
Abstract
We aimed to verify the influence of intrinsic and extrinsic cell apoptotic pathways on the inhibition of cellular apoptosis in patients with tropical spastic paralysis/myelopathy related to human T cell lymphotropic virus type 1. The databases accessed were PubMed, Scopus, Science Direct, and Web of Science. Neither the time of publishing nor the language of the articles was limited. The descriptors used for this systematic literature review were: Tropical Paraparesis, Proto-Oncogenic Protein C, Bcl-2, Bcl-X Protein, Bax protein, Fas ligand (FasL) protein, Fas receptor, TNF-related apoptosis-inducing ligand and Fas-associated protein with death domain (FADD)-like apoptosis regulating. The search resulted in 546 articles from which 9 articles were selected for analysis; ranging from serum levels of Bcl-2, Fas and tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) measured by enzyme-linked immunosorbent assay and the levels of cellular expression of Bcl-2 and Bcl-xL the TCD4+ lymphocytes accessed by western blot. Most studies accessed either gene expression or polymorphism of Fas, FasL, and TRAIL in patients with HTLV-1-associated myelopathy/tropical spastic paraparesis (HAM/TSP), whereas one study used flow cytometry and fluorescence to determine Fas expression. Increased Bcl-xL expression inhibited T lymphocyte apoptosis, whereas Bcl-2, serum levels, and cellular expression did not influence T lymphocyte apoptosis and serum levels of Fas were significantly higher and associated with markers of leukocyte activation in patients with HAM/TSP. In addition, Fas polymorphism (FAS-670AA) was associated with higher proviral load. There is a need for additional research on this issue since the number of patients was small and the studies presented higher heterogeneity.
Introduction
At present, there are ∼20 million people infected with the human T cell lymphotropic virus type 1 (HTLV-1) all over the world. Brazil concentrates 2.5 million of these carriers (11,12,26,42,53). Although ∼95% of the infected individuals remain asymptomatic, 2–4% HTLV-1 develop HTLV-1-associated myelopathy/tropical spastic paraparesis (HAM/TSP) (13,27,65) and 2–3% develop T cell leukemia or lymphoma (43). Other rarer manifestations may also be noticed, such as polyneuropathy, cognitive changes (29), arthropathies, polymyositis, uveitis, infectious dermatitis, and Sjögren's syndrome (51,55,69).
It is hypothesized that infected CD4+ T lymphocytes of the individuals with HAM/TSP could cross the blood–brain barrier (BBB) causing chronic inflammation in the central nervous system (CNS) and demyelination (89). In the spinal cord, the most affected areas are the thoracic and the lumbar regions (60,62), whereas in the brain, the most affected area is the white matter (6,48). However, the specific mechanism related to the development of this neurological disease has not yet been elucidated, and up until now, the increase in viral load, polymorphisms of the human leukocyte antigen complex, and being infected with the transcontinental HTLV-1 subtype are considered risk factors for the development of HAM/TSP (54).
It is known that cell–cell contact is essential to spread HTLV-1 infection into the host,11 and apoptosis is a physiological and crucial mechanism for the development and homeostasis of many tissues, including the immune system (61). Thus, apoptosis can play an important role in the clonal deletion of autoreactive T cells in autoimmune diseases (34).
Cellular apoptosis can be controlled by two distinct pathways: one extrinsic and the other intrinsic. The extrinsic pathway is mediated by specific receptors present on the cell surface or cell death receptors, such as Fas, which, through connection with its ligand (FasL), Fas-associated protein with death domain (FADD), and tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) activates pro-caspase proteins, causing cell death. The intrinsic pathway is controlled by mitochondria and occurs because of intracellular stress. It also causes cell death through the activation of specific proteins that inhibit the anti-apoptotic action of Bcl-2 (bad, bak, bax) (4,10,19,39).
Bcl-2 is a mitochondrial membrane protein with the unique ability to suppress apoptosis by physiological or pathological stimulus (80,88). Bcl-xL is another mitochondrial protein of the Bcl family 2, which has the ability to inhibit apoptosis through cytochrome c regulation and increase leukocyte survival (31).
Fas, FasL, and TRAIL are death receptors. The link between trimers of Fas and FasL results in the recruitment of adapter proteins called FADD (death domain associated with FAS), triggering cell death through the activation of caspase-8 autocleavage. Cells infected with HTLV and normal cells will induce apoptosis (39,47).
Considering that there are still no specific consolidated interventions for patients with HTLV, it is important to understand the pathophysiological mechanism of HTLV, which will enhance future drug interventions by improving the control of this infection, the clinical aspects, and the quality of life of these patients. Therefore, the aim of this review was to verify the role of Bcl-2, Bcl-xL, Fas, FasL, FADD, and TRAIL proteins in the cell apoptosis in patients with HAM/TSP.
Materials and Methods
To conduct this systematic review the research was performed in the following databases: PubMed, Scopus, Science Direct, and Web of Science. Neither the time of publishing nor the language of the articles was limited.
The descriptors used in the research were “Tropical Spastic Paraparesis,” “Proto-Oncogone Proteins c-bcl-2,” “bcl-X Protein,” “Bax protein,” “Fas ligand protein,” “Fas receptor,” “TNF related apoptosis inducing ligand,” and “FADD-like apoptosis regulating” using the following search combinations: Tropical Spastic Paraparesis AND Proto-Oncogone Proteins c-bcl-2; Tropical Spastic Paraparesis AND bcl-X Protein; Tropical Spastic Paraparesis AND Bax protein; Tropical Spastic Paraparesis AND FasL protein; Tropical Spastic Paraparesis AND Fas receptor; Tropical Spastic Paraparesis AND TNF related apoptosis inducing ligand; Tropical Spastic Paraparesis AND FADD-like apoptosis regulating. The search strategy was used according to the specificity of each database.
The inclusion criteria were as follows: studies whose sample included patients with tropical spastic paraparesis from HTLV-1, adult individuals (over 18 years old) of both sexes, and studies that analyzed serum or cell expression levels of Bcl-2, Bcl-xL, Fas, FasL, BAX, TRAIL, or FADD.
The exclusion criteria were as follows: studies with individuals coinfected with the human T cell lymphotropic virus type 2 (HTLV-2); coinfected with HIV and hepatitis C; causes of spastic paraparesis other than HTLV-1; articles in which studies were developed with animals or T cell leukemia/lymphoma (ATL); review studies; case reports; studies in which basic demographic information of the sample did not include sex and age; studies with no description about techniques used to measure serum and cellular levels of the protein.
The research and data extraction were carried out by two evaluators. The selection of articles was initiated by reading their titles and abstracts. Then, a meeting was held between the evaluators to align the choice of articles that would pass to the stage of complete reading of each article. In case of disagreement, a third evaluator was called. Finally, the articles that met the eligibility criteria were read in their full versions.
The calibration between the evaluators was carried out through the Simple Agreement Index (ICS), performed after the selection of articles in the databases, reaching for 100% agreement. The protocol for this systematic review was registered with PROSPERO, under the CRD 42018098917 number. This is a review study, so the study does not include samples of humans or animals, and does not apply to Instutional Review Board (IRB) approval.
The variables analyzed were information about the research team (authors and year of publication), characteristics of the population (sample, sex, and groups), groups, cases and controls (number of patients, mean age, and diagnostic criteria), the way in which data were obtained (type of biological material and sample selection), and measuring methods. Table 1 provides the general data of the studies included in this review.
General Data from the Included Articles in the Systematic Review
ATL, adult T cell leukemia/lymphoma; CSF, cerebrospinal fluid; ELISA, enzyme-linked immunosorbent assay; FasL, Fas ligand; HAM/TSP, HTLV-1-associated myelopathy/tropical spastic paraparesis; HTLV-1, human T cell lymphotropic virus type 1; OIND, other inflammatory neurologic diseases; RT-PCR, real-time-polymerase chain reaction; TRAIL, tumor necrosis factor-related apoptosis-inducing ligand; UFPA, Federal University of Para (Brazil).
The main results regarding differences in expression levels of proteins (Bcl-2, Bcl-xL, Fas, FasL, and TRAIL) or polymorphisms frequencies between groups are given in Table 2.
Main Results on Bcl-2, Bcl-xL, Fas, FasL and TRAIL Observed in the Articles Included in the Systematic Review
HLA-DR, cell surface marker of human leukocyte antigen.
The quality analysis of the articles was made with a modified version of the Strengthening the Reporting of Observational studies in Epidemiology Checklist (STROBE) (85), adjusted specifically for the evaluation of case–control studies.
Results
Overall, 546 articles were identified in the databases: PubMed (n = 490), Scopus (n = 42), Science Direct (n = 7), and Web of science (n = 17). Then, a total of 241 duplicates were removed. The remaining 305 articles had their titles and abstracts analyzed in relation to the inclusion and exclusion criteria. After analyzing the titles and abstracts, the 26 remaining articles were thoroughly read.
After the full reading, 17 articles were excluded, owing to the following motives: n = 1 it was an editorial piece; n = 3 experimental studies using animals; n = 4 there was not patients with HAM/TSP; n = 3 the patients studied had ATL, and n = 6 investigated other pathology or different type of study. After this last phase, nine articles were finally included in the systematic review. They were published between 1997 and 2017 and carried out by different research groups located in Japan, Peru, Iran, and Brazil. One study originated from an international effort from researchers from Belgium, United States, Brazil, Peru, France, and Portugal. The flowchart shows this process in detail, following the model of the Preferred Reporting Items for Systematic Reviews and Meta-Analyzes (PRISMA) (51) (Fig. 1).
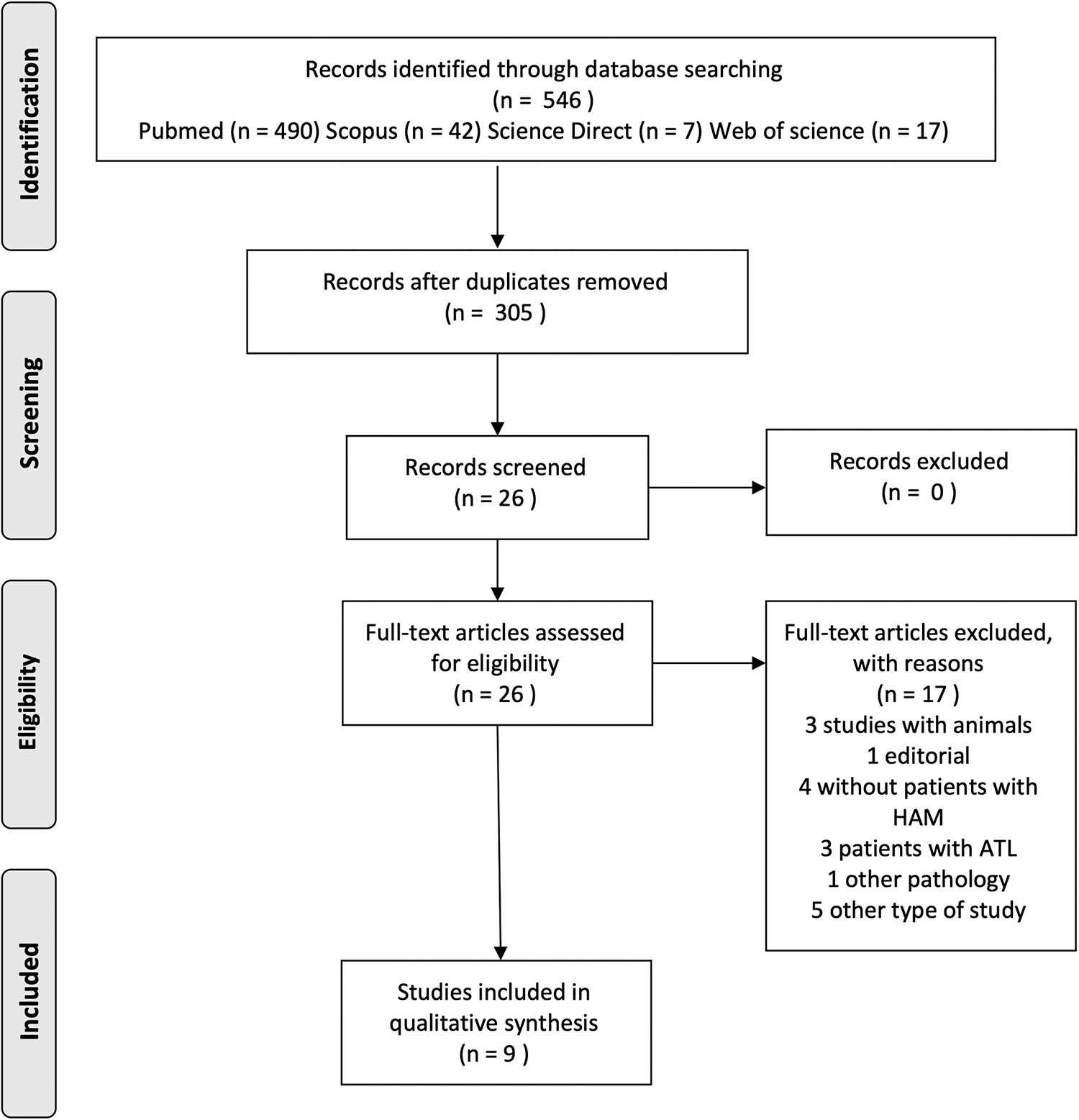
Framework of research and article selection for the systematic review according to preferred reporting items for systematic reviews and meta-analyses.
Table 1 provides the characteristics of the studies included in this review. The studies sample ranged from 12 to 216 patients. Six studies used the Osame criteria to diagnose cases of HAM/TSP. Five articles did not inform the location for selection of the patients and one did not inform the method used for sample selection. In eight of nine articles, peripheral blood samples were used for analysis, two studies used cerebrospinal fluid (CSF) samples, and only one used spinal cord tissues.
The serum levels of Bcl-2, Fas, and TRAIL was accessed by enzyme-linked immunosorbent assay and for cellular expression of Bcl-2 and Bcl-xL in the TCD4+ lymphocytes the western blot approach was used. Expression and polymorphisms of Fas, FasL, or TRAIL were evaluated by real-time-polymerase chain reaction. Only one study used flow cytometry and fluorescence to quantify the levels and expression of Fas (Table 1).
Results presented in Table 2 show Bcl-xL expression significantly higher in patients with HAM/TSP compared with controls (33); however, this was not observed in relation to Bcl-2 levels (33,68). Serum levels of Fas were significantly higher in patients with HAM/TSP compared with controls (49), asymptomatic (49), and other inflammatory neurologic diseases (35). In the CSF of patients with HAM/TSP, the increase in Fas levels was associated with the presence of HTLV-1 antibodies (67). T lymphocytes presented in the active inflammatory lesions in the spinal cord (82) and peripheral blood (49) of patients with HAM/TSP showed higher expression of Fas. The increase in Fas levels was associated with activation of leukocyte markers (49). The FAS-670AA genotype was more frequent in patients with HAM/TSP (84) and was associated with an increase in proviral load (63,84).
Discussion
Several factors are related to the pathogenesis of HAM/TSP and among the several mechanisms involved, the following stand out: virological components (proviral load, bZIP factor mRNA, and expression of basic leucine zipper factor [HBZ]) (8,25), immunological and inflammatory factors represented by cytotoxic T lymphocytes (CTLs) and natural killer cells (54). In patients with HTLV-1, apoptosis can be triggered by intrinsic (Bcl-2 bad, bak, and bax) and extrinsic (Fas, FasL, FADD, and TRAIL) mechanisms (39).
From the phylogenetic point of view, proteins of the Bcl-2 family and their orthologous have been identified in several metazoans (92). Wang suggests that genes of the Bcl-2 family evolved from the multicellular life forms. Humans and rats have orthologs that produce proteins of the Bcl-2 family, which indicates that this family arrangement is stable among mammals (5).
The studies included in this review demonstrated that there was no increase in serum levels or in Bcl-2 expression compared with control groups. One of the studies found an increase in Bcl-xL expression. A Bcl-xL is an anti-apoptotic member of the Bcl-2 family, whose function may change during viral infections and neoplasias (3,79). The proteins expressed by HTLV-1 can modulate levels of proteins of the Bcl-2 family, using specific mechanisms and the inhibition of Bcl-xL expression. However, the Bcl-2 expression seems not to be affected. The Bcl-xL may impact the survival of the T cells, turning this protein crucial on the pathology condition (3,41).
The results related to Bcl-2 should be interpreted with reservations, because these studies did not consider age to correlate cases and controls (33,68). Both groups have a reduced amount of samples (33) and the control group had individuals with several inflammatory neurological pathologies in one of the studies (68). The effects may have an influence on the results especially if they represent an association of small magnitude.
The regulatory role of Bcl-2-mediated apoptosis was demonstrated for Epstein–Barr virus (EBV). The transformation process of epithelial cells related to EBV is probably regulated by the coexpression of LMP-1 proteins (latent membrane oncoprotein) and Bcl-2 observed in the tumor epithelium infected with EBV and nasopharyngeal carcinoma (3). The LMP-1 molecule can overregulate the expression of the anti-apoptotic proteins Bcl-2, MCL-1, and bfl-1, and the process involves the induction of adhesion to the cell surface and the TNFR/CD40 pathway. Another transmembrane protein, LMP2A, appears to have an effect on the transformation of epithelial cells, but not on B lymphocytes. The LMP2A protein inhibits apoptosis mediating the expression of Bcl-2 and Bcl-xL (23).
A 40 kDa HTLV-1 protein called Tax, function of which is to accelerate gene expression (14,20,22,71,73), has been mentioned in various oncogenic mechanisms in T cells by HTLV-1. Some of them concern the transcriptional activating function of this protein, not only of HTLV-1 itself, but also of several cellular genes in trans. This transactivation is mediated through the interaction with the transcriptional factors CRE binding protein, nuclear factor kappa B (NF-kB), and serum response factor (21,74,75,93), causing an increase in the production of these proteins.
They include genes for interleukin-2 (IL-2), IL-6, granulocyte–macrophage colony-stimulating factor, the IL-2 receptor chain, Fra-1, c-Myc, c-Fos, and c-Jun (64,91). The increased expression of these proteins could be related to unregulated cell proliferation in individuals infected with HTLV-1, which is associated with a lymphocyte resistance to apoptosis, and could increase proviral load, the main marker related to the worsening of the disease (7,81).
In patients with HTLV-1, the increase in Bcl-xL expression is mediated through TAX, which promotes the activation of anti-apoptotic genes, inhibiting pro-apoptotic genes interfering on the NF-kB pathway. Increased expression of Bcl-xL inhibits apoptotic signals and favors the survival of T cells infected with HTLV-1 allowing leukemogenesis (3). The regulation of Bcl-xL levels can also be modulated by HTLV-1 proteins (p12), which activate IL-2 receptors and stimulate Janus kinases (JAK 1 and JAK 3), inducing the expression of genes that will give rise to Bcl-xL and the inhibition of pro-apoptotic proteins (BAK1) through the activation of transcription factors (STAT5) (3).
Studies indicate that the persistence of inflammation is related to increased expression of Bcl-xL by T lymphocytes from patients with more severe forms of autoimmune encephalomyelitis (36). In this context, two distinct pathways that lead to T cell apoptosis have been described: passive cell death and cell death induced by activation (AICD). Passive cell death is mediated by the removal of activated T cell growth factor (32,56) and can be performed experimentally by blocking IL-2 production pathways (as by costimulatory signal block) (70). In contrast, AICD occurs when T cells are repeatedly exposed to the antigen and is mediated by the Fas/FasL interaction (86,87) and it depends on previous exposure to IL-2.
Tsukahara et al. (81) showed that constitutive expression of Bcl-xL was associated with resistance to apoptosis; however, no association was found with Bcl-2, Bcl-xL, Bak, or Bax proteins after the decrease of IL-2 in CTLL-2 cells, in which TAX was expressed in response to decreased levels of IL-2. This may be one of the molecular mechanisms of resistance to apoptosis, where TAX renders the CTLL-2 growth independent of IL-2 (37).
Pro-apoptotic action mediated by Bad is inhibited by protein kinase B(AKT) preventing the death of infected T cells (38,79), but their activation depends on other cytokines. Type I and III interferons (IFN-α and IFN-β, and IFN-λ, respectively) are potent antiviral cytokines with immunomodulatory potential in viral infections (16). The high expression of AKT1 in the HTLV-1-infected cell can induce high expression of IFN-λ3 mRNA in patients with HAM/TSP (79). IFN-induced leukocyte has also been associated with more severe cases of HAM/TSP (66,77).
The Fas/APO1 molecule (also known as CD95) is an important surface receptor that signals apoptosis in several cells (35). Fas/FasL interactions regulate a major pathway in apoptosis, which may play an important role in mediating both the antiviral effects and the inflammatory process in neurological diseases, regulation of the immune response and tissue homeostasis, and immunological clearance of virus or tumor cells (82).
Genetic changes and increased soluble levels of this molecule have been associated with the development of autoimmune diseases in mice (2,15), suggesting that it may also play an important role in immune responses to viral infections, such as HTLV-1 (17). During the immune response, the destruction of activated T cells can be induced by signaling through Fas antigen (52,90) and might be the important factor of the mechanisms of demyelination (35). The studies included in this review demonstrated that there was no increase in serum Fas levels in patients with HAM/TSP compared with controls (49), asymptomatic (49), and other inflammatory neurologic diseases (35).
The pathophysiology of HAM/TSP involves T cell migration through the BBB and chronic and long-lasting CNS inflammation (7). Patients with this condition also have an imbalance related to inflammatory markers (89). Persistent activation of T cells of patients with HAM/TSP also modifies the expression of metalloproteinases (MMPs) (40) by astrocytes (28).
The expression of MMPs is regulated by cytokines and its increase promotes the opening of the BBB, cleavage of the constituents of myelin, alteration of the cell–cell connection, and conversion of TNF from inactive to active form (28), aggravating the functional impairment presented by these patients. In one study included in this review, the increase in Fas levels was associated with activation of leukocyte markers (49). However, as it presents several stages, the total number of participants of the study was divided among the stages of the research.
Besides that, in the CSF of patients with HAM/TSP, the increase in Fas levels was associated with the presence of HTLV-1 antibodies (67), and T lymphocytes were the cells that expressed more Fas in the active inflammatory lesions in the spinal cord (82) and peripheral blood (49) of these patients. Sensitivity to induction of apoptosis by anti-FasrAPO-1 was acquired during long-term culture of activated T cells in the presence of IL-2 (17). MMP-2 and MMP-9 were higher in patients with HAM/TSP than those with OIND (40,83) and FasL is easily converted by MMP-like enzymes (78). It is also possible that activation of FasL cleavage from the membrane contributes to the increased concentration of sFasL (67).
The FAS-670AA genotype was more frequent in patients with HAM/TSP (84) and was associated with an increase in proviral load (63,84). The FAS-670 A allele polymorphism promotes an increase in Fas expression and its presence was identified as a risk factor for autoimmune diseases and associated with increased inflammation in patients with hepatitis C (1). Fas promotes migration and invasion in apoptosis-resistant malignant cells (9) and has been postulated to enhance inflammation when activated in nonlymphoid cells (46).
However, no adjustment for population structure was performed in the Brazilian study (patients were selected at the outpatient of a university clinic), considering the diverse ethnic origin of Brazilian population. Despite the population of the other study being larger and having been selected from a cohort study (63), all patients were Peruvian. Thus, both studies have limitations because the genetic characteristics restrict the staggering of the results for individuals of different origins.
The basic leucine zipper factor (HBZ—basic leucine zipper factor) is another regulatory protein that plays an important role in the development of adult T-cell leukemia/lymphoma (ATLL) (44). Rabbit studies have found that at the beginning of the chronic stage of HTLV-1 infection there is a loss of TAX and Gag production, whereas HBZ expression is maintained in a steady state. These results confirm an inverse correlation in the transcriptional sense between TAX/Gag and HBZ expression, suggesting that HTLV-1 can use HBZ for the establishment of chronic infection (45).
Gaudray et al. (24) found that the negative strand of HTLV-1 contains a reading frame that encodes a transcription factor called HBZ (factor HTLV-1 bZIP). The functional profile of the HBZ protein is multifaceted, complex, and appears to function not only as a protein, but also as a regulatory RNA, which promotes the proliferation of host cells. As a protein, HBZ activates the transforming growth factor beta/via Smad, which leads to transcriptional activation of the Foxp3 gene (76).
In patients with HAM/TSP, the HBZ protein is located exclusively in the lymphocyte cytoplasm, unlike the predominant nuclear location of HBZ in leukemic cells of patients with ATLL, suggesting the functional need for HBZ in the development of this neurological disease (46). Considering that patients with HTLV-1 present an imbalance between production of pro and anti-inflammatory cytokines (89), we suggest future studies to investigate the influence of these inflammatory mediators on the regulation of apoptosis in patients with HAM/TSP in comparison with HTLV patients without myelopathy.
After evaluating the articles with the STROBE checklist, the main limitations of the study were the lack of display about selection and classification of patients on the results, lack of sample calculation, and control of confounding factors, in addition to not discussing the bias and limitations of the respective studies. Most studies used the same criteria to classify HAM/TSP patients (Osame scale) and it is noteworthy that this tool is used as classification criteria exclusively for the motor function of the lower limbs (72).
Given these conditions, it is possible that there is a classification bias in relation to the diagnosis of myelopathy, which may have generated results that are not entirely reliable. Thus, it is believed that the identification of biases and limitations can guide future studies, increasing their power of external validity.
Observing the effect of Bcl-2 and Bcl-xL proteins on the inhibition of cell apoptosis and the increase of inflammation promoted by elevation of Fas levels (serum and CSF) in patients with HAM/TSP can bring a new interpretation at how biological changes can influence inflammation and, consequently, the clinical status of these patients. In addition, it can open paths for future drug interventions supporting the control of this infection and improve the quality of life of these individuals.
Verifying the existence of associations between clinical (such as pain and functional capacity) and biological variables is important to try to fill-in gaps that still exist in relation to the pathophysiology of this neuroinfection. Thus, it will help to better understand what cell death regulatory mechanisms are beneath the pathology caused by HTLV, which in turn can influence the progression of this disease.
Conclusion
The results of this systematic review reveal a reduced number of studies regarding the HAM/TSP and apoptosis with several methodological issues. In addition, the heterogeneity of the samples and the absence of information on the selection and statistical measures of the sample turn it difficult to generalize the results. Therefore, it was impossible to carry out the meta-analysis. Overall, the intrinsic and extrinsic apoptotic pathways seem to play a role in disease progression stimulating more clinical studies on this issue.
Footnotes
Authors' Contributions
A.P.B.F.—conception and design of the study, data collection, analysis and interpretation of data, preparation and review of the article, approved the final version to be published. –A.P.P.C.—data collection, analysis and interpretation of data, review of the article, approved the final version to be published. P.M.—analysis, interpretation of data, review of the article, and correction of the text of final article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was financed in part by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior—Brasil (CAPES)—Finance code 001.