
Abstract
DNA-dependent activator of interferon regulatory factors (DAIs), interferon gamma inducible protein 16 (IFI16), DEAD-box polypeptide 41 (DDX41), DNA-dependent protein kinase (DNA-PK), meiotic recombination 11 homolog A (MRE11), and cyclic GMP-AMP synthase (cGAS) have been identified as intracellular STING-dependent DNA sensors in recent years. Studies have shown that the DNA sensor-STING-interferon (IFN)-β pathway plays an important role in the defense against intracellular invasion of many DNA viruses. However, the intracellular recognition of hepatitis B virus (HBV) DNA by DNA sensors is still largely unclear. In this study, we aimed to determine whether the DNA sensor-STING pathway in peripheral blood mononuclear cells (PBMCs) can be activated by acute and chronic HBV infections in humans. We first evaluated the expression of these DNA sensors in PBMCs of acute and chronic HBV-infected patients by quantitative real-time polymerase chain reaction. We next compared the expression of the upregulated DNA sensor between monocytes and nonmonocytes to find its cellular source. Finally, by in vitro stimulation, we analyzed the IFN-β response of the DNA sensor-STING pathway in PBMCs and monocytes from chronic HBV-infected patients. The results showed that IFI16, DDX41, MRE11, and the adaptor STING were upregulated in chronic HBV-infected patients, whereas only IFI16 was upregulated in acute HBV-infected patients. However, IFN-β expression was not changed in PBMCs from acute and chronic HBV-infected patients. We next found IFI16 was mainly expressed in monocytes of acute and chronic hepatitis B patients. Finally, by stimulation of monocytes with VACV ds 70mer, a ligand for IFI16, we confirmed the attenuated response of the IFI16-STING pathway. Taken together, our results suggest that HBV might be sensed by DNA sensors in PBMCs of acute and chronic HBV-infected patients, and meanwhile HBV infection attenuates the response of the DNA sensor-STING pathway in PBMCs and monocytes, which may facilitate the persistence of HBV infection.
Background
Hepatitis B virus (HBV) is a noncytopathic hepatotropic DNA virus in humans. Infection with HBV results in acute or chronic hepatitis B (CHB). About 248 million people globally are chronically infected with HBV and at high risk for liver cirrhosis and hepatocellular carcinoma (1). The mechanism of development of chronicity of HBV infection remains to be determined. The host's age and immune status significantly influence the outcome of acute infection, as the rate of chronicity after acute infection is >90% among children <1 year and is <5% among infected adults (21). Thus, the host immune status appears to be a major determinant of the clinical outcome of HBV infection. Besides, HBV is also referred to as a “stealth” virus that elicits negligible innate immune responses in the early infection phase to facilitate the establishment of chronic infection (42). It is well established that a robust and HBV-specific CD8+ T cell response is necessary to eliminate HBV infection. In contrast, the contribution of the innate immune mechanisms is not well understood and whether HBV can be sensed by the innate immune system remains controversial (13,23,35).
The invasion of mammalian cells by pathogens represents a danger signal for the innate immune system, which takes use of pattern-recognition receptors (PRRs) to recognize conserved motifs from pathogens. The intracellular DNA sensors are responsible for the recognition of cytoplasmic or nuclear pathogen-derived DNA. Recently, DNA-dependent activator of interferon regulatory factors (DAIs), interferon gamma inducible protein 16 (IFI16), DEAD-box polypeptide 41 (DDX41), DNA-dependent protein kinase (DNA-PK), meiotic recombination 11 homolog A (MRE11), and cyclic GMP-AMP synthase (cGAS) were identified as intracellular DNA sensors (10). On binding to foreign DNA, these DNA sensors activate stimulator of interferon genes (STING) (10,27), which then activates TANK-binding kinase 1 (TBK1). TBK1 subsequently phosphorylates STING and the downstream transcription factor interferon regulatory factor 3 (IRF3) to promote the expression of type I interferons (IFN-Is) and other cytokines (3,25,44). Thus, the DNA sensor-STING pathway plays a significant role in host defense against invading DNA viruses.
Previous studies have suggested that many viral infectious diseases can induce the activation of STING-dependent DNA-sensing pathways (22), and meanwhile, some viruses utilize a plethora of mechanisms to antagonize the DNA-sensing pathways (30,33,37). However, little is known about the roles of DNA sensors in HBV infection. Our previous study showed IFI16 as a component of inflammasome complexes is upregulated in both acute and chronic HBV infections in humans (4). In this study, we scrutinized all the known STING-dependent DNA sensors in peripheral blood mononuclear cells (PBMCs) of acute and chronic HBV-infected patients and found again that only IFI16, expressed most frequently in monocytes, is involved in acute and CHB in humans. Furthermore, the IFI16-STING pathway is suppressed in CHB, which may contribute to HBV-induced immunotolerance.
Materials and Methods
Subjects
From May 2015 to November 2018, 66 treatment-naive patients with chronic HBV infection were recruited at Shenzhen People's Hospital (Shenzhen, China). The patients were classified into three groups according to the guidelines for the treatment of CHB (36): immune-tolerant (IT) carriers (n = 15), hepatitis B e antigen (HBeAg)-positive hepatitis patients (CHB, n = 31), and inactive (IA) carriers (n = 20). Twenty-eight healthy individuals were enrolled as healthy controls (HCs). Patients with acute hepatitis B (AHB, n = 8) and varicella zoster virus (VZV, n = 8) were also included in this study. Patients with any other disease were excluded. The clinical characteristics of HCs and patients are shown in Tables 1 and 2. This study was approved by the Ethical Committee of Shenzhen People's Hospital. All participants signed written informed consent before sample collection.
Clinical Characteristics of Healthy Controls and Treatment-Naive Patients with Chronic Hepatitis B Virus Infection
Values are n or the median with range.
Chi-squared test.
Kruskal–Wallis H test.
ALT, alanine aminotransferase; CHB, chronic hepatitis B; HBeAg, hepatitis B e antigen; HBV, hepatitis B virus; HC, healthy control; IA, inactive; IT, immune-tolerant; ND, not determined; NS, not significant.
Clinical Characteristics of Healthy Controls and Patients with Acute Hepatitis B Virus Infection and Varicella
Values are n or the median with range.
Chi-squared test.
Kruskal–Wallis H test.
AHB, acute hepatitis B; ALT, alanine aminotransferase; ND, not determined; TBil, total bilirubin; VZV, varicella zoster virus.
Isolation of PBMCs and monocytes
Blood samples were obtained from each subject. PBMCs were isolated using Ficoll-Paque (GE Healthcare) density gradient centrifugation. CD14+ monocytes and nonmonocytes were isolated from fresh PBMCs using Dynabeads untouched human monocytes separation kits (Invitrogen, Shanghai, China). FACS analysis showed that the purity of the obtained CD14+ monocytes was >95%.
RNA extraction, reverse transcription, and quantitative real-time polymerase chain reaction
Total RNA was extracted from cells with TRIzol (Invitrogen). Target mRNA was reverse transcribed using the PrimeScript RT reagent Kit with gDNA Eraser (TaKaRa). The primer sets used in this study are shown in Table 3. The quantitative real-time polymerase chain reaction (qPCR) was conducted with the SYBR Premix ExTaq kit (TaKaRa) on the Applied Biosystems ViiA 7Dx Real-Time PCR system (Applied Biosystems, Shanghai, China). The standard curve for each target gene was established using gradient dilutions of the corresponding DNA fragments amplified by RT-PCR. Based on the standard curve, the mRNA copies of each target gene were determined. The qPCR was performed in duplicate for each sample. The expression levels of each target gene were shown as values normalized against human 103 copies of GAPDH transcripts.
Primer Sequences Used for Quantitative Real-Time Polymerase Chain Reaction
Cell culture and stimulation
PBMCs, monocytes, and nonmonocytes were cultured at 6 × 105 cells/mL in RPMI 1640 containing 10% fetal bovine serum, 2 mmol/L L-glutamine, 100 μg/mL streptomycin, and 100 U/mL penicillin at 37°C in 5% CO2 incubator. The fresh cells were cultured for 2 h and then stimulated with VACA ds 70 mer (InvivoGen) at a concentration of 2 μg/mL for 24 h according to the manufacturer's instructions. The mRNA expression of IFI16, STING, and IFN-β was detected by qPCR.
HBV DNA assays and serological assays
Serum HBV DNA loads were quantified on the ABI 7500 Real-time PCR System (Applied Biosystems). The serum levels of hepatitis B surface antigen (HBsAg), HBeAg, and anti-HBe were measured with electrochemiluminescence immunoassay on the Roche Diagnostics Cobas e601 analyzer (Roche Diagnostics, Germany). The normal range of serum ALT was 0–40 U/L.
Statistical analysis
Data were expressed as mean ± standard deviation. GraphPad Prism 8 software was used for statistical analysis. The Student's t-test was used for comparing the means of two groups. When comparing more than two groups, analyses were conducted using one-way analysis of variance, followed by the Bonferroni post hoc test. p < 0.05 was considered significant.
Results
Upregulated mRNA expression of IFI16, DDX41, and MRE11 in PBMCs of chronic HBV-infected patients
We first assayed STING-dependent DNA sensors in PBMCs from chronic HBV-infected patients. Chronic HBV-infected patients were classified into three groups on the basis of their natural histories: IT carriers, characterized by HBeAg-positive with normal ALT levels and high HBV DNA loads (>20,000 IU/mL); immune-active patients, also known as “chronic hepatitis B” (CHB) patients, characterized by elevated ALT levels and viral loads (>2,000 IU/mL) with HBeAg or HBeAb positive; the inactive carriers, characterized by HBeAg-negative with normal ALT levels and low viral loads (<2,000 IU/mL). As shown in Figure 1A, IFI16 mRNA expression was upregulated in CHB patients. DDX41 and MRE11 mRNAs also increased in IT and IA patients as compared with those in HCs and CHB patients (Fig. 1B, E). Despite the anti-HBV effect of cGAS and DAI in cell culture and animal models (5,7,15), the cGAS and DAI expression was not enhanced in the chronic HBV-infected groups (Fig. 1D, F), and DNA-PK expression did not change as well (Fig. 1C). In addition, as shown in Supplementary Figure S1, the expression levels of IFI16 and DDX41 mRNA were significantly positively correlated with the CHB patients' HBV DNA loads (r = 0.6578, p < 0.0001; r = 0.5889, p = 0.0016) (Supplementary Fig. S1A, B). Enhanced serum IFI16 protein levels were also observed in CHB patients (Supplementary Fig. S2). With enhanced expression of IFI16, DDX41, and MRE11 in CHB, IT, and IA patients, the adaptor STING expression was also upregulated in all chronic HBV-infected groups (Fig. 1G). Interestingly, we did not observe the induction of the downstream effector cytokines IFN-β (Fig. 1H) and TNF-α (data not shown), in keeping with other evidence showing that HBV polymerase could interfere with STING ubiquitination modification to suppress IRF3 activation and IFN-β expression (26). Taken together, these data suggest that HBV might be sensed by intracellular DNA sensors in PBMCs during chronic HBV infection but may exert opposite effects on activation of the DNA sensor-STING pathway.

Expression of DNA sensors in PBMCs of chronic HBV-infected patients. The amounts of IFI16, DDX41, DNA-PK, cGAS, MRE11, DAI, STING, and IFN-β mRNA were quantified using qPCR
IFI16 expression is upregulated in PBMCs of AHB patients
The incidence of AHB is extremely low (<4.4 per 100,000 population) (11,17,29,34), making it difficult to conduct clinical studies. Therefore, AHB is always less understood as compared with chronic HBV infection and the contribution of innate immunity to the control of AHB is even less reported. Varicella is an acute and self-limited infectious disease caused by VZV. VZV and HBV are both DNA viruses. However, in contrast to AHB, VZV infection of healthy people at any age is almost always self-limited. Therefore, we used patients with varicella as the control cases of AHB to compare differences in the expression of DNA sensors in PBMCs.
As shown in Figure 2A, only IFI16 expression was upregulated in AHB patients as compared with HCs, and other DNA sensors in AHB patients were strikingly downregulated (Fig. 2B–F), analogous to the reduced innate immune responses in AHB patients in a previous study (12). More interesting is, except IFI16, all the other DNA sensors in varicella were also significantly downregulated (Fig. 2B–F). Despite the elevated expression of IFI16, IFN-β expression was not altered in AHB (Fig. 2H), supporting the idea of a lack of induction of IFN-Is in AHB (12). These results suggest that HBV might be sensed by IFI16 in acute HBV infection and can counteract the induction of IFN-β in PBMCs.
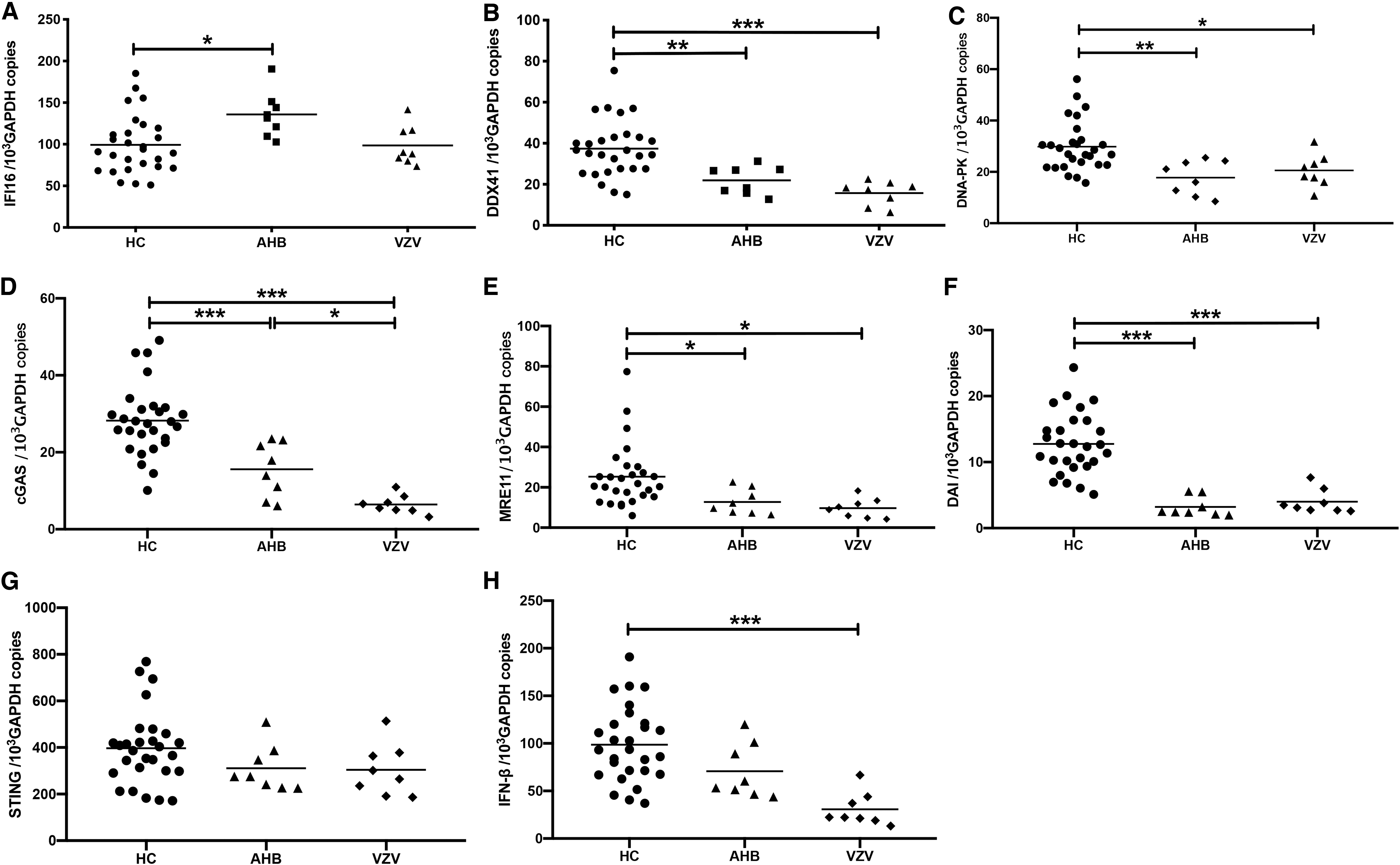
Expression of DNA sensors in PBMCs of acute HBV-infected patients. The amounts of IFI16, DDX41, DNA-PK, cGAS, MRE11, DAI, STING, and IFN-β mRNA were quantified using qPCR
IFI16 is mainly expressed in monocytes of AHB and CHB patients
As enhanced IFI16 expression was involved in both CHB and AHB, we next investigated the cellular source of IFI16. Previous studies suggested that IFI16 is mainly expressed in the CD14+ monocyte lineage (2,8). Besides the monocyte lineage, lymphoid cell lines also constitutively express IFI16, but the expression is not affected by IFNs (8,9). Therefore, to compare IFI16 expression in immune cells, we divided PBMCs into monocytes and nonmonocytes. As shown in Figure 3, IFI16 was mainly expressed in monocytes in HCs (p < 0.001) and its expression level was significantly upregulated in monocytes from AHB and CHB patients, suggesting that HBV might be sensed by monocytes, which is in keeping with previous studies (6). In addition, IFI16 expression levels in PBMCs were significantly positively correlated with serum HBV DNA loads in CHB patients (Supplementary Fig. S1A). Taken together, these findings imply that HBV might be sensed by monocytes and its infection induces enhanced expression of IFI16 in monocytes of AHB and CHB patients.
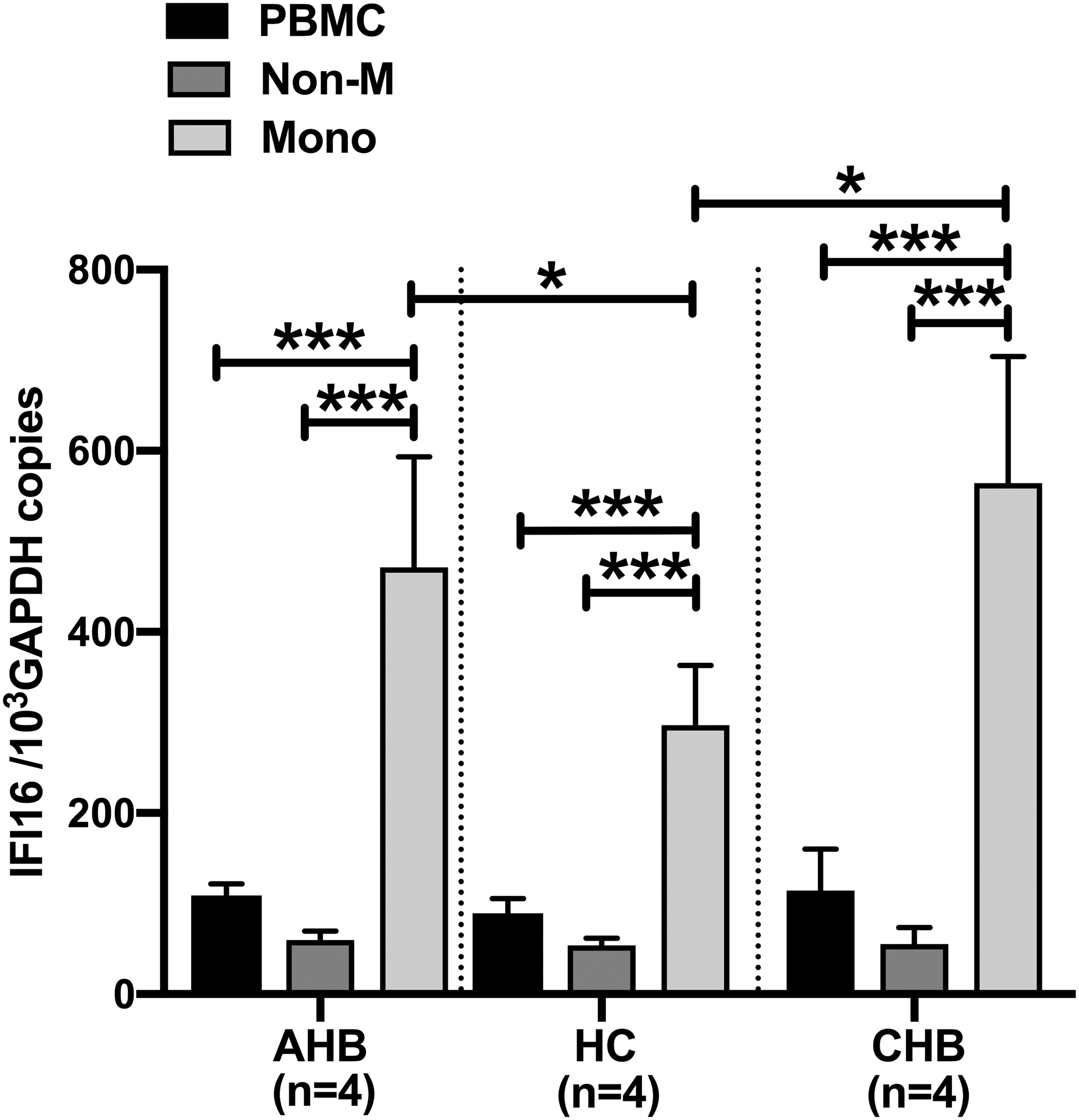
IFI16 expression in PBMCs, monocytes, and nonmonocytes. IFI16 mRNA expression was determined by qPCR. Statistical analyses were performed using a Student's t-test. *p < 0.05, ***p < 0.001.
Stimulation of IFI16 expression by its ligand VACV ds 70mer is attenuated in monocytes from CHB patients
Although HBV infection resulted in enhanced expression of IFI16 and STING in CHB patients, we did not observe increased expression of IFN-β in CHB patients accordingly. We speculate that the IFI16-STING-IFN-β pathway may be suppressed by HBV in monocytes of CHB patients. Previous studies showed that as a ligand for IFI16, the VACV double-stranded DNA 70-base pairs motif (VACV ds 70mer) can potently induce IFN-β in PBMCs and monocytes (38). To confirm our speculation, PBMCs and monocytes were isolated from HCs and CHB patients and stimulated in vitro by VACV ds 70mer to assay the response of the IFI16-STING-IFN-β pathway. As shown in Figure 4A–D, the expression of IFI16 and STING in PBMCs and monocytes of CHB patients was significantly attenuated as compared with HCs after 24 h of stimulation by VACV ds 70mer. As a result, the IFN-β response to VACV ds 70mer was severely suppressed in PBMCs and monocytes of CHB patients (Fig. 4E, F). These results support our speculation that the response of the IFI16-STING-IFN-β pathway to HBV is attenuated in monocytes of CHB patients, consistent with the finding that HBV is recognized mainly by liver macrophages (Kupffer cells) but does not induce an interferon response (16).
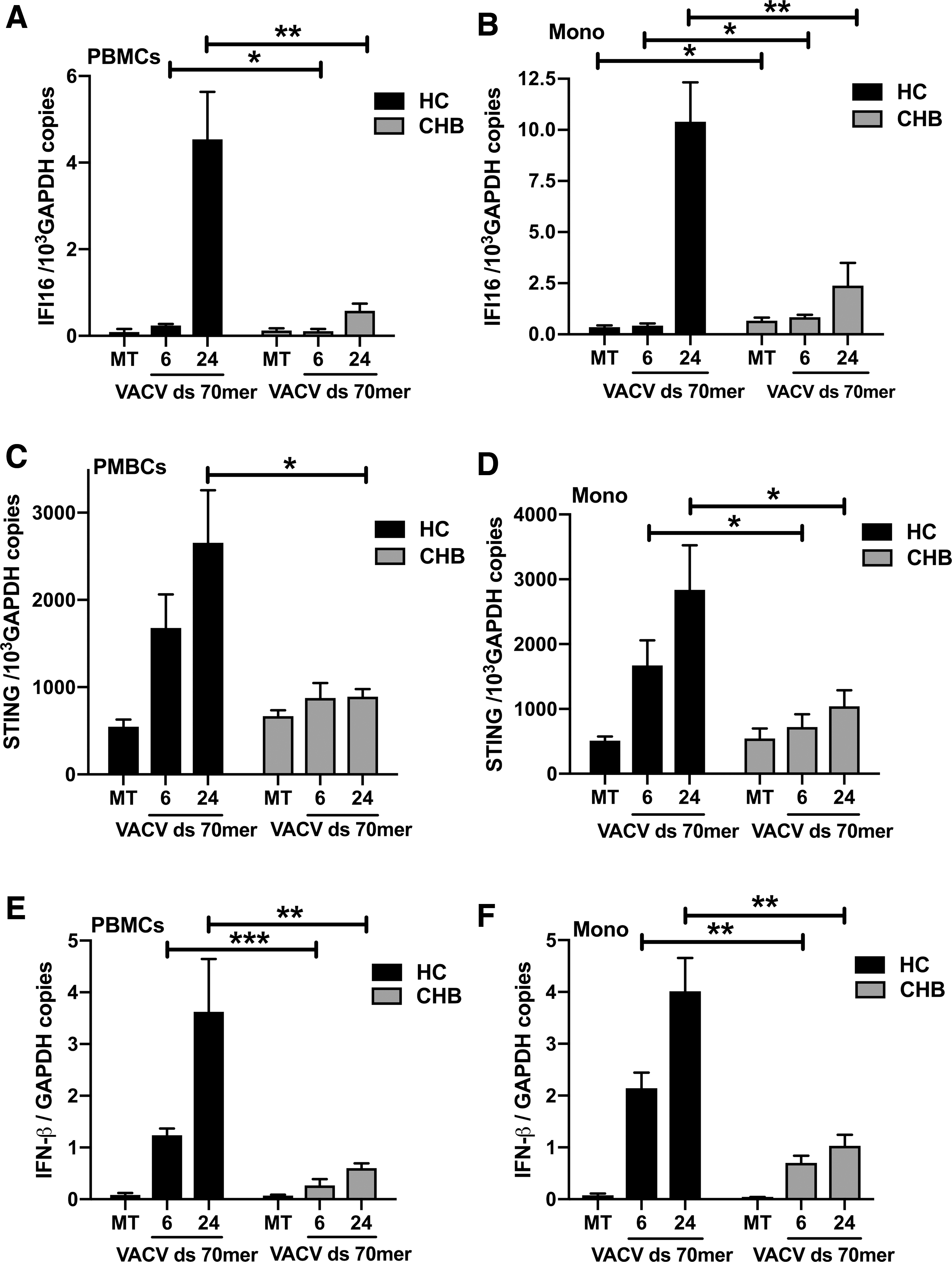
Stimulation of IFI16 expression by VACV ds 70mer is attenuated in PBMCs and monocytes from CHB patients. PBMCs and monocytes from CHB patients were MT or transfected for 6 or 24 h with 2 μg/mL of VACV ds 70mer. The transcripts of IFI16, STING, and IFN-β were then measured by qPCR
Discussion
Capable of recognition of pathogenic DNA, the innate immune defense system composed of intracellular DNA sensors plays an important role in preventing intracellular infection of pathogens such as DNA viruses. However, their role in HBV infection is still largely unclear. Published studies have shown that early in acute infection, HBV is poorly sensed by innate immunity and does not or weakly induce innate immune responses (12,31,41,42). It incubates stealthily in the liver for several weeks until elicitation of adaptive immune responses. Little is known about the involvement of DNA sensors in the process of acute and chronic HBV infection in humans. Collectively, our results reveal that (1) HBV might be sensed by DNA sensors in acute and chronic HBV-infected patients, (2) IFI16 expression is enhanced in PBMCs of both AHB and CHB patients and it is mainly expressed in monocytes, and (3) the IFI16-STING pathway in monocytes is attenuated by HBV infection.
The intracellular recognition of HBV infection by DNA sensors is still less understood. Moreover, it remains controversial in whether HBV can be sensed by intracellular DNA sensors or actively evades from the sensing pathways. To address this issue, we first analyzed the expression of STING-dependent DNA sensors in PBMCs of chronic HBV-infected patients. The results showed the expression of IFI16, DDX41, and MRE11 was enhanced in CHB, IT, and IA patients. Published data showed that IFI16 was closely related to the degree of liver inflammation in CHB patients (28). Upregulated expression of IFI16 only in chronic hepatitis patients suggests that it may be associated with inflammation within the liver. It was recently demonstrated that IFI16 could recognize and directly bind to HBV covalently closed circular DNA (cccDNA) through an interferon-stimulated response element (ISRE) within the cccDNA in hepatocytic nuclei (43). If this is the case, the enhanced expression of IFI16 in PBMCs occurs only when cccDNA is exposed in blood through inflammatory necrosis of hepatocytes, as HBV cccDNA only originates from hepatocytes, not from PBMCs (19). This may explain why IFI16 expression in PBMCs is upregulated only in acute or chronic hepatitis patients, but not in IT or IA groups that have normal ALT levels. DDX41 and MRE11 were reported to serve as cytosolic dsDNA sensors by Liu's and Kawai's groups, respectively (20,45). The DDX41 and MRE11 expression increased both in IT and IA groups, suggesting that they were involved in chronic HBV infection. Although there is still no report concerning the role of DDX41 and MRE11 playing in HBV infection, other reports have shown that they are involved in the control of virus infections (14,24,32). Thus, further investigation of their roles in HBV infection is warranted. Although activation of the cGAS-STING pathway was demonstrated to be able to inhibit HBV replication in cell culture and mice (7,15,18,40), HBV could also actively suppress cGAS expression and function in vitro and humanized liver chimeric mice (40). Our results showed cGAS expression level was not altered in chronic HBV patients, suggesting HBV may evade from cGAS sensing. In parallel with the increased expression of IFI16, DDX41, and MRE11, the adaptor STING expression was also enhanced in chronic HBV patients. However, the induction of the effector IFN-β was not observed in chronic HBV patients, implying the DNA sensor-STING pathway is suppressed. This may be explained by Liu's report showing HBV polymerase blocked DNA-sensing pathways by disrupting k63-linked ubiquitination of STING (26). Taken together, these data suggest HBV might be sensed by DNA sensors in PBMCs in chronic HBV-infected patients and support the notion that HBV plays an active role in evading and inhibiting innate immune responses (13).
Different from the expression profile of DNA sensors in chronic HBV infection, IFI16 was found to be the only upregulated DNA sensor in AHB patients, in accordance with a previous report that showed induced expression of the IFI16 gene in an acute HBV infection model of chimpanzees (41). However, the other DNA sensors were significantly downregulated as compared with HCs. This is reminiscent of the report by Dunn in which he demonstrated that with abundant production of IL-10 in AHB patients, the responses of NK cells along with CD4+ and CD8+ HBV-specific T cells from PBMCs were reduced (12). Interestingly, varicella patients and AHB patients displayed a similar expression pattern of DNA sensors, except IFI16. Despite with severe liver injury and a rapid short-term drop in HBV DNA levels in these AHB patients, the STING and IFN-β expression was not induced by acute HBV infection. The absence of a robust IFN-I response in AHB is in accordance with other studies (12,41). Of note, IFN-β expression in varicella patients was significantly suppressed as compared with HCs, which may be explained by the evidence that IFN-β expression is inhibited by VZV ORF47 kinase through preventing IRF3 activation in VZV-infected cells (39). Taken together, these data suggest that HBV might be sensed by IFI16 in acute HBV infection and utilize an immunosuppressive strategy to actively prevent innate immune responses.
Next, we determined the cellular source of IFI16. Early reports showed IFI16 was constitutively expressed in all CD34+ myeloid progenitor cells from human bone marrow and later exclusively in the CD14+ monocytoid lineage (8). IFI16 is present in peripheral blood monocytes and lymphocytes, but not in granulocytes. In lymphocytes, it is not regulated by IFN-α, IFN-γ, and IL6 (9). In 2010, IFI16 was first described as a STING-dependent dsDNA sensor in monocytes and macrophages by Bowie's group (38). Recently, Liang's group revealed that HBV was not sensed by the infected hepatocytes due to the lack of DNA sensing machinery but could induce human peripheral macrophage activation (6). Based on these data, we divided PBMCs into monocytes and nonmonocytes by magnetic separation. Quantitative RT-PCR analysis revealed that IFI16, whether in HCs, the CHB group, or the AHB group, is mainly expressed in monocytes. Furthermore, IFI16 expression in monocytes from both AHB and CHB patients was significantly upregulated as compared with that from HCs. These findings, along with the data from the aforementioned studies, indicate that HBV infection results in enhanced expression of IFI16 in monocytes of AHB and CHB patients.
Finally, we investigated the response of the IFI16-STING pathway in monocytes to VACV ds 70mer, a ligand for IFI16, to further confirm the impact of HBV infection on IFN-β expression. In concordance with the results from patients, the response of the IFI16-STING pathway to VACV ds 70mer was significantly attenuated in monocytes of CHB patients in comparison with HCs, suggesting that this DNA sensing pathway is impaired in CHB patients. These data further support the view that HBV is not always invisible to innate immunity but has evolved strategies to actively counteract recognition by innate immunity (13).
Our study has some limitations. First, the expression of DNA sensors in PBMCs may not be representative of the expression profile in liver. Therefore, studies of liver tissues should be included. Second, more cases of AHB patients are needed to verify the expression characteristics of these DNA sensors in AHB. Third, the response of the IFI16-STING pathway to VACV ds 70mer in monocytes from AHB patients was not assayed because of the difficulty of obtaining adequate specimens from AHB patients. Thus, additional studies are warranted. Finally, the detailed mechanism of suppression of the IFI16-STING pathway by HBV is needed to identify.
In summary, we investigated for the first time the role of STING-dependent DNA sensors in humans with acute and chronic HBV infection. Our data suggest that HBV might be sensed by DNA sensors in PBMCs of acute and chronic HBV-infected patients, and meanwhile HBV infection also attenuates the response of the DNA sensor-STING pathway in PBMCs and monocytes from CHB patients, which may facilitate the persistence of HBV infection. These findings increase our understanding of HBV immune pathogenesis and warrant further investigations to explore the profound mechanisms by which HBV suppresses the DNA sensing pathway.
Footnotes
Acknowledgments
We thank all the participants in this study. We also thank Dr. Yaying Zhou for flow cytometric analysis.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by the Committee of Innovation of Science and Technology of Shenzhen (No. JCYJ20150403101028209).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.