fluorescence intensity values.
Animal treatment and vaccine formulations
Male BALB/c mice (28 ± 2 g) from La Plata University (Argentina) were used. The animals were acclimated and randomly distributed into five experimental groups. Mice were kept in rodent caging with ventilated systems, and received water and food ad libitum. Handling and housing of animals were according to the guidances of the Institutional Committee for Care and Use of Experimental Animals, CICUAE-INTA, Argentina (Approved Protocol n° 54/2016). Groups of 5 mice were intradermically (i.d.) inoculated in the back, at days 0 and 15, with 15 μg of pCIgD and 75 μg of pDNA3.1-mCD40L (pCD40L) formulated without and with Montanide 101 adjuvant in a final volume of 0.2 mL (distributed in two sites) (The dose of plasmid injected was previously selected based on dose-response curve). The same amount of pCIneo was inoculated as negative control group. Serum samples were obtained at days 14 and 30.
Virus
BoHV-1 strain LA (Los Angeles) was propagated in Madin Darby bovine kidney (MDBK) cells, in E-MEM medium supplemented with 10% fetal bovine serum (Gibco). The virus was added at a multiplicity of infection (moi) of 0.1, and it was allowed to adsorb for 45 min at 37°C before the addition of more culture medium. The supernatant was collected, and cell debris was removed by centrifugation at 23,699 g for 1 h at 4°C, when cytopathogenic effects were observed. Supernatants of infected cells were pelleted by ultracentrifugation at 120,000 g for 1 h at 4°C. The virus suspension was stored at −70°C. Inactivated virus (30 min at 11 cm from two General Electric G875 ultraviolet bulbs) was used for enzyme-linked immunosorbent assay (ELISA) and in vitro cell stimulation.
Assembly of gD expression vector and production of recombinant gD
The sequence coding for full-length gD was amplified by PCR from BoHV-1 DNA, using the following forward and reverse primers: 5′-AAGAATTCATGTTGCCTACACCCGCGCCGCGGGT-3′ and 5′-AAGAATTCTCAGGCGTCGGGGGCCGCGGGCGTA-3′ that contain EcoRI restriction sites and amplify a 1005 bp fragment (nt 138 to 1143) of the gD gene (emb AJ004801 BHV1CGEN). The ATG start codon for expression in bacteria is provided by the pRSETB vector before a 6 His-codon stretch, while the reverse primer includes the gene stop codon. The 1005 bp fragment was inserted into plasmid pRSET-B (Invitrogen) to produce pRSETgD. The correct orientation and the integrity of the gD sequence were confirmed by nucleotide sequencing and restriction analysis (data not shown). E. coli BL21 cells were used to express the recombinant plasmid by standard methods. Expression was induced with 1 mM IPTG (Sigma) for 2 h when the absorbance was 0.3 at 600 nm. Proteins were extracted with 6 M guanidine HCl and 0.5 M NaCl (denaturing buffer), and gD-his protein was purified by Ni-nitrilotriacetic acid (Ni-NTA) Agarose columns (Invitrogen).
ELISA for detection of anti-BoHV-1 antibodies
Immulon 1 plates were coated with 50 μL of inactivated BoHV-1 (iBoHV-1) (2 × 107 TCID50/mL) in 0.1 M carbonate-bicarbonate buffer, pH 9.6. The plates were incubated overnight (ON) at 4°C. They were blocked, and serial dilutions (1:10) of mice sera were diluted in PBS/0.05% Tween 20 (PBST) containing 1% ovalbumin (PBST-OVA) and dispensed in 50 μL/well aliquots. After incubation, plates were washed with PBST, and antimouse IgG peroxidase conjugate (KPL) was added for 1 h at 37°C. Finally, ortho-phenylene-diamine (1,2-benzenediamine) dihydrochloride (OPD; SIGMA) and H2O2 were added, and absorbance was measured at 492 nm in a MR 5000 microplate reader (Labsystems, MN). The cutoff was established as the mean A492 of the negative sera +2 standard deviations. Titers were expressed as the log10 of the reciprocal of the highest serum dilution giving an A492 higher than the cutoff.
IgG1, IgG2a, IgG2b, and IgG3 isotyping ELISA
Greiner microtiter plates were coated with purified recombinant gD (1,4 μg/well) to evaluate the IgG1, IgG2a, IgG2b, and IgG3 subtypes in vaccinated mice. Plates were blocked with PBST containing 0.5% gelatine (PBST-G), and then serial dilutions (1:10) of mice sera were prepared in PBST-G and added to plates in 50 μL/well aliquots, for 2 h at room temperature. PBST was added to wash the plates. Biotinylated goat antimouse IgG1, IgG2a, IgG2b, and IgG3 (Caltag Laboratories, San Francisco, CA) were dispensed and after 60 min incubation, plates were washed, and antimouse IgG peroxidase conjugate (KPL) was added and incubated for 1 h at room temperature. The reaction was visualized as described in section “ELISA for detection of anti-BoHV-1 antibodies.”
Percentage of BoHV-1 neutralization
When seroneutralizing antibodies were measured in sera of immunized mice at 30 dpv using 100 DICT50 infective tissue culture doses, all values were negative. So, it was decided to measure the percentage of neutralized virus incubated 1/8 serum dilution at 25 μL of volume with different fivefold dilutions of infective BoHV-1 (1000 to 1 TCID50 per 25 μL) in quadruplicates. Then, the virus–serum mixture, previously incubated ON at 37°C under 5% CO2, was seeded on MDBK cell monolayers. After 1 h at 37°C, cells were washed with PBS. Finally, cells were incubated with fresh medium supplemented with 2% FCS at 37°C under 5% CO2. After 72 h, cytopathic effects were observed. The remnant infective virus was determined by a TCID50 assay. The calculated viral titer was expressed as the percentage of BoHV-1 that was still infective after serum incubation, compared with the titer of virus suspensions incubated without serum (a 25 μL volume of medium was incubated with the different dilutions of infective BoHV-1).
Cytokines detection
Spleens were extracted from mice, and splenocytes were disaggregated by gentle homogenization in RPMI 1640 medium/10% FCS (Gibco)/1% ampicillin-streptomycin, 15 days after the booster (30 dpv). In 96-well flat-bottomed plates, pool of splenocytes from each group were plated (1 × 106 cells per well) in 100 μL complete medium [RPMI 1640/1% ampicillin-streptomycin/1 mM sodium pyruvate (Gibco)/10% FCS/5 mM HEPES (Gibco)/2 mM L-glutamine (SIGMA)/50 mM 2-mercaptoethanol/10 mM MEM Nonessential Amino Acids Solution (Gibco)]. In triplicate 100 μL of inactivated BoHV-1 was added to the wells in complete medium or medium alone, and incubated for 48 h at 37°C. The supernatant was collected, and interferon gamma (IFNγ), interleukin (IL)-4, IL-6, IL-17A, and IL-10 were measured using BD CBA Kits (Mouse Th1/Th2/Th17) by flow cytometry. The results were expressed in pg/mL.
Cytotoxic T cell assay (JAM test)
Cytotoxicity assay was previously described by Matzinger et al. (50). In brief, splenocytes were isolated as described in section “Cytokines detection.” Cells were cultured with inactivated BoHV-1 or medium alone for 5 days at 37°C and 5% CO2. P815 cells infected with a moi of 5 of BoHV-1 and labeled with [methyl-3H] thymidine were used as target cells. 104 P815 cells per well were plated with dilutions of effector cells (splenocytes) and incubated in triplicate for 3 h at 37°C. The effector/target ratio used in the assay was 80:1. A semiautomatic cell harvester (Skatron) was used to harvest the cells. Radioactivity was measured by liquid scintillation counting (experimental release). Spontaneous release was determined in target cells incubated with complete medium only. Percentages of specific lysis were calculated using the formula:
% Specific lysis = 100 × [(spontaneous release – experimental release)/spontaneous release].
Immunofluorescent staining
The following labeled monoclonal antibodies were used to stain the surface markers of splenocytes: allophycocyanin anti-CD8b, fluorescein isothiocyanate antimouse CD4, and conjugated rat Ig isotype controls (Pharmingen). Splenocytes were incubated with labeled antibodies for 20 min at 4°C. Then, they were washed with PBS containing 1% FCS and fixed with 0.2% paraformaldehyde. The stained cells were evaluated by flow cytometry in a BD FacsCalibur and analyzed with CellQuest software.
In vitro CFSE proliferation assay
Carboxylfluorescein diacetate succinimidyl ester (CFSE; 5 nM) (BD Biosciences) was used to label splenocytes for 40 min at 37°C. Cells were washed four times and resuspended in RPMI complete medium. The cells were cultured with medium, iBoHV-1 or Concanavalin A for 4 days. Then cold PBS was used to wash the splenocytes. Fixation was done by resuspension in 0.2% paraformaldehyde in FACSFlow (BD, Buenos Aires, Argentina) and analyzed by cytometry.
Statistical analysis
Data between three or more groups were analyzed using one-way analysis of variance (ANOVA) and Dunnett's post-tests. Differences were considered significant at p-value <0.05.
Results
pCIgD plus 101 adjuvant enhances endocytosis effect on DCs
Immature DCs are specialized to sense the microenvironment and incorporate the antigen through phagocytosis or endocytosis. With the aim to determine if the selected adjuvant for the genic vaccine or the DNA vaccine were able to affect the antigen uptake by endocytosis in naïve DCs, these cells were cultured in vitro with pCIneo, pCIgD, pCIgD+101, 101 adjuvant alone, or medium like a basal control. Finally, cells were incubated with OVA coupled to FITC for 40 min at 37°C. Since this is a temperature-dependent phenomenon, the reaction control was DCs incubated with OVA-FITC at 4°C. Endocytosis levels were then measured by flow cytometry.
The 101 adjuvant plus pCIgD produced a significant increase of endocytosis in these cells. On the contrary, pCIgD, pCIneo, or 101 adjuvant alone did not modify endocytosis levels (Fig. 1).
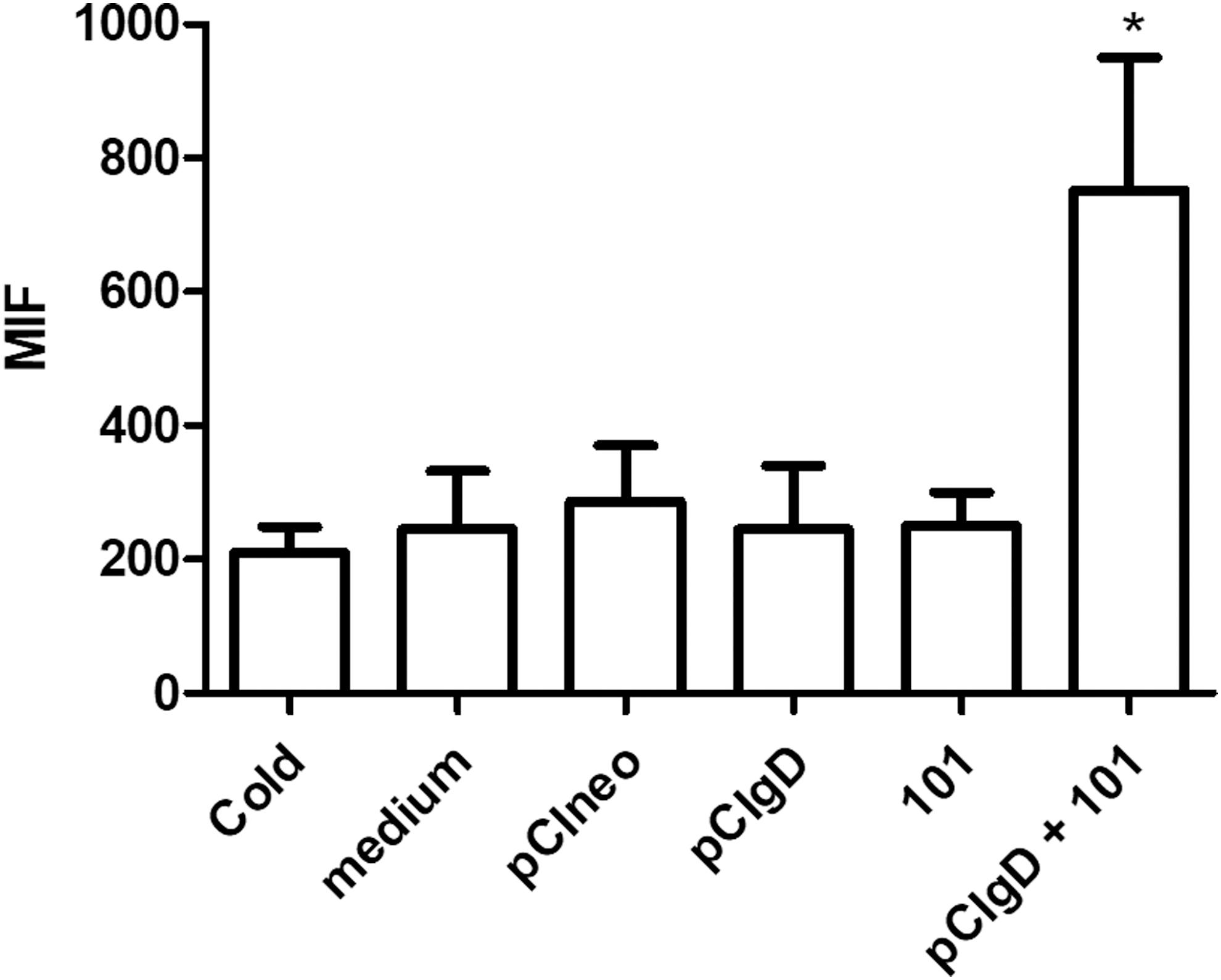
Adjuvant effect on BMDC. Endocytosis of OVA-FITC antigen by BMDC cultured in vitro with different formulations for 48 h was tested. Results are expressed as the MFI ± SEM of five independent experiments. BMDC, bone marrow-derived dendritic cells; MFI, mean fluorescence intensity; SEM, standard error of the mean. *p < 0.05.
In previous studies, cattle were immunized with pCIgD and adjuvant 101. After viral challenge, higher protection levels were observed as compared with animals immunized with pCIgD alone (61). For this reason, adjuvant 101 was selected for this study, with the addition of a plasmid encoding CD40L, to enhance the immune response against gD.
pCIgD vaccines formulated with pCD40L, 101 adjuvant, and pCD40L plus 101 elicit higher specific humoral immune responses than a pCIgD vaccine. Neutralizing capacity of the sera
Groups of mice were vaccinated with 15 μg of pCIgD (per dose) formulated with and without pCD40L, 101 adjuvant, and a combination of both, and the elicited immune responses were evaluated and compared. Analysis of sera by ELISA to BoHV-1 showed that at 14 dpv, all animals in groups immunized with pCIgD+pCD40L, pCIgD+101, and pCIgD+pCD40L+101, were able to induce specific BoHV-1 antibodies. Moreover, at 30 dpv, levels of antibodies against BoHV-1 in groups pCIgD+pCD40L, pCIgD+101, and pCIgD+pCD40L+101 were significantly higher than those in the pCIgD group (p < 0.01). None of the pCIneo-immunized control animals produced any specific immune response at any time compared with normal mouse serum (Fig. 2A). Moreover, analysis of sera by ELISA showed that specific antibodies against the gD-protein were induced at 14 dpv. Total antibody levels were significantly increased (p < 0.05) at 14 dpv in the pCIgD+101 group compared with the pCIgD group. In contrast, at 30 dpv all immunized groups showed the same high antibody titers (Fig. 2B).
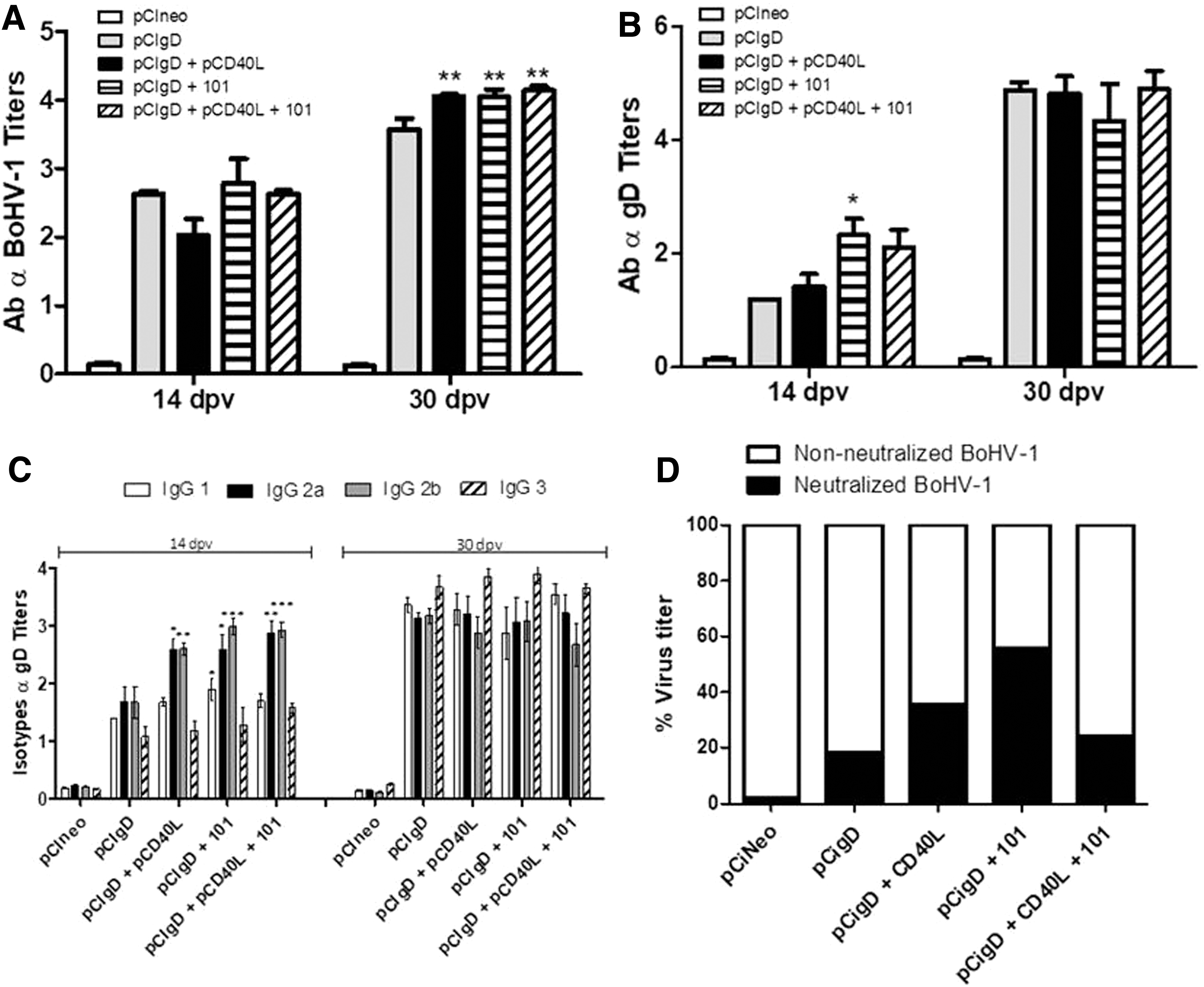
Humoral immune response elicited.
Increases in IgG isotype induced by the different vaccines were studied next. At 14 dpv, IgG2a and IgG2b were the main isotypes in the sera of mice vaccinated with pCIgD+CD40L, and were significantly increased with respect to the pCIgD group (IgG2a p < 0.05; IgG2b p < 0.01). The same was observed in the group immunized with pCIgD+101 adjuvant (vs. pCIgD: IgG2a, p < 0.05; IgG2b, p < 0.001) and with pCIgD+pCD40L+101 adjuvant (vs. pCIgD: IgG2a, p < 0.01; IgG2b, p < 0.001). Furthermore, group pCIgD+101 presented significantly higher levels of specific IgG1 (p < 0.05) than the pCIgD group. At 30 dpv, although specific gD isotype titers were high, IgG1 and IgG3 showed higher levels, and no significant differences between groups were observed (Fig. 2C).
The viral neutralizing capacity of sera from immunized animals was studied in a 1/8 dilution. At 30 dpv (Fig. 2D), the average percentages of neutralized virus were 1.8 for sera in the pCIneo group, 18.4, 35.7, 55.7, and 24.4 for the pCIgD, pCIgD+CD40L, pCIgD+101, and pCIgD+CD40L+101 group, respectively. However, when antibodies were evaluated in a neutralization test with a fixed amount of virus, no serum sample at 30 dpv was able to neutralize 100 TCID50 of BoHV-1 (virus neutralization titers <0.9, data not shown). This assay evidenced a weak capacity of the antibodies to neutralize virus in all animals. However, a differential neutralizing capacity was observed among groups, being the animals immunized with pCIgD+101 those that managed to neutralize the largest amount of viral particles in comparison to the other groups.
Cytokine production
To profile the cytokine levels induced by different vaccines at 30 dpv, production of some cytokines was evaluated by cytometric bead arrays on a pool of supernatant of splenocytes stimulated or not with inactivated BoHV-1 or concanavalin A, as a positive control, from each group of vaccinated mice.
The pCIgD vaccine group secreted lower levels of IFNγ, IL-4, IL-6, IL-17, and IL-10 cytokines than the other formulations containing pCIgD (Fig. 3A–E).
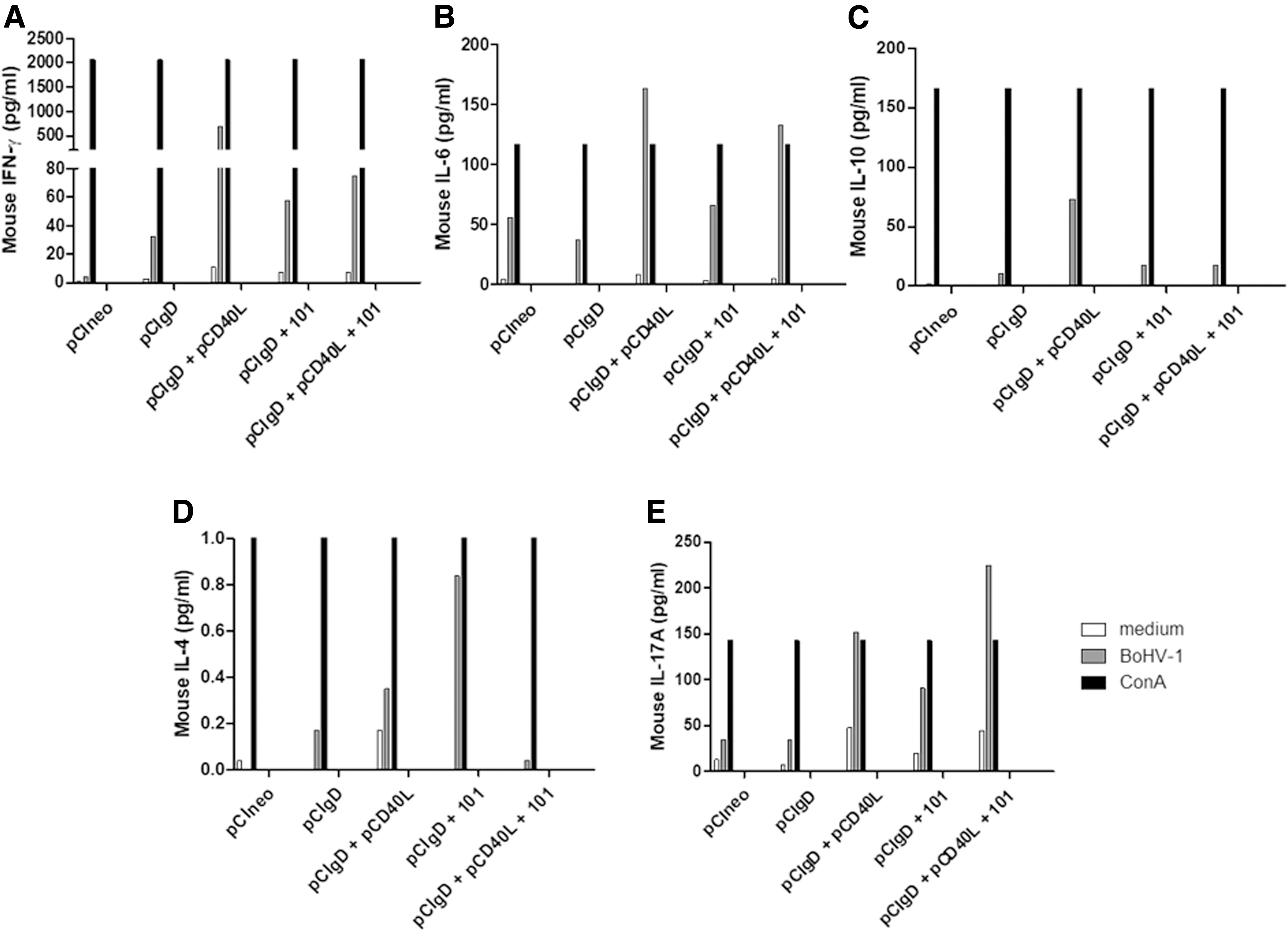
Cytokine detection with CBA. Splenocytes were stimulated or not for 48 h with inactivated BoHV-1 or concanavalin A, as a positive control, and cytokine production was detected using a Cytometric Bead Array (CBA) kit.
The amount of IFN-γ produced upon vaccination with pCIgD+pCD40L (694.64 pg/mL) was ∼20 times higher than the amount produced upon vaccination with pCIgD alone (32.49 pg/mL), and 11 times higher than those in the pCIgD+101 or pCIgD+pCD40L+101 groups (Fig. 3A). Also, in the case of IL-6 and IL-10, pCIgD+pCD40L group is the major producer of both cytokines as compared with the other groups (Fig. 3B, C). To highlight in this sense that the pCIgD+pCD40L+101 group secrete similar levels of IL-6 than the pCIgD+pCD40L group. In case of IL10, the pCIgD+pCD40L group was the only group that produces this cytokine (Fig. 3C).
Besides, low levels of IL-4 were detected among the pCIgD, pCIgD+pCD40L, and pCIgD+pCD40L+101 groups, pCIgD+101 being the major producer group of this cytokine (Fig. 3D).
Interestingly, production of IL-17A could be detected in the pCIgD+pCD40L, pCIgD+101, and pCIgD+pCD40L+101 groups but not in the pCIgD one (Fig. 3E). Animals immunized with pCIneo only secreted low levels of IL-6.
Combination of pCIgD with pCD40L and 101 adjuvant increase splenic CD4+ T lymphocytes cell population
Spleen cells obtained from vaccinated mice were used to determine the effect of these vaccines on T lymphocyte populations.
A significant (p < 0.05) expansion of CD4+ T cells population was observed at 30 dpv in the pCIgD+101+pCD40L group as compared with pCIneo and pCIgD (Fig. 4A). No significant differences, on the contrary, were observed in the CD8+ lymphocyte populations measured (Fig. 4B). In the other groups, no changes were observed in these two populations.
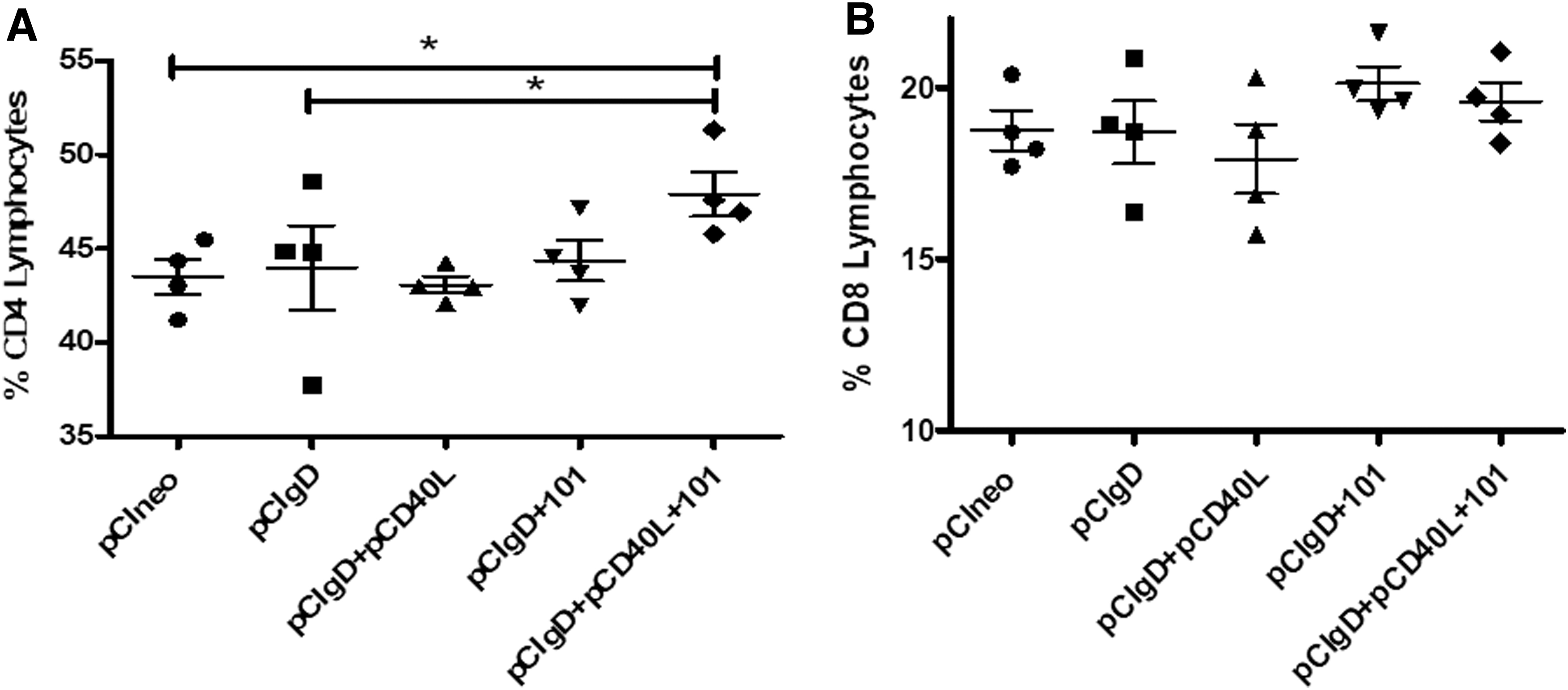
Cellular spleen subpopulations from vaccinated mice at 30 dpv. Each point corresponds to one animal of the group. Results are expressed as population percentages (means ± SEM)
Vaccination with pCD40L+101 enhances the specific cytotoxic T cell in vivo response against BoHV-1
Using a specific viral cytotoxicity test (JAM test), splenocytes of pCIgD mice with or without pCD40L and/or 101 adjuvant and pCIneo groups were restimulated in vitro with UV-inactivated BoHV-1 and then incubated with radioactively labeled BoHV-1-infected P815 cells.
CTL activities in the three groups that received pCIgD plus pCD40L and/or 101 adjuvant, which were restimulated in vitro with iBoHV-1, were significantly higher (pCIgD+pCD40L, 21.12% p < 0.01; pCIgD+101, 18.04% p < 0.05; pCIgD+pCD40L+101, 26.22% p < 0.001) than those in the pCIgD group (6,95%) (Fig. 5). However, when a cutoff was selected (pCIneo mean ± 2DS = 20% specific cytotoxicity) only pCIgD+pCD40L+101 had all animals over the cutoff with specific positive cytotoxicity activity. In contrast, only one and two animals were positive in the pCIgD+pCD40L and the pCIgD+101 groups, respectively. These results indicate that the gD-encoding plasmid plus pCD40L, 101 adjuvant, or both induce specific CTL that recognizes BoHV-1-infected cells. Yet, only combined with 101 and pCD40L, pCIgD has the capability to achieve consistently high specific cytotoxicity toward BoHV-1 in all animals of the group.
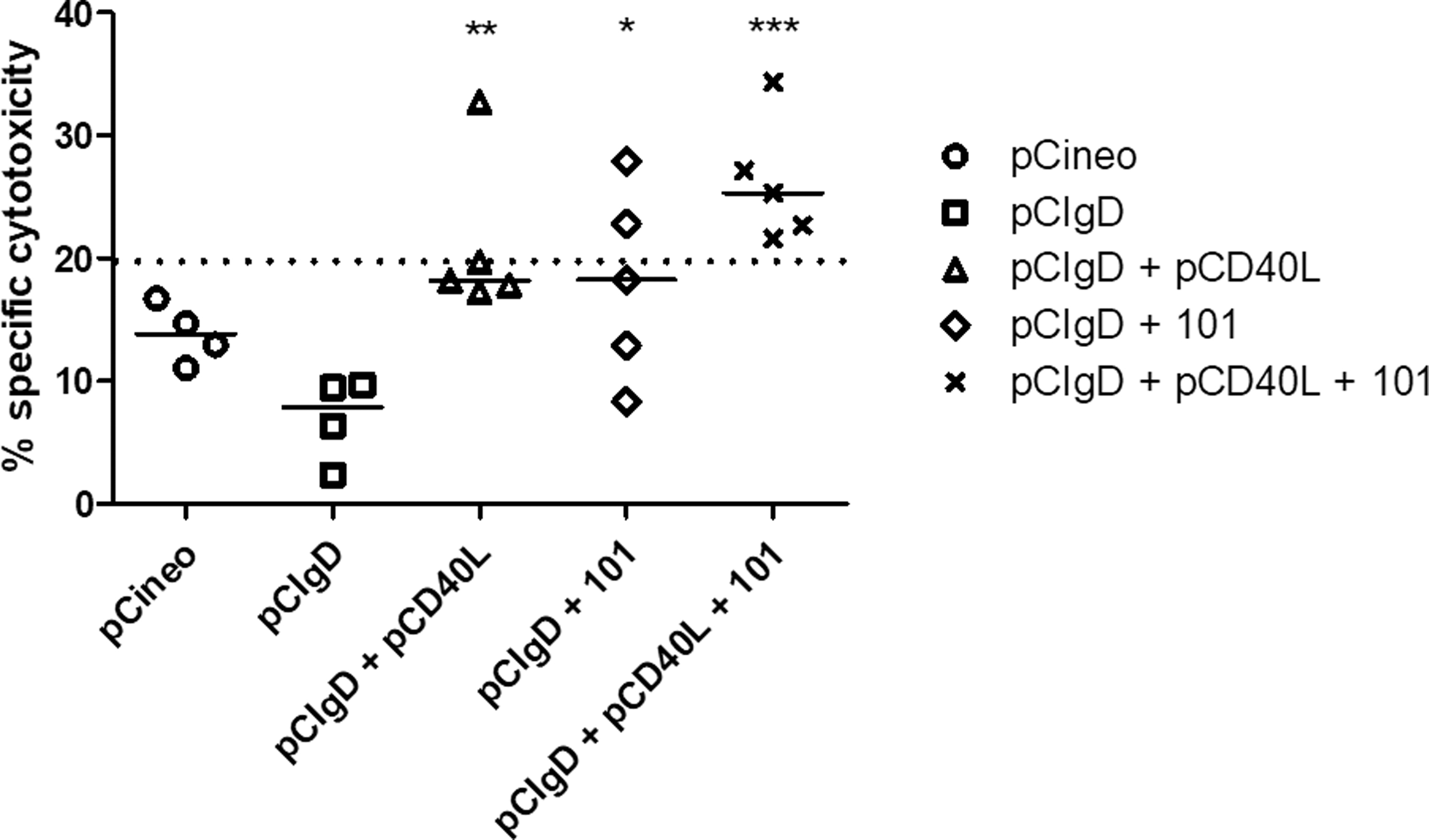
Immunization with pCIgD and adjuvants induces an enhanced CTL response as compared with pCIgD alone. Splenocytes collected at 30 dpv and restimulated in vitro with BoHV-1, and were tested for CTL response by a cytotoxicity assay against the virus. Representative results of percentage specific lysis are shown at an effector/target ratio of 80:1. *p < 0.05, **p < 0.01, and *** p < 0.001 (statistical significance of differences with respect to the pCIgD group). A cutoff of 20 was calculated as the mean of the pCIneo group +2 SD. CTL, cytotoxic T lymphocytes.
Vaccination with pCIgD+pCD40L or 101 adjuvant enhances BoHV-1 specific T cell proliferation
Since the cellular immune response is essential to control a BoHV-1 infection, specific T cell proliferative responses were measured. As shown in Table 1, there are more animals with a viral-specific cell proliferative response considered as positive (above the cutoff = 3.64) in the pCIgD+101 and pCIgD+pCD40L groups (60% positive proliferation in both groups against 25% in the pCIgD group). Surprisingly, under these experimental conditions, no specific proliferation was detected in the pCIgD+pCD40L+101 group. All splenocytes stimulated with concanavalin A had a proliferative response.
T Cell Proliferation in Response to Vaccination with pClgD Alone and Formulated with 101 and/or CD40L
Spleen cells were isolated from animals inoculated with pCIgD with adjuvants and in vitro stimulated with iBoHV-1 for 4 days, followed by measurement of T cell proliferation by lost of CFSE.
% Proliferation = % proliferation stimulated with BoHV-1 − % proliferation stimulated with medium.
Results in bold and underline are over the cutoff. The cutoff is calculated as pClneo mean + 2 SD (cutoff = 3.64).
BoHV, bovine herpesvirus-1; SD, standard deviation.
Discussion
The major problem to control BoHV-1 resides in the capability of the virus to produce latency and immune evasion in infected animals. During infection, the virus propagates intracellularly followed by an extracellular dispersion, and persists even in the presence of neutralizing antibodies. Thus, BoHV-1 vaccines should induce humoral and cellular immune responses with induction of CTL (44,46).
Huang et al. (32) used a plasmid coding BoHV-1 glycoprotein B (gB) to induce cellular and humoral immune responses in mice, and showed production of specific antibodies and CTL responses. The same vaccine induced humoral responses and elicited IFNγ secreting CTL in calves (32). This was the first demonstration that the plasmid induced CTL not only in mice but also in calves. The most important glycoprotein in BoHV-1 is glycoprotein D (gD), which is involved in the protective immune response (3,33), with major targeting to CD8+ CTL (16,17,40), and with the production of neutralizing antibodies (3,17,33,58,76,77,82). In this study, a secreted form of BoHV-1 gD was chosen because previous studies of others performed in mice and cattle showed that this truncated version of gD induced higher antibody titers than the membrane-anchored form (41,42,75).
Given the importance of adjuvants to increase vaccine efficiency, the immunomodulatory effects on a DNA vaccine of Montanide 1113101PR (101) adjuvant (SEPPIC) were previously analyzed in our laboratory. We demonstrated that a DNA vaccine based on BoHV-1 gD formulated with 101 adjuvant induced not only specific antibodies but also cellular immunity against BoHV-1 with viral-specific T cell proliferation and the high levels of IFNγ secretion in cattle (33).
This study examined the benefits of the CD40 ligand, CD40L. This molecule belongs to the tumor necrosis factor superfamily, hasa transient expression in activated T lymphocytes, and can stimulate DCs and B cells (27). This stimulation activates DCs and primes the CD8+ T cell response (5,64,68). Also, CD40L increases the survival and differentiation of activated B cells (20). These qualities make CD40L an attractive vaccine adjuvant (8,30,34,51,69,81).
Incorporation of any of these adjuvants (101, pCD40L, or both) to gD vaccine formulations yielded an increased humoral response toward BoHV-1 and gD as well. Similar results for 101 and CD40L were reported by Quattrocchi et al. (61) and Manoj et al. (48), respectively. Vaccines formulated with these adjuvants yielded high levels of antibodies in cattle, but insufficient protection upon BoHV-1 challenge.
Analysis of isotypes in serum showed that IgG1, IgG2a, IgG2b, and IgG3 levels were similar in all animals, but the major variation was observed in IgG2a and IgG2b at 14 dpv. An increase in IgG2a has been described for DNA vaccines (63) and suggests a T-helper 1 response. Predominance of IgG2a has been described for humoral protective immune responses to virus in mice. In these animals, this isotype is clearly more efficient at complement fixation (11,12,37,56) and as opsonizing agent than IgG1 (1). Detection of lymphocyte proliferation after inactivated BoHV-1 stimulation and isotype switch evidenced by IgG2a secretion indicates a balance to a Th1/Th2 profile of CD4+ T cells (11,12,53,72).
One feature of adjuvants is their inflammatory effect that attracts inflammatory cells, including macrophages and DCs (14,78). This characteristic plays an important role in the immune response. On the contrary, the formulation of pCIgD with 101 adjuvant can be efficiently endocytozed by APCs, generating an increase in the proliferative response of other cells (29). It can be hypothesized that in the present experiments the 101 adjuvant recruited APCs at the site of inoculation, facilitating DNA internalization, as observed by Dupuis and collaborators (19).
Since cellular immune response is important to deal with BoHV-1 infection, the specific T cell proliferative response was measured. Our results showed that this response was incremented in the pCIgD+101 and pCIgD+pCD40L groups (60% positive proliferation in both groups against 25% in the pCIgD group). Surprisingly, under these experimental conditions no specific proliferation was detected in pCIgD+pCD40L+101 group. This could be due to expensive cell death after 4 days of proliferation, when cells were harvested.
Genetic immunizations were used in several animal models and hosts, such as mice and cattle (9,13,61,75). It has been demonstrated by Deshpande et al. (17) that induction of specific CTL against BoHV-1 was elicited by membrane-anchored gD. The same phenomenon was demonstrated by our group inoculating a DNA vaccine encoding a secreted version of gD (40). The pCIgD vaccine was formulated with 101 and pCD40L with the expectation of inducing CTL against the virus, and a specific cytotoxicity assay was set up with BoHV-1-infected P815 cells. This experiment demonstrated that the mice vaccinated with different formulations had the capacity to elicit CTL production. We observed the occurrence of memory CD8+ cells that recognized and lysed P815-infected cells. The specific cytotoxicity was consistent with the increases in IL-17A detected in pCIgD+101, pCIgD+pCD40L, and pCIgD+101+pCD40L. IL-17A is required for optimal specific CD8+ T cells activity. In addition to Th17 cells, there are other sources of IL-17, such as Tγδ, NKT, and CD8+ T cells (45,83). In DCs, IL-17A upregulates surface MHC class I molecules and enhances production of IL-6. This facilitates crosspriming and proliferation of CTL. In the same sense, secretion of IFNγ indicates the activation of a Th1-type response.
Taking together, our results indicate that formulation of a pCIgD DNA vaccine with adjuvants 101 and/or pCD40L elicits a higher humoral and cellular immune response against this virus than the plasmid alone in mice. They also show that only when both adjuvants are included, CTL is increased in all immunized animals. Because of this, we suggest that a pCIgD vaccine formulated with 101 and pCD40L should be tested in bovines since this formulation likely induces protection against BoHV-1 in these animals.
Footnotes
Acknowledgments
The authors thank Dr. Osvaldo Zabal for the valuable assistance with cell cultures, Seppic for providing the Montanide ESSAI 1113101PR adjuvant, Dr. Kipps for providing the pDNA3.1-mCD40L in this study, and Ms. Pamela Angeletti and Fátima Torales for their technical assistance.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by INTA Project PNBIO 1131032 and by a grant from Agencia Nacional de Promoción Científica y Tecnológica PICT2014-0891.