
Abstract
To this day, the coronavirus disease 2019 (COVID-19) pandemic has not shown signs of abating. Moreover, the virus responsible for the pandemic, severe acute respiratory syndrome coronavirus 2, has evolved into three different variants. This phenomenon highlights an even greater need to develop drugs and vaccines to control the rate of infection and spread of the disease. As of July 7, 2020, at least 160 vaccine candidates, 21 of which have entered the clinical trial phase, have been developed. This article describes the latest advances in development, reliable platforms, strategies used, and challenges that remain in developing COVID-19 vaccines.
Background
In late 2019, a unique case of pneumonia resulting in mortality was identified in Wuhan, China. The cause of this disease was immediately identified as a betacoronavirus that had never been reported earlier; this virus was subsequently named severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) by the World Health Organization (WHO) (29). The virus quickly spread to more than 200 countries, with 12.2 million confirmed cases and 555,000 deaths recorded as of July 11, 2020 (52). Normal living patterns have been greatly affected by the virus as a result of calls for isolation and quarantine. The impact of the coronavirus disease 2019 (COVID-19) pandemic poses great challenges not only to health systems but also to the global economy, especially if its spread is not immediately managed (23) (Fig. 1).

Global distribution of COVID-19 cases. As of July 10, 2020, ∼23,000 new cases per day have been reported. The United States has more than 3.0 million confirmed cases (52). COVID-19, coronavirus disease 2019.
Coronavirus is a positive-sense single-stranded [(+) ssRNA] virus encapsulated by an envelope belonging to the Coronaviridae family. Four classes of coronavirus have been discovered: alpha, beta, gamma, and delta. Betacoronaviruses include Middle East respiratory syndrome coronavirus (MERS-CoV), SARS-CoV, and SARS-CoV-2 (4). Most coronaviruses have 8–10 open reading frames (ORFs). ORF1a is translated into polyprotein 1a (pp1a), and ORF 1b is translated into pp1b; these polyproteins are then processed by viral proteases to yield 16 non-structural proteins containing RNA-dependent RNA polymerase (RdRp) enzymes. The viral RNA is replicated via transcription of the minus strand template by RdRp. In this process, the virus generates six to nine subgenomic messenger RNAs (sgmRNAs), which are then translated into accessory and structural proteins (46). Envelope (E), membrane (M), nucleocapsid (N), and spike (S) proteins are translated from sgmRNA located at the 3′-end of the genome (9).
The S protein mediates the viral invasion of the host cell, starting with binding to the angiotensin converting enzyme 2 (ACE2) receptor through the receptor-binding domain (RBD) in the S1 subunit, followed by fusion with the host membrane through the S2 subunit. After entering the cell, the virus releases ssRNA that is translated into a polyprotein, which is separated into effector proteins by papain-like protease (PLpro) and 3C-like protease (3CLpro) proteinases. PLpro also plays a role in deubiquitinating host cell proteins, including interferon regulatory factor 3 and NF-κB, leading to the suppression of the immune system (6). Similar to MERS and SARS, SARS-CoV-2 attacks the lower respiratory system to cause viral pneumonia, coupled with gastrointestinal, heart, liver, kidney, and central nervous system disorders, leading to multiorgan failure (47). The S protein–ACE2 interaction is, therefore, considered a significant step in the initiation stage of the infection process. Recent Cryogenic Electron Microscopy structure analyses have shown that the binding affinity of the SARS-CoV-2 S protein to ACE2 is 10–20 times greater than that of the SARS-CoV S protein (55). As a result, the transmission rate of SARS-CoV-2 is also much higher than that of SARS-CoV (48).
Methodology
This review includes articles in the PubMed database obtained by using the keywords “Covid vaccine” and “SARS-CoV-2 vaccine.” Unrelated articles, such as those focusing on the diagnosis and treatment of COVID-19 or those that are not related to vaccine development, were excluded. Of the 53 articles obtained in early April 2020, 35 articles were finally included in our review. We also included reports on the current pandemic situation published in the WHO website and studies related to the genomic characterization of SARS-CoV-2 and vaccine-induced immunological responses.
SARS-CoV-2 Genomic Characterization
Lu et al. successfully elucidated the complete genome of SARS-CoV-2 derived from the bronchoalveolar lavage fluid samples of nine patients who had visited a fish market in Wuhan. Genome analysis showed that SARS-CoV-2 was closely related to two bat-derived SARS-like coronaviruses, namely, bat-SL-CoVZXC21 and bat-SL-CoVZC45, which had 88% whole-genome identity compared with 79% with SARS-CoV and 50% with MERS-CoV. In certain gene regions, the sequence identity between SARS-CoV-2 and bat-SL-CoVZC45 and bat-SL-CoVZXC21 exceeds 90%. In the S protein region, however, the sequence identity between SARS-CoV-2 and the bat-derived SARS-like coronaviruses is only ∼75% (29).
Comparison of the predicted coding region of SARS-CoV-2 revealed the similarity of its genomic organization with those of bat-SL-CoVZXC21, bat-SL-CoVZC45, and SARS-CoV. The genome contains 12 predicted regions, and nearly all encoded proteins in SARS-CoV-2, bat-SL-CoVZXC21, and bat-SL-CoVZC45 are of the same length, with only a few minor insertions or deletions. The principal difference between these coronaviruses is that the SARS-CoV-2 S protein is longer than those of the other coronaviruses (29) (Fig. 2).
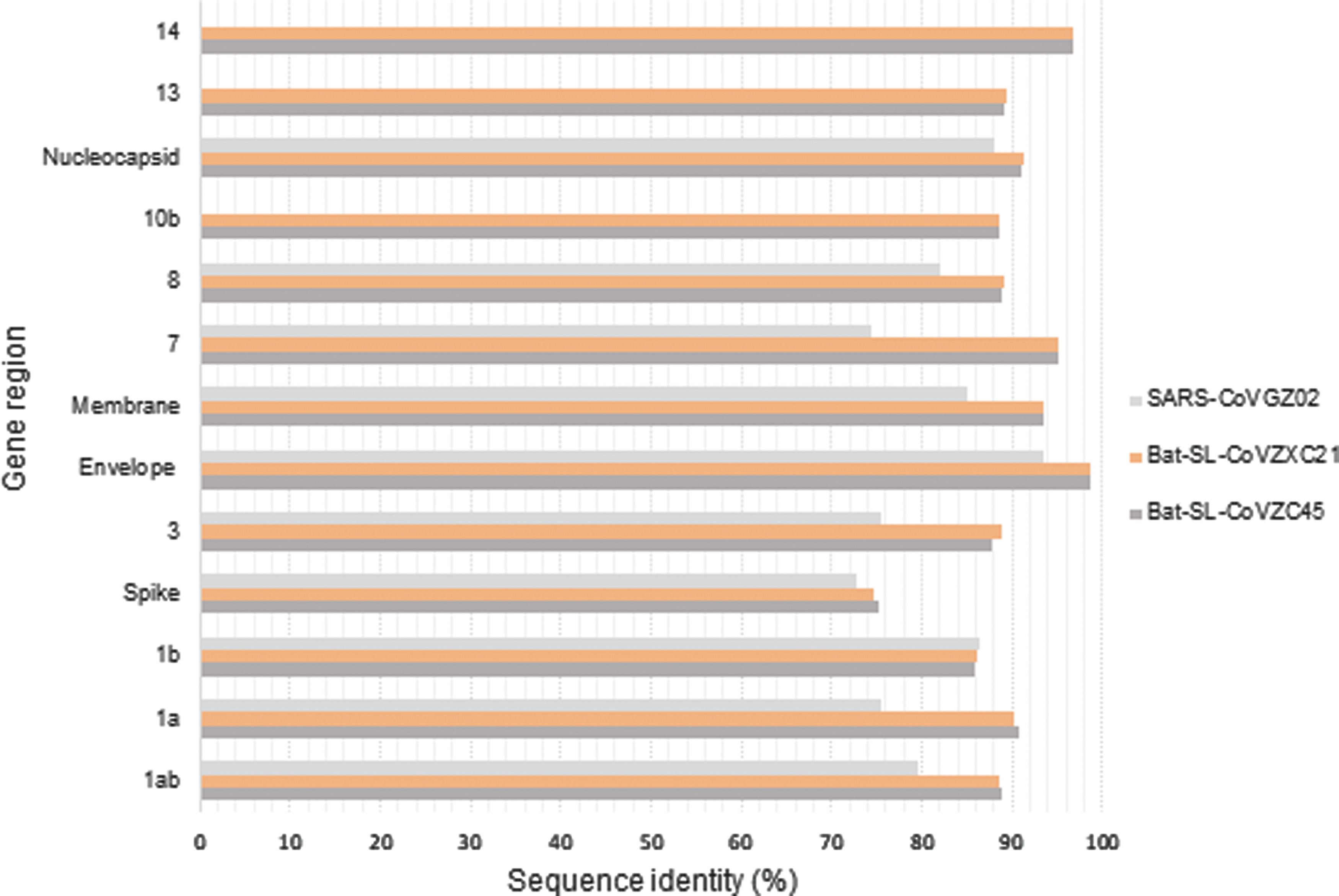
Sequence identity between the SARS-CoV-2 consensus genome and those of other betacoronaviruses (29). SARS-CoV-2, severe acute respiratory syndrome coronavirus 2.
The coronavirus S protein is divided into an S1 domain, which is responsible for binding to the receptor, and the S2 domain, which is responsible for cell membrane fusion (15). The S2 domain of SARS-CoV-2 exhibits 93% sequence identity with those of bat-SL-CoVZC45 and bat-SL-CoVZXC2, but the S1 domains of these viruses exhibit only 68% identity. Approximately 50 amino acids in the S1 domain of SARS-CoV and SARS-CoV-2 are similar; in contrast, the amino acid sequences of the S1 domains of SARS-CoV-2 and the 2 bat-SL-coronaviruses exhibit remarkable differences due to mutations (29).
The RBD of betacoronaviruses is located in the C terminal domain of S1 and binds the virus to its receptor (26). Lu et al. performed a phylogenetic analysis of the RBD of four different lineages for betacoronaviruses and concluded that although SARS-CoV-2 is more similar to the bat-derived coronaviruses at the overall genome level, the RBD of SARS-CoV-2 is more similar in lineage to that of SARS-CoV. Three-dimensional RBD modeling using the Swiss Model program with SARS-CoV RBD as a template showed that the SARS-CoV-2 RBD consists of a core and an external subdomain. The external subdomain of SARS-CoV-2 resembles that of SARS-CoV; thus, SARS-CoV-2, similar to SARS-CoV, is believed to use ACE2 as a binding receptor. However, several important RBD residues involved in the SARS-CoV–ACE2 interaction differ from those of SARS-CoV-2 (29).
Sequence homology studies of the B cell epitopes of SARS-CoV and SARS-CoV-2 revealed that among 10 regions identified, 6 had 90% identity, 2 regions had 80–90% identity, and 2 others had <80% identity. Determination of potential B cell epitopes was also conducted by using bioinformatics approaches as an alternative method with predictive tools from Immune Epitope Database and Analysis Resource (IEDB). The results showed that the surface glycoprotein has the highest number of B cell epitopes, followed by M glycoprotein and N phosphoprotein. Of the five S protein regions that are B cell immunodominant, four regions overlapped in the sequence homology analysis and the bioinformatics approach (13). In the case of T cell epitopes, homological analysis using IEDB showed that the N phosphoprotein and M-derived epitope are the most conserved, with >85% identity. Orf1ab and surface glycoprotein epitopes are moderately conserved, whereas Orf3a is the least conserved (13). Kumar et al. identified six identical epitopes between the S proteins of SARS-CoV and SARS-CoV-2 (22). These antigenicity data suggest similarities between SARS-CoV-2 and SARS-CoV, and this similarity presents a novel strategy for developing SARS-CoV-derived epitope-based vaccines or new SARS-CoV-2 epitope-specific vaccines.
A recent study by Forster et al. found that SARS-CoV-2 has developed into three variants, namely, A, B, and C, which can be distinguished by changes in amino acids. Type A was the first variant to be observed and has a close relationship with bat coronavirus. Two subclusters of type A distinguished by the synonymous mutation T29095C have also been identified. The C allele subcluster shows a relatively long branch of mutations, and 15 of 33 patient samples in this subcluster were found outside of East Asia, particularly in the United States and Australia. Two mutations occurring in the type A variant result in the B variant; these mutations are the synonymous mutation T8782C and nonsynonymous mutation C28144T, which converts leucin to serine. The ancestral B variant was found in all samples originating from East Asia, and all samples outside Asia showed a mutation. Forster et al. thus assumed that the initial type B virus found in Wuhan had adapted immunologically or environmentally to a large population and needed to undergo further mutation to overcome resistance outside East Asia. The type C variant is derived from the type B variant with the nonsynonymous mutation G26144T, which involves the replacement of glycine by valine. This variant was found in European samples and was detected in Hong Kong, Taiwan, and South Korea but not China. Phylogeographic patterns can be influenced by travel history, research findings, and sample size. However, the possibility that variations in mutations can modulate the clinical presentation and spread of a disease are worthy of consideration (11).
Immunological Mechanism of Vaccines
The majority of vaccines are believed to provide protection by neutralizing antibodies (37). However, some evidence shows that T cells play an important role in the protection afforded by vaccines. Several widespread diseases, such as HIV, tuberculosis, and malaria, require strong T cell responses for protection. T cell-based vaccines can induce antigen-specific memory T cells that can persist for long periods and provide protection against subsequent infections. T cell differentiation induced by vaccines can lead to the production of cells with diverse phenotypes and functions. Naive CD4+ cells can be differentiated into several subsets of T helper cells with different functions for each pathogen, whereas naive CD8+ cells can differentiate into effector cells that circulate or settle in particular tissues and provide immediate protection against infection at the entry point of the pathogen. In addition, central memory T cells remain in the T cell-rich area of lymphoid organs and provide a collection of precursor cells that undergo rapid clonal expansion in response to antigens and differentiate into effector cells (38).
The persistence and quality of memory T cell responses can be controlled at several stages, for example, by encouraging the clonal expansion phase, reducing the contraction phase, and stabilizing the memory phase. The rate of clonal expansion depends on several variables, including mobilization of antigen-specific T cells to dendritic cells (DCs). T cells undergo three phases of interaction with antigen-bearing DCs. Different DC subsets can produce different types of immune responses (38). When productive interactions occur between DCs and T cells, clonal expansion can be influenced by several variables, including cytokines such as interleukin (IL)-18 and IL-12, which are secreted by DCs. These cytokines induce T cells to produce interferon-γ, which directly promotes CD8+ expansion (21). IL-6 released by DCs can also reduce the suppressive effect of T regulatory cells (36). Antigen-specific T cells may differentiate into several types of cells, including T helper cells, T regulatory cells, effector cells, and central memory cells (38). Innate control of B cell and T cell responses may be considered a hierarchy in which DC is an innate receptor (39) At the cellular level, the type of DC subset and pattern-recognition receptors (PRRs) triggered by vaccines directs how T cells are differentiated. Therefore, adjuvants targeting specific DC subsets or PRRs can be useful in generating specific responses (38).
Besides regulating T cell responses, the primary role of innate immunity is to regulate the quality, magnitude, and persistence of antibody responses. In some cases of some vaccines, such as those for diphtheria, tetanus, polio, and hepatitis A, protection depends on the extent of the antibody response. For other vaccines, the quality of the antibody is much more important. Of the total antibody bound to an antigen, only a small portion is capable of neutralizing the pathogen. The persistence of the antibody response is also important. Understanding how the persistence of a response can be improved through appropriate adjuvant and vectors is important (38). For example, immunization with a nanoparticle-based vaccine containing antigens plus MPL, a TLR4 ligand, could provide better plasma cell persistence for more than 1.5 years (19).
Antigen-specific antibody responses to T cell-dependent antigens occur via two pathways. DC-mediated stimulation of antigen-specific Th cell is regulated by a subset of DCs and PRRs. Activated antigen-specific B cells move toward the surface between B cell follicles and T cell areas. Interactions then occur between B cell and Th cell and are followed by clonal expansion of B cells. Some B cells migrate to the lymphoid area and differentiate into short-lived plasma cells, whereas others move to B cell follicles, proliferate, and form germinal centers. Some T follicular helper cells express CXCR5 and enter follicles. The expression of the CXCR5 chemokine receptor and downregulation of the CCR7 chemokine receptor allow CD4+ to return to the follicle and help differentiate germinal center B cells. In the initial stages of germinal center development, B cells reduce immunoglobulin (Ig) expression and somatic hyperpermutation occurs in their Ig gene. Then, B cells stop dividing and re-express the mutated Ig gene. Cells with heightened affinity for an antigen are selected to increase the affinity of antibody–antigen binding to DCs and T helper cells. These selected cells then differentiate into long-lived plasma cells or memory B cells (38).
COVID-19 Vaccine Development
Vaccines are considered the most effective and economical strategy to prevent and control infectious diseases (50). The development of an effective vaccine against SARS-CoV-2 infection is urgently needed. Genome mapping of S glycoproteins and epitopes of SARS-CoV-2 can accelerate the development of more specific vaccines. The RNA vaccines may achieve a potential immune response against various infectious diseases and cancer (42,51). Conventional vaccines stimulate antibodies by utilizing proteins purified from pathogens or whole cells (live-attenuated vaccines; LAVs). However, although they are very effective, vaccines can take several years to develop (40).
Challenges in developing COVID-19 vaccines
The Coalition for Epidemic Preparedness Innovations has begun to develop a number of COVID-19 vaccine platform technologies. An ideal platform would accelerate development efforts from viral sequencing to clinical testing in less than 16 weeks, show a consistent immune response among pathogens, and be suitable for large-scale production by using a pathogen-agnostic platform. Unfortunately, even with a new platform technology, vaccine development is still hampered by several challenges. Although the S protein of the virus is known to be a promising immunogen, the optimization of the antigen design is still debated. Previous preclinical testing of SARS and MERS vaccine candidates showed lung disease susceptibility. Therefore, testing with appropriate animal models and thorough safety monitoring in clinical trials are very critical (31).
Vaccine development is a long and expensive process, requires the development of a number of candidates, and takes many years to produce an approved product (12). Because of the high cost and risk of failure, vaccine developers generally follow linear stages, pausing several times for manufacturing process control and data analysis. Rapid vaccine development requires a new pandemic paradigm with a fast start and several steps executed in parallel before confirmation of success is obtained at other stages; these complex processes increase the financial risk of vaccine development. In instances when a vaccine development platform requires clinical evaluation, phase 1 clinical testing can be conducted in parallel with testing on animal models (31).
The challenge for regulatory agencies to deliver a SARS-CoV-2 vaccine in a timely manner is identical to that for researchers. Evaluation of the safety of vaccine candidates is expected to be of great interest to the global community. Immunopathogenesis plays an important role in COVID-19. Therefore, ensuring that the vaccine does not cause the same adverse immune response as the native virus is crucial; the related work can help determine the immunogens to be designed. The development process will face other challenges as well, for example, whether any qualified manufacturing process that can realize the development of vaccine candidates exists, whether production requirements and vaccine regulations in one country will be accepted in other countries, and whether political and commercial considerations will become obstacles to solving global SARS-CoV-2 vaccine problems (30).
SARS-CoV-2 vaccines under development
As of July 7, 2020, at least 160 vaccine candidates from pharmaceutical industries and academic institutions across the world were reportedly being developed. Eight technological platforms and several types of vaccines have been proposed (Table 1).
Coronavirus Disease 2019 Vaccine Candidates
VLP, virus-like particle.
Source: World Health Organization (53).
Replicating and non-replicating viral vectors
Viral vectors provide a means to provide vaccine antigens to cells or tissues. These vectors may generally be classified as replicating or non-replicating vectors. Adenoviruses (Ads) and poxviruses are examples of vectors with both traits, whereas non-replicating vectors include adeno-associated viruses, alphaviruses, and herpesviruses. Replicating vectors include the measles virus (MV) and vesicular stomatitis virus (VSV) (41). Each vector has unique features, including insert capacity, advantages, and disadvantages.
Ads are one of the most highly exploited vectors for vaccine development. According to WHO data for COVID-19 vaccine development, several institutions are relying on non-replicating Ads for their programs. The insert capacity of an Ad-based non-replicating vector is 7–8 kb, and this vector is relatively safe, physically and genetically stable, and can infect DCs and dividing or non-dividing cells. Moreover, this type of vector is not integrated and has been used in gene therapy. However, Ad vectors require high doses to enhance immunity (7).
Modified vaccinia virus Ankara (MVA) is a type of non-replicating poxvirus vector obtained from the loss of ∼15% of the genome due to serial passaging in embryo fibroblasts. The advantages of MVA over avian poxvirus include excellent immunogenicity; however, prior immunity must be considered (35).
The use of an MV-based vector, an ssRNA virus, is generally new for HIV vaccines and several other viruses, such as HBV, mumps, and SARS (27). Attenuated MV is able to provide cellular and humoral immunity over long periods, reaching even up to 25 years. Its ability to infect DCs and macrophages is believed to be responsible for this ability (60). MV vectors are also genetically stable and do not cause integration. However, similar to MVA, the presence of prior immunity is an obstacle in the use of a measles vector (41).
VSV has also been used for SARS-CoV-2 vaccine candidate development programs. The advantages of VSV over other vectors include replication in the cytoplasm, which avoids integration with host cell DNA, high expression of the transgene due to cessation of the translation of host cell mRNA, easy production processes, low level of prior immunity, and ability to be administered through the mucosa. However, VSV is neurovirulent to rodents and nonhuman primates. Thus, aggressive attenuation must be carried out to meet safety criteria, and ways to enhance immunogenicity of the attenuated vector must also be explored (18).
mRNA vaccines
The development of mRNA synthesis, modification, and delivery technologies has helped mRNA vaccine research generate remarkable interest over the past two decades. Indeed, mRNA vaccines are not only limited to infectious diseases but can also be used for cancer (34). mRNA-based vaccines contain the antigen genome, which can then be translated in the host cell after vaccination (58). The mRNA vaccine provides a promising alternative to conventional vaccine approaches because of its high potential, short production cycle, low production cost, and safe delivery (34). The development of an mRNA vaccine includes antigen selection, sequence optimization, screening of modified nucleotides, optimization of the delivery system, and evaluation of immune responses and safety testing (17).
The mRNA-based vaccine for COVID-19 developed by Moderna, Inc., in collaboration with the National Institute of Allergy and Infectious Diseases, has entered phase 2 clinical trials. In contrast to conventional vaccines produced in cell culture systems, Moderna, Inc., vaccines are designed in silico so that the development and evaluation process may be performed faster (32).
DNA vaccines
DNA vaccines provide protection through direct injection of plasmids encoding antigens to generate a broad immune response. This advantage has been applied to both prophylactic and therapeutic vaccines (56). DNA vaccines are also considered superior to mRNA vaccines in terms of formulations needed to maintain stability and delivery efficiency. Nevertheless, DNA vaccines must enter the cell nucleus, where they are subject to the risk of integration and mutation in the host genome (28). Various DNA vaccine platforms have evolved to increase the efficacy of the resulting vaccines; for example, electroporation has been employed for plasmid delivery, and the addition of adjuvants can enhance the immune response (43).
The INO-4800 DNA vaccine developed by INOVIO Pharmaceuticals, Inc., has entered the clinical trial phase (53). INO4800 is able to induce T cell activation through the delivery of DNA plasmids encoding the SARS-CoV-2 S protein. The primary advantage of this vaccine is its ability to induce antibodies and immune cells through intradermal delivery (16).
Inactivated and LAVs
Inactivated (whole cell-killed) and LAVs similarly include several antigenic components and, thus, are able to induce a broad immunological response (45). Both platforms are classified as conventional vaccines with mature preparation technologies (59). Inactivated vaccines are produced from microorganisms that are killed either physically or chemically. One the one hand, these vaccines are not always able to induce an immune response and, even if a response occurs, it may not last long. Therefore, several vaccine doses may be needed to generate an adequate immune response. On the other hand, this vaccine platform has no risk of inducing disease and is considered more stable compared with LAVs (54).
First developed in the 1950s, LAVs are mainly derived from disease-causing pathogens, including bacteria and viruses that have been attenuated. The pathogens in LAVs can grow in vaccinated individuals but will either not cause disease or produce only mild symptoms on account of their weak condition. LAVs are believed to be able to stimulate good immune responses and are comparable with the original form (wild-type) of the pathogen. Living microorganisms can induce an immune response until the stage in which memory cells are formed. However, the use of LAVs is limited because of their poor security and stability. Attenuated pathogens may still return to their original form and cause disease, as in the case of vaccine-associated paralytic poliomyelitis. Individuals with weakened immune systems, such as those with HIV, may be unable to receive LAVs. Moreover, LAVs must be cultured and are therefore prone to contamination by other viruses present in the culture tissue. In addition, there is a possibility for errors to occur during the immunization process. Some LAVs are obtained in lyophilized form and must be reconstituted with specific solvents (54).
Virus-like particle vaccines
Virus-like particles (VLPs) are a supramolecular structure of multiproteins and carry various characteristics of a virus (24). VLPs mimic the conformation of native viruses without having infectious properties, because they do not carry genetic material (1) VLPs can be produced in more than 170 hosts, including bacteria, insects, fungi, mammalian cells, and, interestingly, plant cells (20). When inside the host immune system, VLPs induce an immune response without triggering the emergence of side effects, which occurs with native viruses. Some VLP-based vaccines such as HBV and HPV vaccines are currently available. An important advantage of this platform is the ability of VLP vaccines to carry antigens in a dense and repetitive manner and effectively generate crosslinking of B cell receptors (BCRs) (57). BCR crosslinking by multivalent antigens causes conformational changes in BCR and induces BCR microcluster formation, which leads to B cell activation and signaling (44).
VLP-based vaccines are basically designed to target B cells and induce potent antibody responses after the activation of T helper cells and presentation of MHC class 2 molecules by antigen-presenting cells. The presentation of antigens by VLPs is not only limited to MHC class 2 molecules but also includes MHC class 1 molecules for initiating CD8+ T cell responses. The size and structure of VLP-based vaccines facilitates cross-presentation of their peptides to MHC class 1 molecules (14). This phenomenon is an added value of VLP-based vaccines and is advantageous when designing VLP-based vaccines for cancer and other chronic diseases (3).
Protein-subunit vaccines
Subunit vaccines include one or more antigens with strong immunogenicity that can stimulate the host immune system efficiently. In general, this vaccine platform is relatively safe and easy to produce but requires the addition of adjuvants to induce a strong immune response (59). Studies on subunit-based vaccines show an increase in the T cell immune response and significantly higher titers of neutralizing antibodies in vivo (33). Similar to inactivated vaccines, subunit vaccines do not contain living components and are considered safe. However, whether memory cells can be formed for future protection is unknown. Subunit vaccines are generally categorized into three classes: protein subunit-based vaccines, polysaccharide vaccines, and conjugate subunit vaccines (54).
Protein subunit-based vaccines present antigens to the immune system without using virus particles; instead, these vaccines utilize a specific pathogenic protein isolate. The main drawback of this platform is that denaturation of the protein isolates may produce bonds with other nontarget antibodies (54).
Several institutions with a COVID-19 vaccine development program utilize this platform, especially on using the S protein as an antigen. Clover Biopharmaceuticals, Inc. is currently conducting phase 1 clinical trials for a SARS-CoV-2 trimeric S protein vaccine (8). SARS-CoV-2 S glycoprotein has three S1 heads and trimeric S2 stems (25). The development of these native-like trimeric viral S proteins is conducted on mammalian cell expression systems. Clover Biopharmaceuticals together with GSK and Dynavax is developing an adjuvant system for S-Trimer to maximize the immune response (8). Development of a protein subunit-based vaccine with transformative molecular clamp technology is being carried out by the University of Queensland. Molecular clamp technology enables the maintenance of the shape and conformation of surface proteins, which are often disrupted when using recombinant technology. Thus, a stronger immune response is obtained (49).
Peptides or synthetic epitope-based vaccines have also been investigated. This type of vaccine contains only a few fragments of antigens and is usually prepared through chemical synthesis techniques; as such, they are relatively easy to prepare and control. However, the low molecular weight and structural complexity of the antigen renders its immunogenicity relatively weak. Therefore, modifications in their structures, delivery systems, and adjuvant selections are needed (5). Generex Biotechnology is developing a peptide vaccine using NuGenerex Immuno-Oncology Ii-Key technology. Researchers from the Hong Kong University of Science and Technology are currently in the stages of screening for B and T cell epitopes from S and N proteins, both of which are conserved in SARS-CoV-2 (2).
Conclusion and Future Prospects
The impact of the COVID-19 pandemic has far exceeded the initial estimations. The virus has spread across the world, and the inflection point has yet to be reached, indicating that transmission will continue to occur unless the virus is firmly controlled. Thus, developing a COVID-19 vaccine that can be used globally is a major priority. Vaccine development is a long and expensive process. New approaches to accelerate the preparation of a suitable COVID-19 vaccine have been introduced, including platform development, preclinical testing in parallel with phase 1 clinical trials, and easy licensing. Increasing our understanding of the characteristics of SARS-CoV-2 can help guide the development of more specific vaccines. To date, more than 160 vaccine candidates, 21 of which have entered phase 1, 2, or even 3 clinical trials, have been reported. An important consideration before the approval of a COVID-19 vaccine is its proven safety and efficacy. Careful and thorough planning must be conducted to ensure that all individuals have the same access to vaccines. Vaccine administration for medical personnel and high-risk individuals should be prioritized. Problems related to vaccine ownership, large-scale production financing, and supply chains must be resolved. High-income countries must be prohibited from monopolizing global supplies, similar to the case during the influenza A/H1N1 pandemic (10).
Footnotes
Author Disclosure Statement
The authors report no conflicts of interest in this work.
Funding Information
This study received no funding.