
Abstract
Interferon-induced transmembrane proteins (IFITMs) are transmembrane proteins induced by interferon that can provide broad-spectrum antiviral activities. However, there are few reports on the antiviral activity of monkey-derived IFITMs. In this study, the IFITM1 and IFITM3 genes of African green monkey (AGM) were cloned and overexpressed in Vero cells, followed by infection with mouse norovirus (MNV) and severe fever with thrombocytopenia syndrome virus (SFTSV). The results showed that monkey IFITM1 and IFITM3 can be stably overexpressed in Vero cells. Both IFITM1 and IFITM3 from AGM could effectively restrict infection by SFTSV, and the viral inhibition rate of IFITM3 was more obvious compared with IFITM1. However, both monkey IFITM1 and IFITM3 had no significant effect on the replication of MNV. These results indicate that different IFITMs have different functions, which may be related to the structure of the host IFITMs and the types of pathogens.
Introduction
Interferon-induced transmembrane proteins (IFITMs) can activate host innate immunity and resist the infection of external pathogens, especially IFITM1, IFITM2, and IFITM3, which can effectively inhibit the invasion and replication of many kinds of viruses (18,32). In 1996, researchers found that IFITM1 can effectively inhibit the replication of vesicular stomatitis virus (1). To date, it has been reported that IFITMs can not only inhibit the invasion of cells by influenza A virus but also significantly inhibit West Nile virus (WNV), Ebola virus (EBOV), SARS coronavirus (SARS-CoV), Rift Valley fever virus (RVFV), dengue virus (DENV), Semliki forest virus (SFV), Zika virus, respiratory syncytial virus (RSV), human immunodeficiency virus-1 (HIV-1), hepatitis C virus (HCV), reovirus, primate lentiviruses, and other capsular or noncapsular RNA viruses (4,6,9 –12,14,19,21,23 –25,29). Moreover, the IFITM proteins also exhibit antiviral activity against individual DNA viruses (17,21,33). For example, IFITM1 inhibits frog iridovirus (RGV) replication by preventing viral entry (33). IFITM1, 2, and 3 can inhibit the early stage of African swine fever virus (ASFV) infection (21). We previously found that IFITM3 protein restricts vaccinia virus (VACV) infection by interfering with virus binding and entry in a low pH-dependent manner, and VACV can also inhibit IFITM3 translation (17). IFITM3 can suppress H5N1 replication in the early stage of infection (11).
Nonhuman primates are evolutionarily closely related to humans. They share many common characteristics with humans in terms of physiology, behavior, and genetics. Therefore, they are widely used as models for human disease, behavior, and cognitive research (13). African green monkey (AGM) is a long-tailed monkey, mainly distributed in the South Saharan of Africa. Because AGMs are natural hosts of the simian immunodeficiency virus (SIV), they are often selected as animal models for AIDS research (27). AGMs are widely used in the study of zoonotic infectious diseases due to their sound immune system and virus adaptability (28). Recently, it was reported that primate lentiviruses are differentially inhibited by human IFITMs, and SIV derived from AGM is significantly inhibited by human IFITM2 as well as AGM-origin IFITM3 (23). However, the strong immune adaptation mechanism of AGM is not clear, and it is not clear whether AGM-origin IFITMs also participate in and play an important role in AGM against other viruses. In this study, the IFITM1 and IFITM3 genes were cloned from liver tissue of AGM, a eukaryotic expression vector was constructed, and protein was successfully expressed in the Vero cell line. The stable expression cell lines were screened, and the expression characteristics of the recombinant protein were identified. Mouse norovirus (MNV) and severe fever with thrombocytopenia syndrome virus (SFTSV) were used as exogenous viruses to verify the inhibitory effects of IFITM1 and IFITM3 on viral gene replication in vitro, which may provide a reference for further study on the immune protection mechanism of IFITMs.
Materials and Methods
Tissue, cells, and virus
The animal protocol for this experiment was approved by the Animal Care Committee of Laboratory Animal Center, Academy of Military Medical Sciences. Animals were maintained and processed following the Guide for the Care and Use of Laboratory Animals.
Liver tissues of AGM were obtained from Laboratory Animal Center, Academy of Military Medical Sciences, and stored at 80°C. SFTSV and MNV were provided by the Institute of Microbiology and Epidemiology, Academy of Military Medical Sciences.
AGM kidney epithelial cells (Vero) were grown in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum (Invitrogen) and 1% penicillin/streptomycin (Invitrogen) at 37°C in a 5% CO2 incubator.
Vero cells were inoculated with MNV and SFTSV at a ratio of 1:50. Culture supernatants were collected when cells were no longer attached to the culture flask. Viruses were determined by plaque formation and reverse-transcription quantitative PCR (RT-qPCR) and stored at −80°C.
Reverse-transcription PCR and real-time PCR
Total RNA was extracted from AGM livers or cells using the RNA Pure Tissue and Cell Kit (CWBIO, Beijing, China) according to the manufacturer's instructions. Then, total RNA was used to synthesize cDNA by using oligo-dT primer and PrimeScriptRT Enzyme (TaKaRa, Dalian, China).
To amplify IFITM1 and IFITM3 genes, PCR was performed using the synthesized cDNA as the template in a 50 μL volume of PCR solution containing 0.5 U of PrimeSTAR® HS DNA Polymerase (TaKaRa). PCR conditions were as follows: predenaturation at 98°C for 3 min; 30 cycles of 98°C denaturation for 10 s, 60°C annealing for 15 s (IFITM1) or 65°C for 5 s (IFITM3), and 72°C extension for 30 s; and one cycle of 72°C for 5 min. PCR products were purified using the DNA Agarose Gel Recovery Kit (TIANGEN, Beijing, China) and cloned into the pGEM-T vector (Promega). The recombinant plasmids, pGEM-T-IFITM1, and pGEM-T-IFITM3 were transferred into DE3 competent cells (QUANSHIJIN, Beijing, China), and positive clones were identified using restriction analysis and sequencing. The primers used in this study are listed in Table 1.
Primers Used in This Study
IFITM, interferon-induced transmembrane protein; RT-qPCR, reverse-transcription quantitative PCR.
To analyze gene expression, RT-qPCR was performed using SYBR Green Premix Ex Taq II (TaKaRa). Samples were normalized based on the expression of the gene encoding human GAPDH as a reference. The cycling conditions consisted of initial denaturation for 3 min at 95°C and 39 cycles of 5 s at 95°C and 30 s at 60°C for IFITM1 or 30 s at 65°C for IFITM3. To verify the lack of nonspecific amplification, following the completion of RT-qPCR, a melting curve analysis was performed. The melting protocol consisted of heating from 65°C to 95°C at a rate of 0.5°C per step, and each step was held for 1 s for data acquisition. Each sample was repeated three times.
Viral RNA was extracted from cells according to the instructions of the QIAamp MinElute Virus Spin Kit (QIAGEN), and RT-qPCR was performed as follows. The cycling conditions consisted of initial denaturation for 3 min at 95°C, followed by 35 cycles of 5 s at 95°C, and 30 s at 62°C for VP1 of MNV or 30 s at 65°C for NSs of SFTSV. The copies of viruses were calculated according to the standard curve. To construct a standard plasmid, VP1 of MNV and NSs of SFTSV were obtained by PCR and cloned into pGEM-T Easy Vector. Plasmids were diluted to a series of 101–1011 copies/μL, followed by RT-qPCR to generate the standard curve. The experiment was performed at least three times.
Construction of expression plasmids
The IFITM1 and IFITM3 genes were digested from pGEM-T-IFITM1 and pGEM-T-IFITM3, respectively, by EcoR I and BamH I, and subcloned into the expression vector plv-EGFP (Inovogen Tech, Beijing, China), resulting in recombinant plasmids IFITM1-EGFP and IFITM3-EGFP, respectively. Then, the recombinant plasmids, as well as plv-EGFP, were transferred into DE3 competent cells and verified by PCR and sequencing.
Cell transfection and immunofluorescence
Vero cells were cultured in DMEM containing 10% FBS and 1% penicillin/streptomycin at 37°C and 5% carbon dioxide. When the cells reached 80% confluence, they were transfected with 10 μg of IFITM1-EGFP, IFITM3-EGFP, or plv-EGFP using the Lipofectamine 2000 Kit (Invitrogen) according to the manufacturer's instructions. Then, cells were cultured with DMEM containing an appropriate concentration of puromycin in an incubator at 37°C with 5% carbon dioxide.
Positive clones were verified by immunofluorescence (IFA). Briefly, when cells were stable, 10 μL of Hoechst (Hoechst AG, GER) was added into each well and mixed gently. The cells were cultured in the incubator according to the above conditions for 10 min. Then, the medium containing the Hoechst was removed, and each well was washed three times with sterile phosphate-buffered saline (PBS). Subcellular localization of IFITMs-EGFP was visualized by using a Nikon Eclipse TS100 inverted microscope (Nikon, Tokyo, Japan). The resulting cells were designated as Vero-IFITM1 and Vero-IFITM3.
Western blot
To determine the expression of IFITM1 and IFITM3 proteins, total proteins were extracted with RIPA buffer with protease inhibitor cocktail (Roche, SWIT). The samples were electrophoresed by sodium dodecyl sulfate/polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to nitrocellulose blotting membrane (GE Healthcare, GER). After incubating in blocking buffer (5% bovine serum albumin in Tris-buffered saline) at room temperature for 1 h, the samples of membrane were probed with Anti-IFITM3 (1:500; Proteintech), Anti-IFITM1 (1:1,000; Proteintech) or actin antibody (1:5,000, MBL, catalog no. PM053MS). After washing three times with PBS, the membrane was incubated with secondary antibody, anti-rabbit IgG antibody (1:10,000; Jackson ImmunoResearch), and anti-mouse IgG antibody (1:10,000; Jackson ImmunoResearch). The bands were then visualized using the Enhanced Chemiluminescence Detection Kit (Pierce Biotechnology, Rockford, IL) and X-ray film and quantified using ImageJ software with normalization to GAPDH.
Plaque assay
MNV and SFTSV were diluted with a concentration gradient of 10−1 to 10−6. Vero cells were incubated with MNV and SFTSV at the designated PFU/cell for 2 h in a 37°C incubator containing 5% CO2, washed with PBS, and cultured with complete DMEM containing 1% methylcellulose for 3 d. Thereafter, cells were fixed with 10% paraformaldehyde and stained with Crystal Violet. The virulence of the virus was analyzed by plaque analysis.
Viral infection assays
Vero-IFITM1 and Vero-IFITM3 cells were infected, respectively, with MNV and SFTSV at the indicated multiplicity of infection for 1 h. Vero-vector cells were used as positive controls. The inoculant was removed, and the cells were washed with cold PBS. Fresh complete culture medium was added. Cells were harvested at various times after inoculation, and viral infection was determined by quantitative PCR.
Statistical analysis
Data were calculated as the mean ± standard deviation. Student's t-tests were performed for all analyses using GraphPad Prism 7 software (GraphPad Software, Inc., La Jolla, CA). Differences among groups were determined by one-way ANOVA with a repeated measures test. Statistical significance was determined by two-tailed p values: *p < 0.05, **p < 0.01, ***p < 0.001, and ****p < 0.0001.
Results
IFITM1 and three genes were cloned from AGM
Both IFITM1 and IFITM3 genes of AGM were amplified (Fig. 1) and successfully cloned into pGEM-T vector, generating pGEM-T-IFITM1 and pGEM-T-IFITM3, respectively. As shown in Figure 1a, the length of the IFITM1 gene was 378 bp, while that of IFITM3 was 441 bp. The sequencing results showed that there were two mutations in the IFITM1 gene and four mutations in IFITM3 compared with the target sequence (Fig. 1b). Further analysis showed that monkey IFITM1 contained 125 amino acids and IFITM3 contained 146 amino acids. Unlike in the target sequence, the 13th amino acid of IFITM1 is proline instead of alanine, and the 111th is phenylalanine instead of isoleucine. The 15th amino acid of IFITM3 is serine instead of glycine, and the 135th amino acid is isoleucine instead of valine (Fig. 1c).
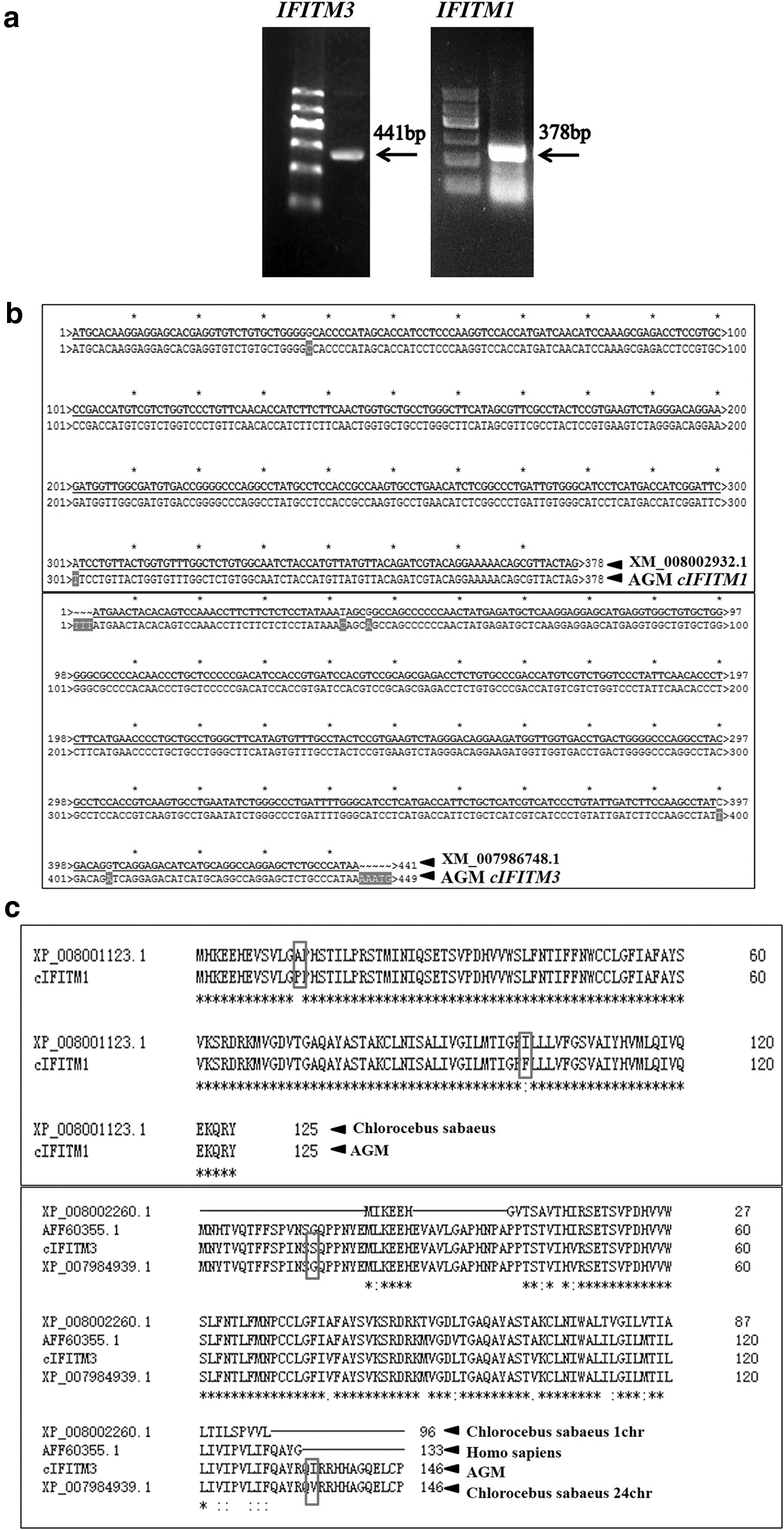
Amplification and cloning of monkey IFITM1 and IFITM3 genes.
Monkey IFITM1 and IFITM3 were overexpressed in Vero cells
The IFITM genes were digested from pGEM-T-IFITM1 and pGEM-T-IFITM3 and subcloned into plv-EGFP, resulting in the recombinant plasmids IFITM1-EGFP and IFITM3-EGFP, respectively. Then, the expression plasmids were transfected into Vero cells, followed by evaluating the expression by RT-qPCR, IFA, and Western blotting.
The RT-qPCR results showed that the transcription levels of the IFITM1 and IFITM3 genes were significantly higher in the transfected cells than in the normal cells (Fig. 2a). Then, these results were further confirmed using IFA. As shown in the figure, the fluorescence signal of EGFP proteins in the transfected cells was obvious (Fig. 2b), which indicated that the plv-EGFP, plv-IFITM1, and plv-IFITM3 plasmids were expressed normally in Vero cells. The western blotting results indicated that the IFITM1 and IFITM3 proteins were overexpressed in the transfected cells, and their expression was significantly increased compared with that in the control groups (Fig. 2c).
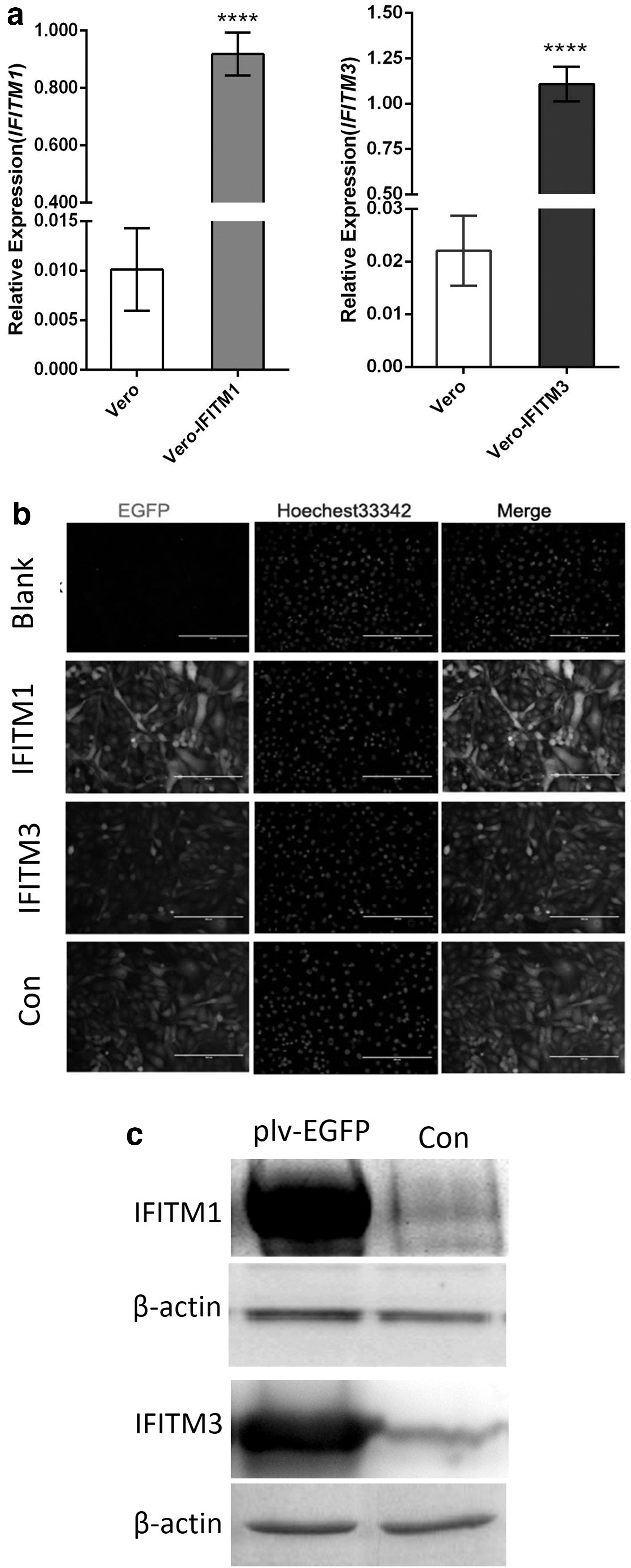
Overexpression of monkey IFITMs.
Monkey IFITMs interfered with the replication of SFTSV rather than MNV
To evaluate viral infection, plaque formation of Vero cells infected with the viruses was analyzed by Crystal Violet staining. As shown in Figure 3a, cytopathic effects were observed in the virus-infected cells, as plaques with different shapes and sizes appeared in these cells. Moreover, plaque number was reduced with decreasing amounts of inoculated virus (Fig. 3b).
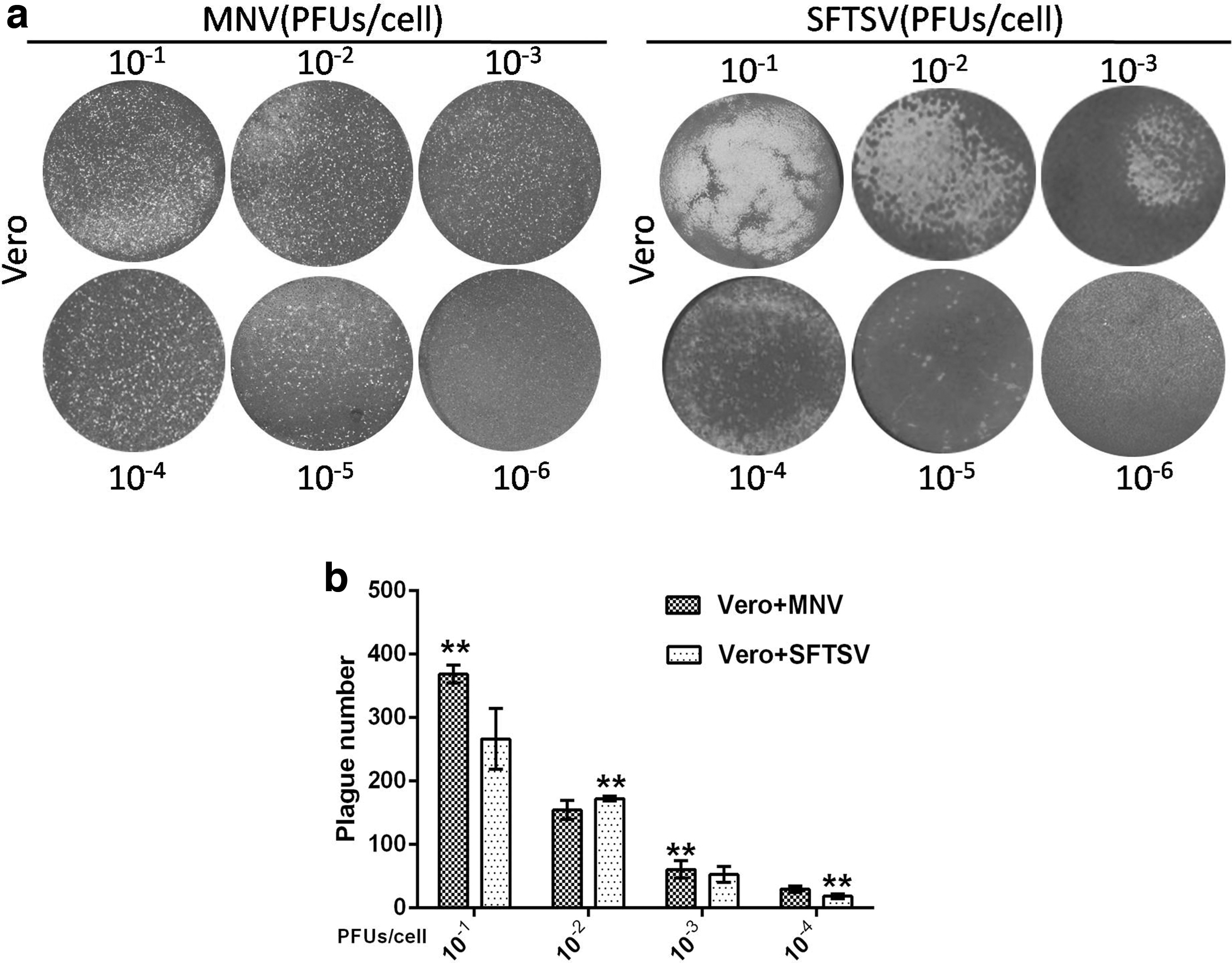
Results of plaque analysis. Cells were infected with viruses at the designated PFU/cell for 2 h.
To evaluate the inhibitory effect of monkey IFITMs on the viruses, RT-qPCR was performed to detect the expression of the VP1 gene in MNV and NSs gene in SFTSV. The results showed that the expression levels of the VP1 gene in Vero-IFITM1 and Vero-IFITM3 cells had no significant difference compared with the level in the control group, and their proliferation amount was 108/copies (Fig. 4a, b), suggesting that monkey IFITM1 and IFITM3 do not affect the proliferation of MNV. The levels of NSs gene expression in Vero-IFITM1 and Vero-IFITM3 cells were 103–104/copies, which was significantly reduced by 100–1,000 times compared with that in the control group, indicating that monkey IFITMs can inhibit SFTSV infection (Fig. 4c, d). Furthermore, the copy numbers of the NSs gene in Vero-IFITM3 cells were ∼10 times lower than those in Vero-IFITM1 cells, and the proliferation speed of the NSs gene was relatively slow. Therefore, the inhibitory effect of IFITM3 on SFTSV infection may be higher compared with IFITM1.
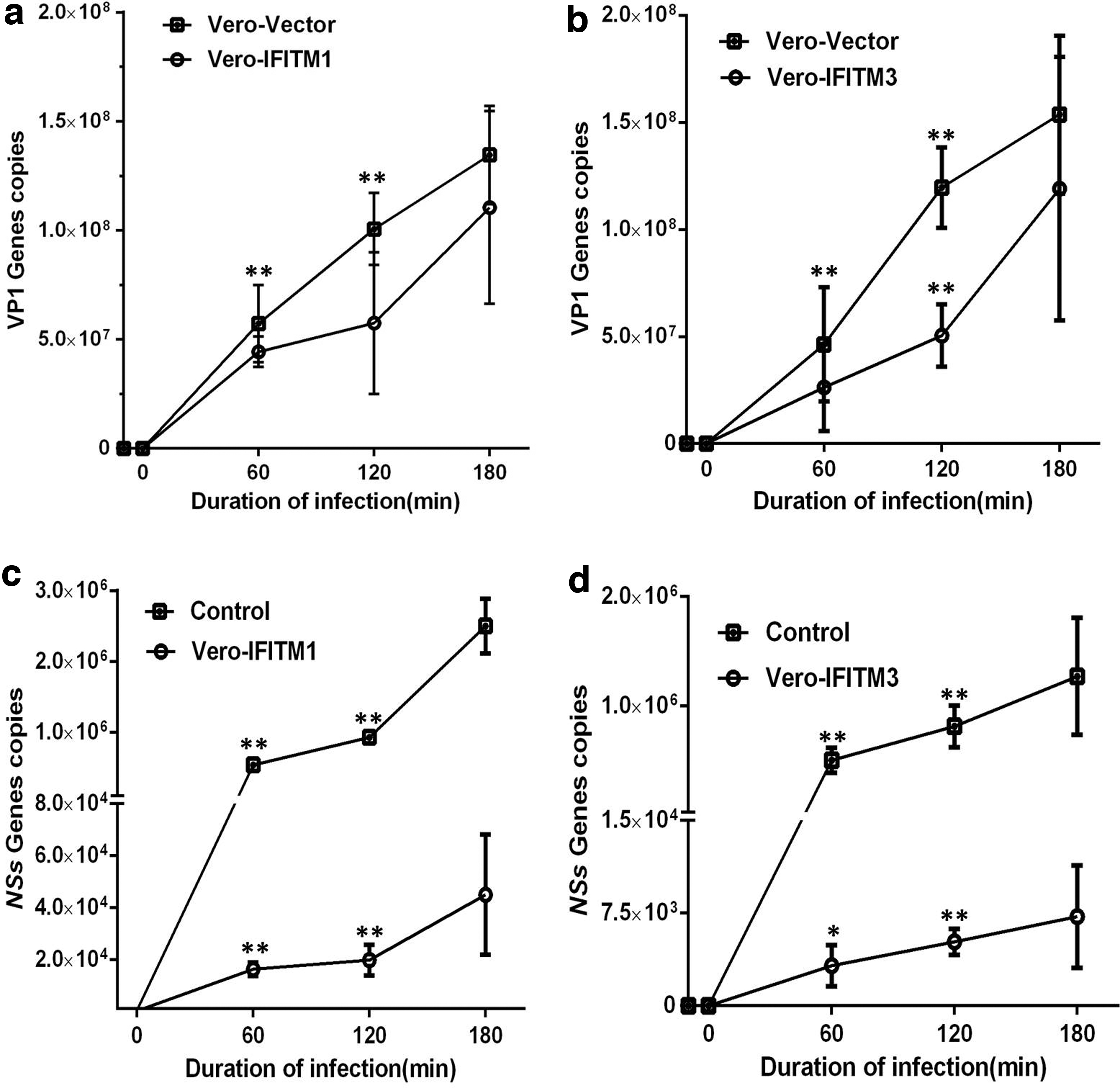
Inhibitory effect of monkey IFITM1 and IFITM3 on MNV and SFTSV. Cells were infected with MNV (MOI = 5) or SFTSV (MOI = 2) for 3 h, followed by evaluation using RT-qPCR. The copy numbers were calculated by referring to the corresponding standard curve.
Discussion
AGM is a nonhuman primate model animal that has been widely used in recent years. It plays an important role in research on antiviral medicine. AGMs carry SIV but seldom become sick, indicating that they have a perfect natural immune system (2). As an endogenous host-limiting factor, IFITMs have broad-spectrum antiviral effects (3) and may also play an important role in immune regulation in AGM. In this study, IFITM1 and IFITM3 cloned from the liver of AGMs had individual amino acid site mutations in the target protein compared with that of the reference sequence. IFITM1 contained 125 amino acids; alanine at the 13th position was proline, and isoleucine at the 111th position was phenylalanine. IFITM3 contained 146 amino acids; glycine at position 15 was serine, and valine was at position 135 instead of isoleucine (Fig. 1c). The mutation sites were located in the N-terminal hydrophobic domain (NTD) and C-terminal variable domain (CTD) of the protein (5), which did not affect CD225. CD225 is a highly conserved region of IFITMs. Some researchers reported that the CD225 region is the core functional region of IFITMs in virus inhibition, whereas the NTD and CTD of the protein influenced the topological structure and cell location of IFITMs (7,15,31). According to our results, these amino acid mutations did not affect the function of IFITMs.
The virus inhibition spectrum of IFITMs is very broad, and they can inhibit both enveloped RNA virus and nonenveloped RNA virus (4,6,9 –12,14,19,21 –25,29). It has been reported that IFITM1 does not affect the invasion of some viruses (14,26,30). Mudhasani et al. found that both IFITM2 and IFITM3 can effectively inhibit the RVF virus, whereas IFITM-1 has no inhibitory effect (20). In this study, we selected a single-strand positive RNA virus without an envelope, MNV, and a single-strand negative RNA virus with an envelope, a new Bunia virus, also known as SFTSV, as infectious pathogens, to verify the antiviral effect of AGM IFITMs. The results showed that the NSs gene of SFTSV in AGM IFITM1 and IFITM3 overexpression cell lines decreased 1,000-fold compared with that in the control cells (Fig. 4c, d), indicating that SFTSV particles were inhibited by IFITM1 and IFITM3 when entering the cell membrane. Furthermore, the inhibition of IFITM3 was significantly higher compared with IFITM1 (Fig. 4), indicating that different IFITMs have different functions. These functional differences may be related to the structure of the host-limiting factors IFITM1 and IFITM3 and the types of pathogens.
IFITMs are host-limiting factors that play an important role when the virus enters the host. They are the first line of defense for host cells to resist viral infection and are mainly inhibited in the early stage of virus replication (3,11,17). However, some viruses can evade the restriction of IFITMs and even hijack antiviral proteins to promote their infection of host cells or assembly of virosomes, probably by regulating membrane fusion (24,28,29,32). In this study, we also found that the replication level of the VP1 gene of MNV in AGM IFITM1 and IFITM3 overexpression cell lines was similar to that in normal cells, indicating that they did not strongly interfere with the replication of MNV particles (Fig. 4a, b). The possible reasons are that the uncoated nature of MNV helps it enter the host cell better, or the adaptation mechanism of the high variability of MNV may allow it to escape the defense line of innate immunity of the host (8,16). However, the exact mechanism needs to be further studied.
Footnotes
Authors' Contributions
J.Y.B. and L.Z.R. conceived and designed the study; acquisition, analysis, and interpretation of data were performed by S.H.Z., C.L., L.Y., T.T.F., J.W.F., and H.W.X.; J.Y.B. and L.Z.R. drafted or revised the article; and final approval of the version to be published was given by H.W.X., J.W.F., L.Y., T.T.F., C.L., S.H.Z., L.Z.R. and J.Y.B. H.W.X, J.W.F., L.Y., T.T.F., C.L., S.H.Z., L.Z.R., and J.Y.B. are accountable for all aspects of the work and for ensuring that questions related to the accuracy or integrity of any part of the work are appropriately investigated and resolved. All authors read and approved the final version of the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the New Drug Project of China (No. 2018ZX09J18105-006 and No. 2018ZX10101003), the National Natural Science Foundation of China (No. 31772747), and the Science and Technology Research Program during the 13th Five-year Plan Period of Jilin Educational Committee (No. JJKH20190172KJ). The sponsors of the study had no role in the study design, data collection, data analysis, interpretation, or writing of the article.