
Abstract
Hepatitis C virus (HCV) infection is a major worldwide problem with the highest incidence rates in Egypt. It affects B cells that serve as reservoirs for persistent HCV, resulting in phenotypic B cell alterations. Interleukin-7 (IL-7) is a cytokine with antiviral activity, important for B cell physiology. In addition, B cell-intrinsic toll-like receptor-7 (TLR7) signaling is required for optimal B cell responses during chronic viral infection, and the deficiency of TLR7 in B cells is sufficient to significantly impact antibody responses. Based on their known immunomodulatory effects, we hypothesized that direct-acting antiviral interferon-free therapy may affect TLR7 expression and the exhausted peripheral B cell compartment with the possibility of their restoration in patients who achieved a sustained virological response and their correlation to IL-7 level. This prospective study was accomplished on 80 Egyptian HCV patients and 75 controls. Frequencies of peripheral B cell subsets, TLR7 gene expression, TLR7 protein, and serum IL-7 levels were investigated by flow cytometry, quantitative polymerase chain reaction, and enzyme-linked immunosorbent assay, respectively. B cell subpopulations were exhausted and partially restored among HCV patients after receiving treatment, but not recovered with regard to activated mature or resting memory B cells. Almost all responders to direct antiviral drugs showed upregulation of TLR7 gene expression and correlated with the frequency of memory B cell, but not with IL-7. Moreover, IL-7 was not significantly different between groups although correlated with immature transitional B cells. Results may indicate the interplay between TLR7 and B cells during remission or progression of HCV. Thus, TLR7 could be used as a promising biomarker for assessment of antiviral treatment efficacy among chronically infected HCV patients, and that targeting TLR7 may be used as a potential prophylactic and/or therapeutic agent during chronic HCV as well as immune-potentiation of memory B cells.
Introduction
Chronic hepatitis C virus (HCV) infection is a major health burden worldwide with annual mortality due to HCV-associated complications of ∼700,000 deaths (69). Recently, the World Health Organization reported a global prevalence of chronic HCV infection with 71 million individuals (68). Egypt has launched promising strategies for HCV treatment using interferon (IFN)-free direct-acting antiviral (DAAs) regimens to reduce HCV prevalence from 11.9% to <2% by 2025. According to the Egyptian Health Issues Survey and the Egyptian Demographic Health Survey, there was a significant reduction in the overall seroprevalence of HCV in Egyptians from 14.7% to 10%, between 2008 and 2015 (35). Despite this progress, existing evidence indicates that Egypt exhibited the highest incidence rates compared with other countries (35), and HCV infection is still a national health problem. Also, the standard therapy of HCV remains expensive, with associated side effects and inconsistent cure rates (20).
HCV genotype 4 demonstrates an extraordinary predominance of 92.5% in infected Egyptians. The prevalence of genotypes 1–3 is 3.6%, 0.4%, and 0.2%, respectively, whereas genotype 5 is found in 0.1% and mixed infection with these five genotypes is found in 3.2% (54).
HCV is considered a lymphotropic virus and directly infects B cells that serve as a reservoir for persistent HCV infection (33), resulting in phenotypic alterations and dysfunction of B cell compartment (11,53). Infected B cells undergo proliferation, polyclonal hyperactivation, protein tyrosine phosphorylation, and hypermutation of immunoglobulin (Ig) genes. The consequences of this phenomenon are bystander activation of B cells, decreased numbers of memory B cells, hypergammaglobulinemia, and increased levels of activation markers on B cells (33,57). These molecular events may result in the viral escape, causing impairment of antiviral immune responses and/or inducing extrahepatic manifestations of HCV infection (57).
Moreover, memory B cells, which are phenotypically characterized as CD19+ CD27+, may be the main reservoirs of HCV because of their long life span and the upregulation of chemokine receptor (CXCR3), which contributes to their recruitment to the inflammatory sites in the liver to secure their places for persistent infection (33). Many studies have reported phenotypic changes of peripheral B cell pools during persistent human immunodeficiency virus (HIV-1) infections (4,47), which may be reversed after successful regimens of antiretroviral treatment or infection clearance (64).
Interleukin 7 (IL-7) is a cytokine with antiviral activity (55), important for B and T cell physiology, and produced by bone marrow (BM) and thymic stromal cells. In fact, the role of IL-7 in murine B lymphopoiesis has been extensively reported (34,63). Besides, some studies reported that IL-7 also induces B cell lymphopoiesis in cultures containing adult human BM stromal cells and cord blood hematopoietic precursor cells, indicating the importance of IL-7 receptor (IL-7Rα)-mediated signals for normal human B cell production (45). Recently, Zhang et al. suggested that IL-7 may regulate HCV-specific activated follicular helper T (Tfh) cells (72). In addition, IL-7R might be considered a potential therapeutic agent for persistent HCV infection and a useful marker of functional T cells in HCV (50) as it recovers their reactivity in vitro (27), and its expression correlates with the virologic outcome of acute HCV that might contribute to viral clearance (29). Moreover, the presence of IL-7R polymorphisms seems to be related to severe liver disease in HIV/HCV coinfected patients (49).
The correlation between the IL-7/IL-7R axis and immature transitional (IT) B cell subsets has been shown in some pathological conditions such as HIV and idiopathic CD4+ T lymphocytopenia (25,41); however, this association was not studied well in chronic HCV infection and is still unclear.
Toll-like receptor 7 (TLR7) (CD287) is a pattern recognition receptor encoded by the Xp22.2 chromosome and is expressed mainly in the endosomal compartment of B cells, plasmacytoid dendritic cells, and eosinophils. It contains leucine-rich repeats that mediate the recognition of pathogen-associated molecular patterns such as the HCV genome. The hallmark of this nucleic acid-sensing TLR is that it potently promotes the production of type I IFN and proinflammatory cytokines (37). B cell-intrinsic TLR7 signaling is essential for an optimal B cell response during chronic viral infection. Besides, the deficiency of TLR7 in B cells alone is sufficient to significantly influence antibody (Ab) responses (14).
It has been demonstrated that TLR7 expression is dysregulated in patients with chronic HCV infection, and this mechanism may enable HCV to evade the host innate immune response (51). Moreover, IL-7 and TLR7 induce synergistic T and B cell activation and Ig production in mice in the absence of B cell receptor crosslinking (8). They synergistically stimulate Th1 and Th17 cytokines (9) that are known to drive B cell activation (8), and upon viral triggering, they could be together involved in the early stages of B cell immunopathology.
So, clearly the understanding of the actual molecular basis of human B cell homeostasis, both at a balanced state and through chronic HCV infection, could help in unraveling HCV/B cell interactions. This will provide useful tools not only to avoid extrahepatic pathogenesis but also may have relevance for designing new functional HCV prophylactics and/or therapeutics. Subsequently, this study aimed to explore the alterations and the possibility of recovery (restoration or normalization) of peripheral B cell phenotypes among chronic HCV-infected Egyptian patients who received IFN-free DAA combination treatment. Our target was to study the immunomodulatory effects of these drugs on patients after achieving a sustained virological response at week 12 (SVR12) compared with their treatment-naive state, and if this is different from healthy controls. We also aimed to correlate B cell subpopulation with the expression of the innate immune receptor TLR7 as an intrinsic-signaling molecule required for optimal B cell response during chronic HCV, as well as with the serum level of IL-7 as a candidate for the acceleration of viral clearance during chronic viral infections (52), and its many roles in B cell survival, expansion, and differentiation (15).
Materials and Methods
Study participants
This study was conducted on 155 subjects who were seronegative for hepatitis B virus infection and HIV. They were divided into two groups matching in age and sex. Group 1 included 80 HCV patients examined before (pre-DAA) and after (SVR12) accomplishment of treatment with antiviral drugs (daily sofosbuvir [SOF, 400 mg] and daily daclatasvir [DCV, 60 mg] with/without daily weight-based ribavirin [RBV] 1,000 or 1,200 mg in patients <75 or ≥75 kg, respectively) for 12 weeks according to the HCV treatment regimens recommended by the European Association for the Study of the Liver guidelines (22).
All patients were followed up at week 12 by HCV polymerase chain reaction (PCR) for the response of treatment. They had negative serum HCV RNA at week 12 (SVR12) below the detection limit (<15 IU/mL). Viral loads were measured according to the manufacturer's protocol via the TaqMan real-time PCR HCV test kit (Roche Molecular System, Mannheim, Germany) using a fully automated COBAS AmpliPrep/COBAS TaqMan analyzer (CAP-CTM) system. Chronic HCV infection was diagnosed by anti-HCV Ab positivity and viremia with abnormal liver function tests for at least 6 months. Group 2 included 75 normal healthy individuals without HCV infection (30 males and 45 females) as a negative control group (mean ± standard deviation [SD] of age: 43.9 ± 10.9).
All relevant information of patients was collected, including personal and clinical data, such as age, sex, residence, a history of smoking, blood transfusion, surgical interference, hypertension, diabetes mellitus, chronic kidney disease, liver ultrasonography, and grading and staging of histological activity according to the pretreatment liver biopsy METAVIR score (6) (Supplementary Table S1). Laboratory assessment was carried out in pre-DAA and SVR12, including complete blood count and liver function tests (Table 1).
Comparison of Studied Laboratory Parameters Before and After Direct-Acting Antiviral Therapy
All data are expressed as mean ± SD and (range = Min.–Max.), range was preceded by median in abnormally distributed data only.
Bold p value: statistically significant at p ≤ 0.05.
Median of abnormally distributed data was compared using Mann–Whitney.
Mean ± SD of normally quantitative data was compared using Student t test.
pre-DAA: before treatment; SVR12: patients with sustained viral response after 12 weeks of treatment.
DAA, direct-acting antiviral; Ctl, control; ALT, alanine aminotransferase; AST, aspartate aminotransferase; HB, hemoglobin; PLT, platelets; PT, prothrombin time; RBC, red blood cell; WBC, white blood cell; MCV, mean corpuscular volume; MCH, mean corpuscular Hb; MCHC, mean corpuscular Hb concentration; SD, standard deviation; HCT, hematocrit.
Ethical considerations and exclusion criteria
All patients were selected from those attending the outpatient clinic of the Internal Medicine Unit of Medical Research Institute at Alexandria Main University Hospital. Before enrollment, ethical rules were approved by the Ethics Committee of the Medical Research Institute of Alexandria University (ethics board approval number: IRB No.: 00007555-FWA No: 00015859), and written informed consents were obtained from all participants before sampling in accordance with the guidelines of the 1975 Declaration of Helsinki.
Subjects with the following criteria were not included in the study. Pregnancy, age <14 years, immunosuppressive therapy within 6 months of the enrollment date, vaccination within 4 weeks before the blood draw, recent transfusion of blood or other blood derivatives, treatment with any anti-inflammatory medications, seropositivity to anti-HIV Ab and/or HBsAg, aviremia in seropositive anti-HCV Ab untreated patients, as well as a history of transplantation or malignancies. Exclusion criteria also included Child's C cirrhotic patients, hepatocellular carcinoma (HCC), and extrahepatic malignancy.
Peripheral blood mononuclear cell isolation
Peripheral blood mononuclear cells (PBMCs) from all subjects were isolated by centrifugation over Ficoll-Hypaque-1077 g/mL (Sigma-Aldrich; catalog number: 10771) solution by the density gradient method (26). The mononuclear cell layer was carefully aspirated, resuspended in phosphate-buffered saline (PBS), and then pelleted after adjustment to the appropriate number of cells by trypan blue exclusion method.
B cell immunophenotyping by flow cytometry
Frequencies of B cell subpopulations were analyzed by the four-color flow cytometry technique using a panel of specific mouse anti-human monoclonal antibodies (BD, Pharmingen™), peridinin chlorophyll protein (PerCP) CD19 (Clone: 4G7; isotype: mouse IgG1, κ), phycoerythrin (PE) CD10 (Clone: HI10a; isotype: mouse IgG1, κ), fluorescein isothiocyanate (FITC) CD21 (Clone: B-ly4, isotype: mouse IgG2a, κ), or allophycocyanin (APC) CD27 (Clone: M-T271; isotype: mouse IgG2a, κ) according to the direct immunostaining protocol (24). Nonspecific binding was determined using isotype control antibodies processed with each sample to validate the Ab specificity and to check for nonspecific background staining: a total of 20 μL of mouse IgG1-PerCP (Clone MOPC-21; cat. no. 559425), mouse IgG1-PE (Clone MOPC-21; cat. no. 555749), mouse IgG2a-FITC (Clone G155–178; cat. no. 555573), and mouse IgG2a-APC (Clone G155–178; cat. no. 551414) antibodies (all BD Pharmingen Biosciences) were added into the control tube in all cases. A dot plot of forward scatter cells (FSC) versus side scatter cells was used to identify total lymphocytes and exclude monocytes and dead cells from stained whole blood.
A plot of PerCP CD19 versus FSC was used to gate at least 5,000 events from the total gated peripheral lymphocytes that represented B cell fraction. From gated B cells, IT B cells were identified as CD19+ CD10+ CD21− CD27− by blotting PE CD10 versus APC CD27, while the gated CD19+ CD10− B cells were identified by FITC CD21 versus APC CD27 dot plot and dissected in naive (NAI) B cells (CD19+ CD10− CD21+ CD27−), activated memory (AM) B cells (CD19+ CD10− CD21−/low CD27+), resting memory (RM) B cells (CD19+ CD10− CD21+ CD27+), and tissue-like memory (TLM) B cells (CD19+ CD10– CD21– CD27–) according to the threshold established using the negative isotype control antibodies (73). Also, total memory B cells were identified as CD19+CD27+.
Daily machine calibration of the instrument and fluorescence compensation were performed using standardized fluorescence beads (CaliBRITE beads; BD Biosciences) with FACSComp software (BD), and compensation was also performed using single-stained normal peripheral blood samples as appropriate to allow determining the levels of compensation. Unstained sample was included to look at autofluorescence. Cells were tested by BD FACSCalibur™ flow cytometry (Becton-Dickinson) and were analyzed using BD CellQuest™ Pro software for windows 7 by an investigator who was blinded to the patients' clinical and laboratory data.
IL-7 and TLR7 quantification by enzyme-linked immunosorbent assay
Quantitative determination of concentrations of serum human IL-7, as well as TLR7 in cell lysates, was assayed in blood samples of HCV patients before antiviral therapy (pre-DAA), 12 weeks after starting treatment (SVR12), and from the control group using the Quantikine® HS enzyme-linked immunosorbent assay (ELISA) Human IL-7 Immunoassay kit (R&D Systems, Inc.) and TLR7 ELISA assay kit (MyBioSource, Canada), respectively, according to the manufacturer's instructions. The cell lysates from PBMCs (2 × 106) were used for TLR7 protein quantification. Cell suspension was diluted with PBS until cell concentration reached 1 × 108/mL, stored overnight at −20°C, then subjected to two freeze/thaw cycles to break up the cell membranes, and finally centrifuged for 5 min at 5,000 g, 2–8°C to collect the supernatant. The optical density (O.D.) of each well was determined within 30 min, using a microplate reader set (BIO-TEK® ELx800 UV™ Universal ELISA Microplate Reader) to 450 nm. The absorbance of each well was read on a spectrophotometer using 450 nm. Standard curves were generated for each of IL-7 and TLR7, and the mean O.D. in each well was converted into a concentration using the linear portion of the standard.
Assessment of TLR7 gene expression by quantitative real-time PCR
Total RNA extraction from PBMCs
Total cellular RNA from human PBMCs adjusted to 5 × 106 cells/mL (1.3 mL of cell suspension containing 6,500,000 cells, which equals 6,500/μL) was extracted by the QIAamp® RNA Blood Mini kit (QIAGEN) following the manufacturer's instructions, where PBMCs were pelleted after adjustment to the appropriate number of cells by centrifuging for 10 min at 1,800 rpm in a centrifuge tube. All supernatants were carefully removed by aspiration. Three hundred fifty microliters of RNeasy Lysis Buffer (RLT) was added to each sample in a QIAshredder spin column, mixed to homogenize the lysate. Three hundred fifty microliters of 70% ethanol was added to the homogenized lysate, and 700 μL of the sample was pipetted into a new QIAamp spin column and centrifuged for 15 sec at ≥10,000 rpm. Samples were washed with 700 μL of RNeasy Wash Buffer (RWT) and then 500 μL of RNeasy Pre-Ethanol Wash Buffer (RPE). The QIAamp spin column was transferred into a 1.5 mL collection tube and 30–50 μL of RNase-free water was pipetted directly onto the QIAamp membrane. The extract was centrifuged for 1 min at ≥10,000 rpm to elute. This step was repeated once. The purity (quality), concentration, and quantity of RNA were assessed using a NanoDrop™ 2000 UV-Vis spectrophotometer, RNA amounts and purity were determined by O.D. measurements at 260 and 280 nm, and the ratio A260/A280 in the UV-spectrophotometer (Photobiometer, Eppendorf, Germany). The RNA extract was dispensed in aliquots and stored at −80°C for the preceding work.
TLR7 gene quantification using SYBR® Green real-time PCR
All extracted RNA samples from subjects were used as templates for preparation of cDNA sequences using the commercially available kit (QuantiTect® Reverse Transcription Kit) using a conventional PCR thermocycler (Biometra® TProfessional Basic PCR Thermocycler) Each reaction volume includes the following: 2 μL gDNA Wipeout Buffer, 10 μL Template RNA (a total of 1 μg of RNA), and 2 μL RNase-free water. Template RNA (14 μL) was added to each tube containing reverse-transcription master mix, mixed, and then stored on ice. The reaction components were incubated for 15–30 min at 42°C and then incubated again for 3 min at 95°C to inactivate Quantiscript Reverse Transcriptase. The products were analyzed on 1% agarose gel stained with a green viewer. Reverse-transcription reactions were stored at −20°C for further use in real-time PCR.
The TLR7 gene expression in reverse-transcribed cDNA was quantified via the SYBR Green real-time PCR protocol using the QuantiTect SYBR Green PCR kit according to the manufacturer's instructions. The gene expression of the target gene, human TLR7-specific ready-made human primers QuantiTect Primer Assay (Assay name: Hs_TLR7_1_SG, Cat. No.: QT00030030), was normalized to human glyceraldehyde-3-phosphate dehydrogenase (GAPDH) as an internal control gene (Assay name: Hs_GAPDH_1_SG, Cat. No.: QT00079247). Samples were amplified with GAPDH primers for determination of the initial relative quantity of cDNA in each sample, and all PCR products were normalized to that amount. Negative controls (without template) were produced for each run.
The quantitative, real-time PCR experiment was conducted using the Rotor-Gene Q thermocycler (Qiagen). Each PCR mixture included 12.5 μL 2 × QuantiTect SYBR Green PCR Master Mix, 2.5 μL TLR7 or GAPDH primers (in separate tubes), 2 μL RNase-free water, and 8 μL Template cDNA to reach a total reaction volume of 25 μL. Thermal profile was as follows: 95°C for 15 min (PCR initial activation step), followed by 40 cycles of three-step cycling (denaturation at 94°C for 15 sec, annealing at 50–60°C for 30 sec, and extension at 72°C for 30 sec.
Data were analyzed by Rotor-Gene Q Series software (QIAGEN Company) and the fully automated COBAS AmpliPrep/COBAS TaqMan analyzer (CAP-CTM) system Sysmex CA-1500 System (Siemens, Germany). The melting curve of gene expression that was used in quantitative PCR experiment for both target gene (TLR7) and the normalizer gene (GAPDH) is illustrated in Supplementary Figure S1. The relative quantification (RQ) of the target (TLR7) mRNA was normalized to the level of the internal control, and GAPDH mRNA level by the comparative 2−ΔΔCt method. Cycle threshold (Ct) values were recorded. Data were transformed and relatively compared using the comparative Ct method (60).
Statistical analysis
Data were statistically analyzed using Statistical Package for the Social Sciences (SPSS) software version 20 (Chicago, IL). Categorical variables were expressed as numbers and percentages and the chi-square test was used to compare different groups represented as qualitative data. The Shapiro–Wilk test was used to determine whether quantitative data were normally distributed (represented as mean ± SD) or non-normally distributed (represented as median and range). Student t-test and one-way analysis of variance test were used to assess the statistical significance of independent normally distributed quantitative data of two groups and more than two groups, respectively. Mann–Whitney and Kruskal–Wallis tests were used for statistical comparison of non-normally distributed data. Pearson's correlation coefficient (r) and Spearman's coefficient were used to investigate correlations of normally distributed and non-normally distributed data, respectively. A receiver operating characteristic (ROC) curve analysis was performed to assess the performance value of TLR7 gene expression as a valuable prognostic biomarker of chronic HCV infection, and an operational cutoff level was determined for distinguishing pre-DAA chronic HCV infection from the SVR12 group. p-Values <0.05 were considered statistically significant.
Results
Alterations in a number of peripheral B cell subpopulations in pre-DAA HCV patients
As summarized in Table 2 and Figure 1, the frequencies of CD19+ B cells in peripheral blood were insignificantly different between controls (mean ± SD = 16.14 ± 6.09%) and pre-DAA HCV patients (mean ± SD = 17.32 ± 1.45%; p = 0.879). IT B cells (CD10+ CD27−) showed low frequencies in control subjects (mean ± SD = 9.92 ± 5.31%) compared with pre-DAA HCV patients, who showed a higher frequency of this B cell subset in their peripheral blood (mean ± SD = 14.24 ± 5.65%; p = 0.035). In contrast, total memory B cells (CD19+ CD27+) showed a significant percentage in the control group (mean ± SD = 28.5 ± 10.64%) compared with the treatment-NAI HCV patients (mean ± SD = 21.9 ± 5.61%; p = 0.034).
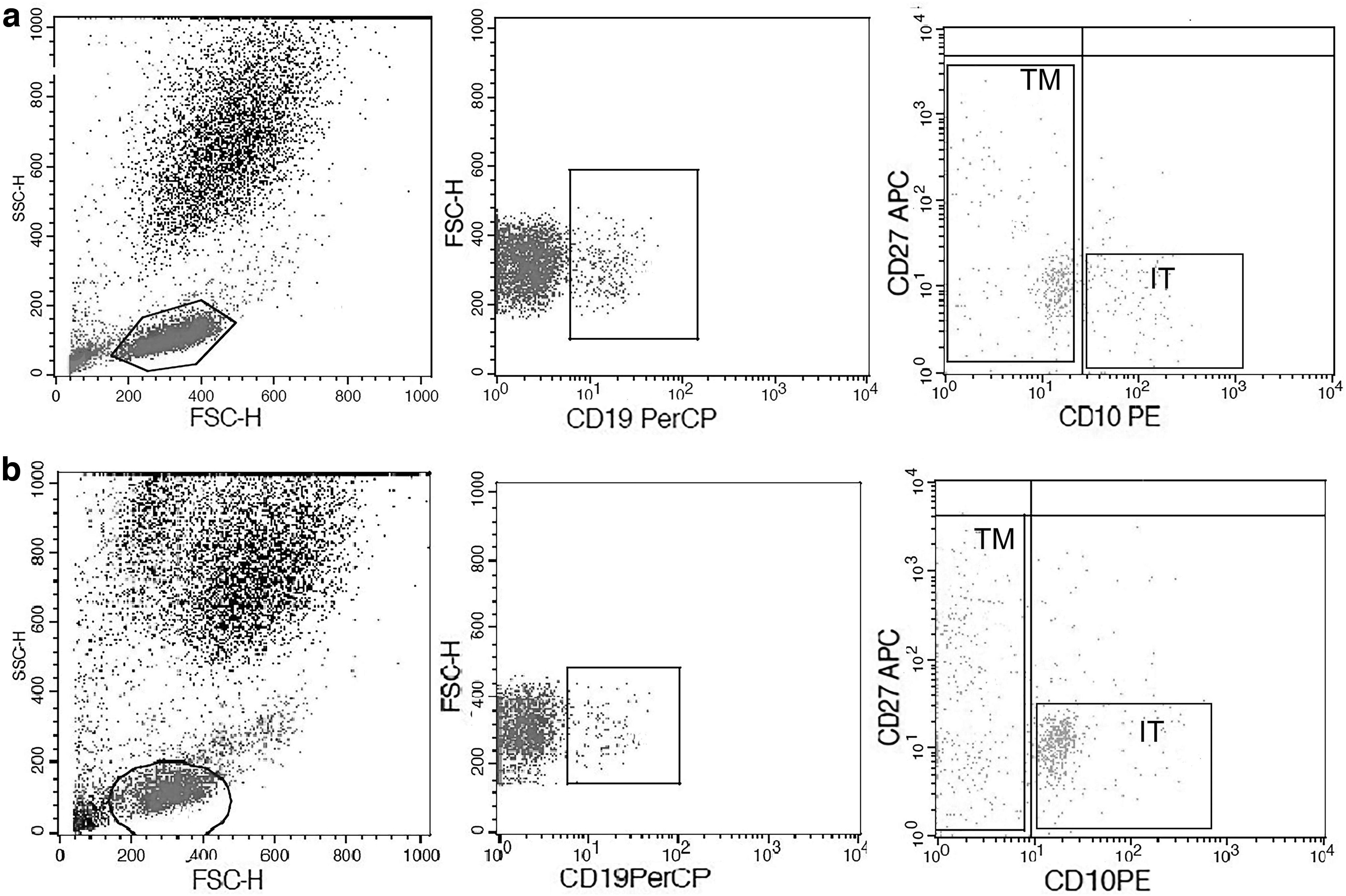
Alterations in percentages of B cell subsets in untreated pre-DAA HCV-infected patients (No. = 80): A dot plot of FSC versus SSC was used to identify total lymphocytes and exclude monocytes and dead cells from stained whole blood. A plot of PerCP CD19 versus FSC was used to gate at least 5,000 events from total gated peripheral lymphocytes that represented B cell fraction. From gated B cells, IT B cells were identified as CD19+ CD10+ CD21− CD27− by blotting PE CD10 versus APC CD27 and showed higher frequencies in pre-DAA HCV patients than in control subjects. TM B cells were identified as CD19+ CD27+ and were decreased in pre-DAA HCV patients than controls.
Frequencies of Peripheral B Cell Subsets in the Studied Groups
All data are expressed as mean ± SD and (Min.–Max.) and (range = Min.–Max.), range was preceded by median in abnormally distributed data only.
Bold p value: Statistically significant at p ≤ 0.05.
Median of abnormally distributed data was compared using Mann–Whitney.
Mean ± SD of normally quantitative data was compared using Student t test.
pre-DAA: before treatment; SVR12: patients with sustained viral response after 12 weeks of treatment.
IT, immature transitional; AM, activated mature; RM, resting memory; TLM, tissue-like memory; NAI, naive.
Among the CD10− CD21+ B cells, it was obvious that the control subjects had more RM B cells (CD21+ CD27+) than pre-DAA HCV patients; p = 0.007. However, control subjects and pre-DAA patients had similar percentages of CD21+ CD27− NAI B cells in their peripheral blood; p = 0.610.
Concerning the median of activated mature (AM) B cells (CD21−/low CD27+), it was lower in the control group (median [range] = 5 [2–15] %) than that in the HCV patients before treatment (median [range] = 9 [3–15] %); yet, a nonsignificant difference existed; p = 0.167. Likewise, the median of CD21− CD27− TLM B cells was significantly lower in the control subjects (median [range] = 8.5 [4–19] %) when compared with the pre-DAA patients (median [range] = 18 [13–27] %; p < 0.001).
Recovery of a number of B cell subsets in DAA-treated HCV patients
So far, our findings revealed profound phenotypic alterations in the B cell compartment of HCV patients when compared with healthy non-HCV individuals. We then investigated the effect of DAA on B cell subpopulations in the HCV individuals after they experienced an undetectable HCV viral load (<15 IU/mL) after the last 12 months of therapy (SVR12) and retained an alanine aminotransferase (median = 20.25 IU/L; p < 0.001) and aspartate aminotransferase (median = 22.5 IU/L; p < 0.001).
Gating strategies that were followed to represent peripheral B cell subpopulations among healthy control, DAA-NAI HCV patients (pre-DAA), and SVR12 are illustrated via a dot plot of flow cytometry in Figure 2. Compiled data from all studied individuals indicated that the percentage of IT cells decreased in patients after receiving treatment (mean ± SD = 11.45 ± 5.02%) versus (mean ± SD = 14.24 ± 5.65%) in pre-DAA, but with a nonsignificant difference (p = 0.233). However, when comparing SVR12 patients with controls, the percentage was also nonstatistically different (p = 0.638).
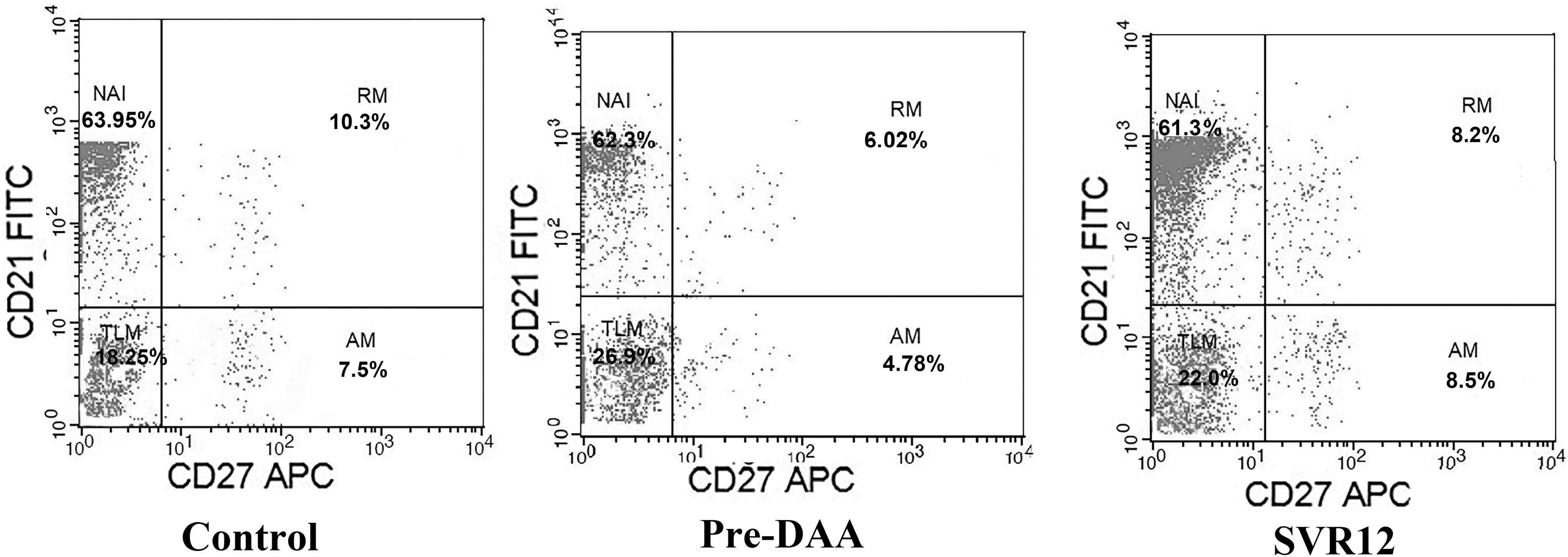
Effect of DAA therapy on memory B cell subset distribution in HCV-infected patients. Percentages of B cell subsets were determined by four-color flow cytometry analysis of CD19, CD21, CD27, and CD10 surface expression molecules. Profiles of expression of CD21 and CD27 are shown for one representative of each group. Cells were gated according to the lymphocyte forward and side scatter pattern and the CD19+ cells. The gated CD19+ CD10− B cells were identified by FITC CD21 versus APC CD27 dot plot and dissected in NAI B cells (CD19+ CD10− CD21+ CD27−) (upper left panel), RM B cells (CD19+ CD10− CD21+ CD27+) (upper right panel), AM B cells (CD19+ CD10− CD21−/low CD27+) (lower right panel), and TLM B cells (CD19+ CD10− CD21− CD27−) (lower left panel). Control (No. = 75), pre-DAA (No. = 80), and SVR-12: (No. = 80). NAI, naive; FITC, fluorescein isothiocyanate; RM, resting memory; AM, activated memory; TLM, tissue-like memory.
After achieving SVR12, HCV patients expressed more frequencies of total memory B cells than these frequencies before treatment ([mean ± SD = 25.55 ± 7.30%] vs. [21.9 ± 5.61%] respectively), although this increase was not statistically significant (p = 0.337); however, it was also insignificantly different from healthy subjects (p = 0.489).
At the same time, the median percentage of RM B cells was significantly elevated in SVR12 HCV patients (median [range] = 17.5 [8–39] %) than in pre-DAA (median [range] = 13[6–21] %; p = 0.027), but the increased level was not normalized and was still significantly lower than that in the control group (median [range] = 22.5 [6–42] %; p = 0.038). In addition, SVR12 patients sustained nonsignificant AM B cells from that measured in their pre-DAA (median [range] = 10 (5–18) % vs. 9 [3–15] %; p = 0.639). On the contrary, this subset was statistically different comparing SVR12 patients with controls, p = 0.005.
Intriguingly, the percentage of TLM B cell subpopulation was significantly lower in DAA-treated (median [range] = 13 [5–22] %) than untreated HCV patients (18 [13–27] %; p = 0.02), and not different than the healthy group, p = 0.067 (Fig. 3). Finally, we observed also a similar NAI B cell percentage between SVR12 HCV patients (mean ± SD = 62.1 ± 8.45%) and both pre-DAA HCV (mean ± SD = 60 ± 6.40; p = 0.749) and controls (62.75 ± 11.76%; p = 0.973).

Percentages of TLM B cells using flow cytometry: differences between groups were evaluated using Mann–Whitney U test, and the level of significance was set at *p ≤ 0.05. Number of HCV patients: 80, Number of control: 75. Data represented as means ± SDs and were summarized as dot plot. Each dot represents the expression percentage of TLM B cells for one individual. SD, standard deviation.
Decreased TLR7 gene expression and circulating levels in treatment-NAI HCV patients and their restoration after DAA therapy
To examine the involvement of TLR7 in chronic HCV infection, we studied TLR7 protein levels and mRNA expression in PBMCs. We found that TLR7 gene expression in healthy individuals was almost 1.31-fold higher than those in treatment-NAI HCV patients (Table 3; Fig. 4). Simultaneously, TLR7 levels in the cell lysates of healthy subjects were obviously higher compared with those in treatment-NAI HCV patients (median [range] = 6.82 [5.2–8.54] ng/mL vs. 2.88 [0.198–3.79] ng/mL; p = < 0.001).
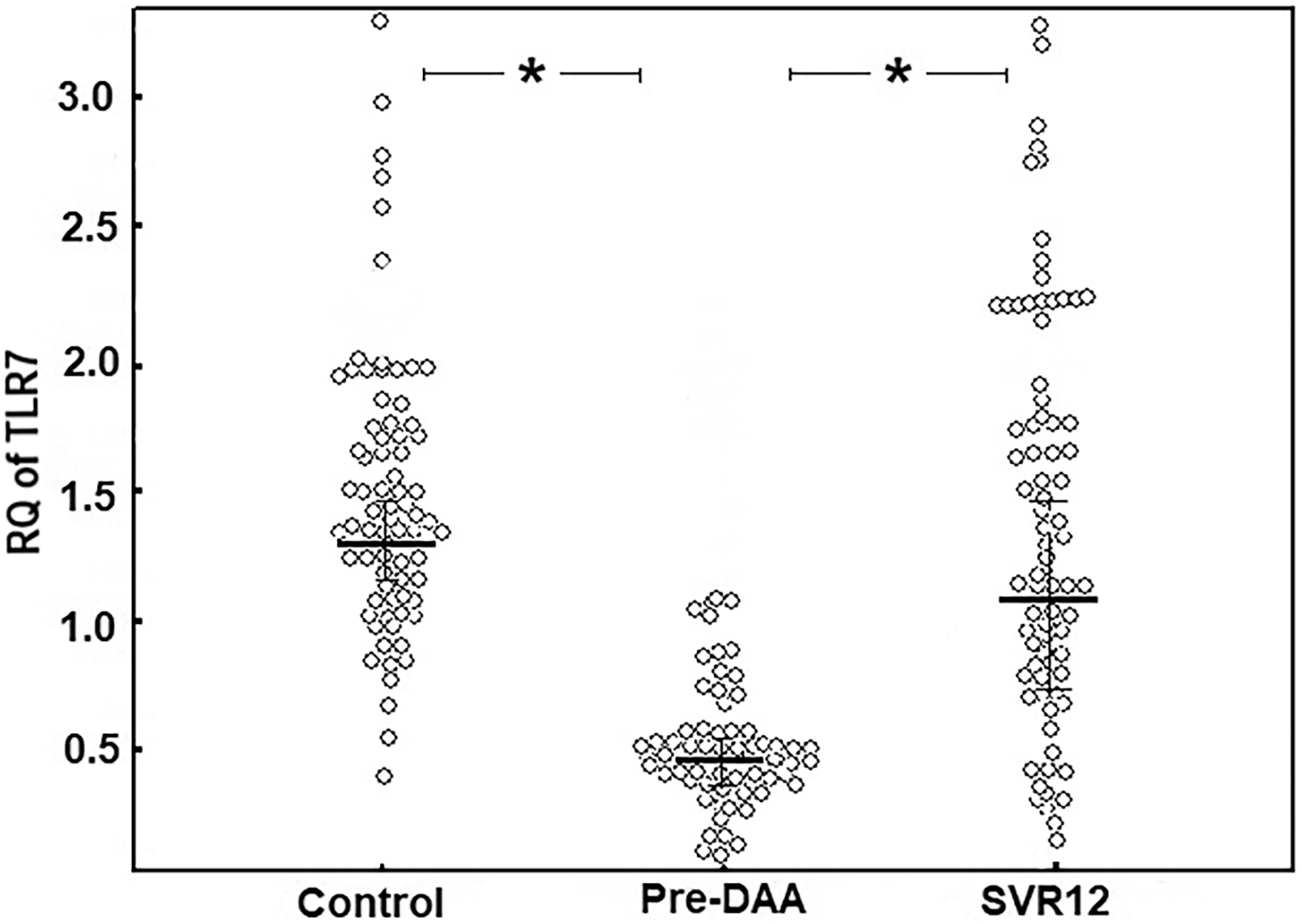
Comparison between gene expression of TLR7 in HCV patients (pre- and post-DAA) and controls using quantitative polymerase chain reaction. Data represented as means ± SDs and were summarized as dot plot. Differences between groups were evaluated using Mann–Whitney U test; each dot represents the mRNA expression of TLR7 for one individual. Number of HCV patients: 80, Number of control: 75. The level of significance was set at *p ≤ 0.05. TLR7, toll-like receptor-7.
Toll-Like Receptor-7 and Interleukin-7 Among Different Studied Groups
All data are expressed as mean ± SD and (Min.–Max.) and (range = Min.–Max.), range was preceded by median in abnormally distributed data only.
Bold p value: Statistically significant at p ≤ 0.05.
Median of abnormally distributed data was compared using Mann–Whitney.
Mean ± SD of normally quantitative data was compared using Student t test.
pre-DAA: before treatment; SVR12: patients with sustained viral response after 12 weeks of treatment
TLR7, toll-like receptor-7; IL-7, interleukin-7; RQ, relative quantification.
Untreated patients with fibrosis stage of F1 + F2 (No. = 37) tended to have a significantly decreased gene expression of TLR7 (0.28 ± 0.14 fold) versus patients with no fibrosis (F0: No. = 43) with (0.58 ± 31) (p = 0.048), which may implicate a role of TLR7 in the progression of the disease. HCV patients (F1 + F2) also tended to have a lower level of TLR7 (2.22 ± 0.43 ng/mL), while the level was increased in F0 patients (3.02 ± 0.15 ng/mL), but without a significant difference.
To determine if the change in TLR7 expression is associated with the increase in certain B cell subtypes, correlation analyses were carried out. We noticed that TLR7 had a direct correlation to the frequencies of total memory B cell subset among pre-DAA HCV patients (r = 0.482, p = 0.031), and after treatment (r = 0.447, p = 0.048) (Fig. 5). On the contrary, an interesting nonsignificant negative correlation was noticed with NAI B cells (r = −0.403, p = 0.078), and no correlations with other studied B cell subtypes, including IT B cells (r = 0.203, p = 0.391).

Correlation analysis between frequencies of peripheral total memory B cell subset and RQ of TLR7 among
DAA therapy was observed to affect both gene expression and protein level of TLR7. The RQ of TLR7 gene expression showed a statistically significant increase among SVR12 HCV patients after successful DAA therapy (p = 0.036) (Fig. 4). Moreover, the TLR7 protein level in PBMCs was also upregulated in treated HCV patients compared with its level before treatment, and TLR7 was significantly increased (p = 0.02). Comparing the medians of both TLR7 mRNA and protein levels in controls and SVR12, no significant differences were obtained (p = 0.893 and p = 0.777, respectively).
ROC curve analysis of TLR7 gene expression as a prognostic marker in chronically infected HCV patients
ROC analysis was used to assess the performance value of TLR7 gene expression as a valuable prognostic biomarker of chronic HCV infection (Table 4; Fig. 6). At a cutoff value of ≤0.42, TLR7 gene expression showed a highly significant (p = 0.01) prognostic value for responsiveness to treatment, 65% sensitivity, 85% specificity, and area under the curve of 0.813, with 95% confidence interval ranging from 0.68 to 0.95.
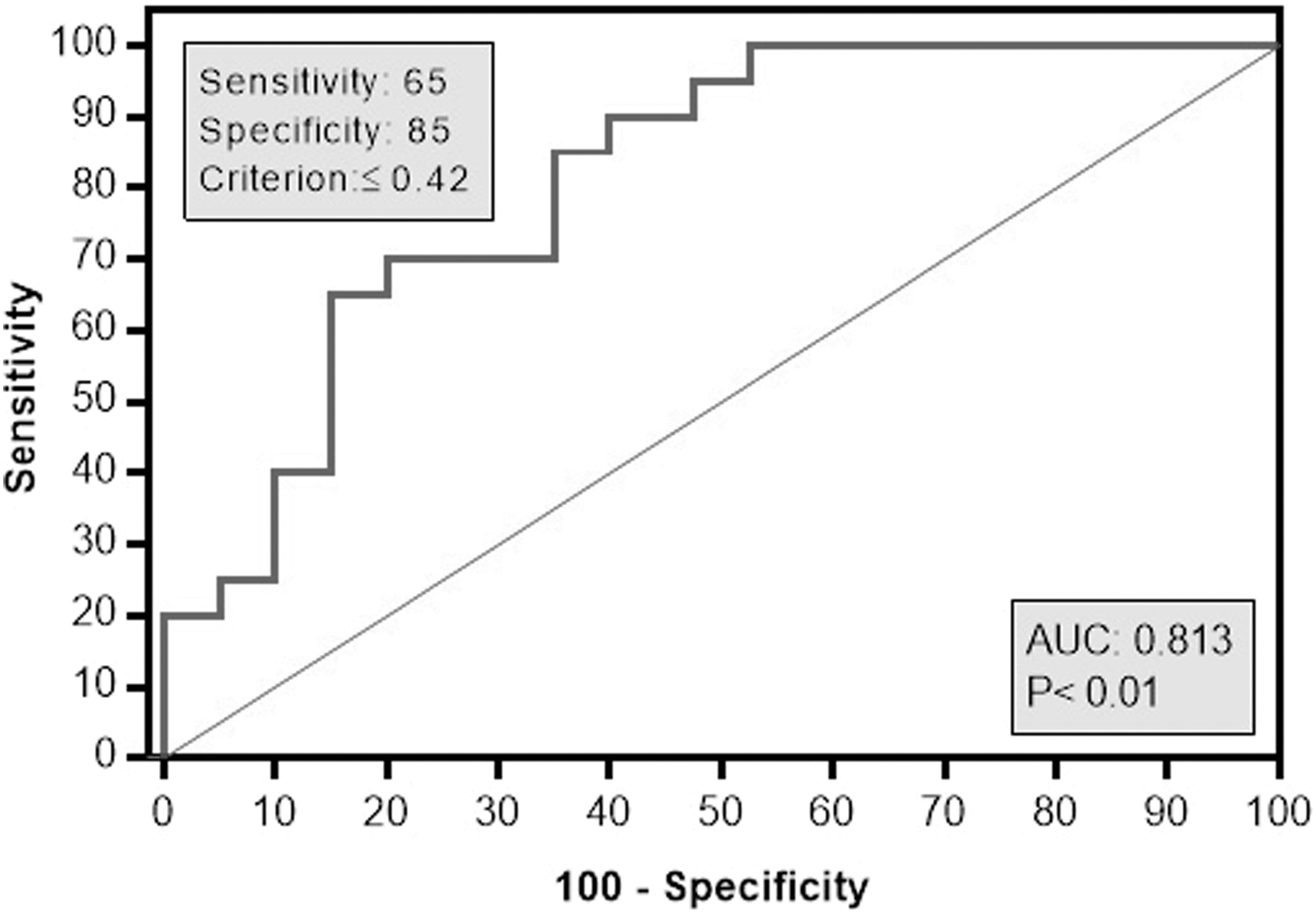
Receiver operator characteristic curve analysis for RQ TLR7 to predict responder cases. At a cutoff value of ≤0.42, TLR7 gene expression showed 65% sensitivity, 85% specificity, and area under the curve of 0.813, with 95% confidence interval ranging from 0.68 to 0.95 for distinguishing responders, showing highly significant statistical difference (p < 0.001).
Agreement (Sensitivity, Specificity) of Relative Quantification of Toll-Like Receptor-7 to Predict Responder Cases
Bold value statistically significant at p ≤ 0.05.
LL, lower limit; UL, upper limit; CI, confidence interval; AUC, area under the curve; PPV, positive predictive value; NPV, negative predictive value.
Serum level of IL-7
The serum level of IL-7 showed a nonsignificant increase in pre-DAA patients compared with control (p = 0.569), as represented in Table 3 and Figure 7. Among HCV patients, the IL-7 serum levels were estimated, and moreover, the variations in IL-7 between pre-DAA (mean ± SD = 10.52 ± 3.05 pg/mL) and SVR12 (mean ± SD = 9.11 ± 3.49 pg/mL) were nonsignificant, p = 0.355. Moreover, we did not notice any variation in IL-7 serum levels among pre-DAA patient characteristic subgroups (fibrosis stage) ([F0: No. = 43; IL-7 = 10.01 ± 3.87 pg/mL] vs. [F1 + F2: No. = 37; IL-7 = 11.21 ± 2.99 pg/mL]; p = 0.786). Also, IL-7 was not correlated to viral load (r = −0.135, p = 0.917).

Serum IL-7 level: serum levels before and after 12 weeks of DAA therapy between HCV patient (No.: 80) and control (No.: 75) groups were measured by ELISA. Comparison between groups was evaluated using Student t test. Data represented as means ± SDs and were summarized as bar chart with error bar. Error bars represented standard error of the mean. The level of significance was set at p ≤ 0.05. IL-7, interleukin-7; ELISA, enzyme-linked immunosorbent assay; ns, not significant.
Correlation analysis between IL-7 and TLR7 expression showed a nonsignificant relation in any of the studied groups (control [r = 0. 0.389, p = 0.091], pre-DAA [r = 0.087, p = 0.714], and SVR12 [r = 0.025, p = 0.915]). This may be due to no considerable IL-7 levels produced by activation of the TLR7 signal pathway (2). However, the obtained results showed a positive correlation between serum IL-7 levels and frequencies of immature/transitional B cell subsets among untreated HCV patients (p = 0.048) (Fig. 8).

Correlation analysis between frequencies of immature transitional B cell subset (flow cytometry) and serum IL-7 level (ELISA) in pre-DAA HCV patients (No.: 80): correlation was conducted by Pearson correlation coefficient. The level of significance was set at *p ≤ 0.05.
Discussion
During extrahepatic HCV infection, envelope 1 (E1) proteins of HCV interact with the surface CD81 receptor of B cells to be used by HCV as reservoirs to replicate (33). Consequently, HCV-infected B cells undergo phenotypic alterations of peripheral B cell pools (62), which were reported in chronic HCV infection in several studies (11,18).
The pathogenic role of HCV has been further demonstrated by studies in which the clinical resolution of B cell dysfunctions was observed after successful regimens of IFN-anti-HCV treatment (43). Egypt has established ambitious national HCV prevention and IFN-free treatment programs aiming to achieve the total elimination of the virus by 2030 (35). However, the standard therapy of HCV remains expensive and the treatment eligibility is still low (21). Furthermore, modulation of the immune response against HCV was reported during IFN-free DAA regimens in immune cells other than B cells, such as CD8+ and CD4+ T cells, and natural killer cells (67). Yet, the immunologic dynamics of HCV-infected patients who were treated with direct acting antiviral drugs, as immunomodulators, is poorly understood in Egyptian patients.
NAI B cells (CD21+CD27−) constitute the largest B cell population in the blood of healthy individuals, followed by memory B cells. Generally, memory B cells are of different classes that have considerable functional differences: resting, activated, and TLM B-cells (39). Our results revealed a decrease in total memory B cells in untreated HCV patients, which were significantly elevated after treatment. This decrease in memory B cells in pre-DAA patients was obviously accompanied by a decrease in TLR7 expression, which was also restored after treatment. Human memory B cells express significant levels of TLR7, while NAI B cells express low levels of TLR7 (2), which were reflected by our correlation analysis. TLR7 expression in human B cells is strongly upregulated by type I IFN as well as by stimulation through the BCR, CD40, or cytokine receptors (2). The recovery of memory B cell subset in SVR12 was not accompanied by a significant change in IL-7 in SVR12 patients, which might suggest an inferior role of IL-7 in maintaining B cell homeostasis in HCV patients than that of DAA therapy.
Possible causes of the decrease of peripheral memory B cells in pre-DAA patients include (i) increased differentiation of peripheral AM B cells into short-lived plasmablasts; (ii) intrahepatic and/or lymphoid compartmentalization of peripheral AM B cells causing upregulation of homing chemokine receptors CXCR3 (17); and (iii) increase of activation-induced B cell apoptosis (58); however, contradicted by studies proposing that B cells among chronically HCV-infected patients are relatively intrinsically resistant to apoptosis (13,18). Moreover, the disappearance of memory B cells was correlated in a study by Doi et al. with impairment of B cell activation and decreased function of B cells as antigen- presenting cells (17).
An alternate explanation is the conversion of AM B cells into a unique B cell population, atypical, exhausted, or TLM B cells, recently introduced as CD21−/low CD27− B cells and coinciding with evidence of B cell anergy (56). This nonconventional memory B cell type, which exists normally at low frequency in the peripheral blood of healthy individuals, expresses multiple homing (CD11c and CXCR3) and inhibitory receptors (inhibitory receptor Fc receptor-like-4 [FCRL4]) (36), and proliferates poorly in response to B cell stimuli (18). They are reported to be increased in immunosuppressive diseases such as HIV (4); however, TLM B cells are not unique to HIV infection and have been described in several chronic inflammatory conditions (56).
Three pathways were proposed for generation of TLM B cells, including prolonged Ag-mediated hyperactivation of previously functional B memory cells during persistent stimulation as in viral infection (5,61), memory B cells derived from prematurely aborted germinal centers or from extrafollicular differentiation pathways, and loss of survival niche that may drive the expansion of a phenotypically and functionally altered memory B cell subsets (61). Apparently both inflammation and chronic immune stimulation are thought to induce the proliferation of TLM B cells that are not able to produce Abs due to a defect in affinity maturation and somatic hypermutation (44). This might clarify the lack of acquisition of immunity against HCV during chronic infection, the IgG1 restriction, and the low-titer and delayed-onset Ab responses (11)
Data of the present work show that TLMs were highly increased in pre-DAA HCV patients. This percentage was normalized after treatment versus the control group, with no significant difference in between.
While total memory B cells and TLM B cells were fully recovered in HCV patients after treatment, current work shows that the frequency of AM subsets among treated HCV patients was not completely normalized or restored and should be monitored after 12 SVR. This finding is in the line with Liu et al. who reported an increased frequency of AM B cells and serum IgG levels in untreated chronic HCV patients compared with healthy control (40). AM B cells are the kind of memory B cells that function as effective antigen-presenting cells to naive T cells, highly activated and prone to extrinsic apoptosis (30).
In contrast, the frequencies of RM B cells were significantly decreased in pre-DAA chronic HCV patients relative to the control group due to the depletion of the classic costimulatory receptor CD27-expressing RM that have been also reported in HIV-infected individuals (48). This loss in RM B cells was characterized in chronic HCV regardless of cirrhosis or HCC, and as early as during the acute phase of HCV (11).
IT B cells are short-lived and functionally immature cells, which are transported to the spleen, where they develop into long-lived mature B cells (28). Holz et al. proposed that increased IT B cells among HCV-infected patients may be due to a secondary egress from the BM to compensate for the loss of peripheral B cell pool via elevation of serum B cell activation factor (31). A role of IL-7 in IT cell composition is suggested, as evidenced by the direct correlation between IL-7 serum level and IT cell frequencies in untreated patients, which was also suggested in chronic HIV infection (42). Results of increased IT B cells in our pre-DAA studied group partially coincide with other studies (39), but contradict the findings of Holz et al. who found no correlation between frequencies of peripheral IT B cells and serum IL-7 levels, which was also supported by Sugalski et al. (31,62). The observed reduction in the frequency of this cell after SVR12, although nonsignificant, suggests a possible involvement of this cell in HCV immunopathogenesis. IT B cells might act as an alternate Epstein–Barr virus (EBV) reservoir in HIV-infected patients with a high EBV viral load and a strong depletion of RM B cells (11).
Upon engagement with specific ligands, TLR7 initiates signal transduction through the cytosolic adaptor MyD88 and induces both type I IFN and an inflammatory response against HCV (71). Beside its antiviral activity against HCV, TLR7 expression in human B cells mediates B cell proliferation and differentiation, as well as secretion of cytokines and chemokines (7), and there are many synthetic TLR7/8 agonists used as vaccine adjuvants (66). Herein, this study observed that the expression of TLR7 was significantly downregulated among pre-DAA HCV patients compared with the expression in the healthy control group, which partially coincides with previous studies (46).
In fact, the downregulation of TLR7 gene expression is considered a strategy of immune evasion by HCV, and it has been suggested to play a key role in viral persistence and development of chronicity (51). Studies reported that TLR7 gene expression in peripheral blood even becomes undetectable among HCV-infected patients with end-stage liver diseases (1). Chang et al. reported that the downregulation of TLR7 gene expression may be due to HCV specifically targeting TLR7 gene expression, which interferes with transcription, posttranscriptional modifications, and instability of TLR7 mRNA by ubiquitin machinery to degrade TLR7 mRNA, production of small interfering RNAs targeting TLR7 mRNA, exploitation of host regulatory factors that bind to TLR7 mRNA, or usage of viral proteins to support the instability of TLR7 mRNA (12). In addition, HCV infection downregulates the gene expression of TLR7 via upregulation of micro (mi)RNA-758 to reduce expression of type I IFN (IFN-α and IFN-β) (70).
In contrast, the results presented in the current study contradicted with other studies reporting an upregulation of TLR7 among HCV patients (16,59), or a nonsignificant difference versus control (65). This difference may be explained by different HCV genotypes, viral coinfections, differences in study methodologies as well as differences of immunogenetics, inclusion criteria of study populations, and protocol of antiviral treatment regimens.
TLR7 gene expression and protein level were fully recovered in treated HCV patients after receiving SOF and DCV therapy with or without RBV. This may reflect the essential role of TLR7 as an antiviral innate immunity tool against HCV, as well as in viral pathogenesis and outcomes of HCV treatment (32). Other treatment regimens such as pegylated IFN α2b with RBV restored upregulation of TLR7/8 gene expressions in peripheral monocytes in previous studies (10,38). In addition, Aldesouki et al. found that SOF plus RBV or SOF plus RBV and pegylated IFN-α upregulated the expression of TLR7 gene in PBMCs (3). Also IFN monotherapy recovered TLR7 gene expression in several studies (12,23). Alternatively, Dowran et al. found no significant differences regarding transcriptomic profiling of TLR7 between responder and nonresponder HCV Iranian patients infected with genotype 1 and who received the standard pegylated IFN-α plus RBV for 48 weeks (19).
Unlike liver biopsy specimens, peripheral blood sample is easy to collect and can be repeated, and PBMCs exhibit higher levels of TLR mRNA compared with other human tissues. Hence, the current study considered TLR7 gene expression in PBMCs as a reliable biomarker for prognosis among HCV-infected patients by ROC curve analysis. A significantly statistical difference was obtained in TLR7 gene expression with a promising value of this innate immune receptor as a predictive biomarker for responsiveness of HCV treatment.
Conclusion
In conclusion, Egyptian treated HCV patients exhibited partially recovered peripheral B cell subpopulations as well as accumulation of CD19+CD21low/− B cells. They may be at risk to develop B cell-associated extrahepatic manifestations, so monitoring B cells at several points of time after completion of treatment should be considered. Moreover, persistent HCV infection downregulated the expression of TLR7, while it has been normalized in treated HCV patients with DAAs, which confirm the evasion of this virus from immunosurveillance by impairing innate immune receptors. TLR7 may be used as a predictive marker for chronic HCV infection and response to HCV treatment between responders and nonresponders and targeting this molecule by its agonists and antagonists may be used as a potential therapeutic during chronic HCV infection or by immune-potentiation of total memory B cell subsets. There was an increased level of IL-7 among treatment-NAI HCV patients without significant difference compared with the patients after successful DAA, and it was also correlated with transitional B cell subsets in untreated patients, but not with the frequencies of other peripheral B cell subpopulations.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The authors received no financial support for the research, authorship, and/or publication of this article.
Supplementary Material
Supplementary Table S1
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.