
Abstract
An efficient method for detection of foot and mouth disease (FMD) and, particularly, differentiation of vaccinated from infected animals is the use of nonstructural (NS) proteins as antigens in Enzyme-Linked Immunosorbent Assay (ELISA) Kits. In this study, only epitopic regions of 3AB and 3D NS proteins were used for recombinant protein production, as a cost-effective method instead of peptide synthesis, for application in in-house ELISA diagnostic kits. Specific primers were designed according to the antigenic regions of 3AB (C-terminus of 3A and the whole 3B) and 3D (N-terminus) proteins, and the polymerase chain reaction (PCR) amplification was performed. Purified amplicons were cloned into pET21a (+) vectors and then transformed into Escherichia coli (BL21). Thereafter, bacteria were induced with 1 mM isopropyl β-d-1-thiogalactopyranoside (IPTG) for expression of antigenic proteins. Antigenic 3AB protein was expressed in soluble form, but 3D protein was extracted from the bacterial lysate. Protein expression was confirmed using sodium dodecyl sulfate–polyacrylamide gel electrophoresis and Western blot analyses. An indirect ELISA was developed for each protein, and the diagnostic sensitivity and specificity were determined. The 3AB-ELISA showed higher sensitivity and specificity than 3D-ELISA (95.24% and 100%, compared with 90.48% and 88.71%, respectively). The epitopic 3AB-ELISA developed here can be used for detection and differentiation of FMD infected from vaccinated animals, but the epitopic 3D-ELISA showed lower efficiency in screening for FMD status.
Introduction
Foot and mouth disease virus (FMDV) infection is the cause of a highly transmittable disease in cloven-hoofed animals. FMDV is a member of family Picornaviridae and has a single-stranded positive-sense RNA genome. This genomic RNA encodes a polyprotein that yields eight nonstructural (NS) proteins (L, 2A, 2B, 2C, 3A, 3B, 3C, and 3D) and four structural components of the virion (vp1–vp4) (3,30). FMD can easily be transmitted through the aerosol or even animal–animal contacts (9). The virus can also be transmitted to calves through the milk of dairy cattle and through semen, urine, and feces to other cattle (1,33).
Factors such as the number of virus serotypes, high mutation rate in the viral genome (resulting in rapid changes in antigenicity), high survival rate of the virus in nature, rapid outbreaks, formation of carrier animals, high disease control costs, and strict international regulations for countries with endemic status increase the importance of FMD (11).
Several tools are of critical importance to the global FMD control strategy. These include the use of vaccines and the timely and prompt diagnosis of the disease. Vaccination with vaccines containing the entire inactivated virus, or recombinant vaccines, can protect against a specific serotype of the seven known serotypes (2). Since the disease can remain latent in the host for a long time leading to emergence of carrier animals, one way to control the disease is to differentiate FMDV infected from vaccinated animals (DIVA). In infected animals, antibodies are produced against both structural and NS proteins, while vaccinated animals produce only antibodies against structural proteins. Therefore, detection of antibodies to NS proteins can be a way for DIVA (4).
Diagnostic kits for detection of viral structural proteins are commercially available. These kits and assays are serotype specific and mostly are used to identify virus serotype in surveillance programs (10,35). However, the use of NS 3AB, 3ABC, 3A, or 3B antigens has been used in diagnostic tests for DIVA, regardless of virus serotypes (6,15,21,26,31). The 3D protein is an immunogenic and the most conserved protein in all FMD serotypes. Antibodies against 3D protein can be detected for a longer period in infected animals. Hence, detection of 3D protein antibodies can be useful for testing FMD status of animals before export/import, epidemiological studies, determination of the FMDV spread in animal populations, and also to identify carrier animals (7,13,25).
The 3AB protein induces high immunity after infection with FMD virus and is highly conserved (19,29). Most commercial Enzyme-Linked Immunosorbent Assay (ELISA) Kits are based on 3ABC proteins; however, it has been demonstrated that the bovine linear B cell epitopes mostly rely on the C-terminal region of 3A and the whole sequence of 3B proteins. No B cell epitope was detected in 3C protein (14). The 3C protein has protease activity (8), which seems to affect recombinant 3ABC protein production in Escherichia coli, leading to cleavage in 3ABC whole protein and reduction in production efficiency (28). Epitopic peptides have been also detected in the N-terminus of the 3D protein (14,37).
In the present study, instead of using the entire sequence of 3A, 3B, and 3D genes, only gene sequences encoding antigenic regions of 3AB and 3D NS proteins were used for cloning and recombinant protein production, to be applied in in-house ELISA diagnostic kits.
Materials and Methods
All blood sample collections from cows were performed in accordance with animal ethics guidelines and protocols.
Cell lines and viruses
The BHK-21 cells infected with FMDV O serotype/PanAsia strain were obtained from the Department of Foot and Mouth Disease Vaccine Production, Razi Vaccine and Serum Research Institute, Karaj, Iran.
Primer design
The FMDV serotype O poly-protein coding sequence in NCBI database (accession number: AH012985.2) was used to design specific primers according to the antigenic regions of 3AB (C-terminus of 3A and the whole 3B) and 3D (N-terminus) proteins (14,37). Primers were designed using Oligo7 software with recognition sites for restriction enzymes. The sequences of the primers were as follows: 3AB-F: 5′-agaca
Cloning and expression of recombinant proteins
Viral RNA was isolated using Blood/Cultured Cell Total RNA Purification Mini Kit (Favorgen, Taiwan) from FMDV infected BHK-21 cells and reverse transcribed. The cDNA was used as template for PCR amplification of 3AB and 3D truncated sequences with expected fragment length of 415 and 560 bp, respectively. PCR was performed using Pfu DNA polymerase with the following condition: initial denaturation at 94°C for 5 min, 35 cycles of denaturation at 94°C for 30 sec, annealing at 58°C for 40 sec and extension at 72°C for 60 sec, and a final extension at 72°C for 10 min. After purification of amplified fragments using Gel/PCR Purification Mini Kit (Favorgen, Taiwan), DNA fragments were cloned into pET-21a (+) vectors. The recombinant plasmids were then transformed into E. coli (DH5α) hosts and cultured in LB-agar medium containing 100 μg/mL ampicillin. Transformants were screened, and the positive clones were further analyzed by colony-PCR and restriction enzyme digestion to determine the presence of the inserted sequences.
The accuracy and reading frame of genes in the selected recombinant clones were confirmed by sequencing in both senses. The resulting sequences were used in multiple sequence alignment using BLAST (Basic Local Alignment Search Tool) and were compared with polypeptide sequences of FMDV O serotype, available in NCBI database. After confirmation of the sequences, accession numbers were assessed in database.
The recombinant vectors were transformed into E. coli (BL21) hosts, and protein expression was induced with 1 mM isopropyl β-d-1-thiogalactopyranoside (IPTG) at 37°C for 2, 3, and 4 h. Cultures were precipitated, and the pellets were treated with 50 mM NaH2PO4, 300 mM NaCl, and 0.5 mM PMSF (phenylmethylsulfonyl fluoride). The suspension was centrifuged at 10,000 rpm, 4°C for 10 min, and the supernatant was collected. The resulting pellet was treated with denaturing buffer (50 mM NaH2PO4, 300 mM NaCl and 8 M urea) for 30 min, and the supernatant was collected after centrifugation at 10,000 rpm, 4°C for 15 min. All samples were analyzed on 12% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE).
Purification of recombinant antigenic 3AB and 3D
The recombinant proteins were produced as fusion proteins with a C-terminal 6 × His-tag and were purified by metal affinity chromatography using Ni-NTA Superflow Columns (Qiagen, Germany) according to the manufacturer's instruction. Purified 3AB and 3D proteins were analyzed on 12% SDS-PAGE. Concentration of both proteins was calculated by BradFord assay. Percentage of hydrophilicity was determined for both proteins by Peptide Calculator software.
Western blot analysis
The presence of His-tagged recombinant proteins was tested by western blotting using anti-6 × His antibody. For Western blotting, the purified proteins were electrotransferred onto a polyvinylidene fluoride membrane for 16 h at 4°C and 20 mA. The membrane was blocked for 2 h. After several washes, the membrane was incubated with Monoclonal Anti-polyHistidine-Peroxidase clone HIS-1 (1: 10,000, Sigma–Aldrich, St. Louis, MO). The reaction was visualized with (3,3′-diaminobenzidine tetrahydrochloride) DAB substrate (Sigma–Aldrich).
Standardization of 3AB and 3D indirect ELISA
For detection of antibodies against the epitopic recombinant 3AB and 3D proteins, indirect ELISA was implemented. To optimize the ELISA condition, checkerboard titration was performed using serial dilutions of antigens and sera. A known FMD-positive serum from an infected cattle (previously tested by IZSLER 3ABC-ELISA, Italy) was used as a positive control, and a negative serum from a naive cattle (not infected or vaccinated) was used as a negative control. ELISA plates were coated with double dilutions of 0.25 to 4 μg/mL 3AB or 3D purified proteins in carbonate buffer, pH 9.6, and incubated overnight at 4°C. Plates were washed with phosphate buffered saline (PBS) containing 0.05% Tween 20 (PBST) for five times. Then, unbound sites were blocked using PBST with 2% bovine serum albumin (BSA), for 1 h at room temperature.
Serum samples were diluted 1:400 to 1:3200 for 3AB and 1:50 to 1:400 for 3D in PBST containing 1% BSA, added to wells and incubated for 30 min at room temperature. After five washes, horseradish peroxidase (HRP)-conjugated goat anti-bovine antibody (Sigma–Aldrich) was added to the wells in the same dilution buffer at appropriate working concentration and incubated at 37°C for 30 min. After five washes, chromogen/substrate mixture (3, 30, 5, 50-tetramethyl-benzidine [TMB]/H2O2) was added until the color was developed. After 15 min, the reaction was stopped by the addition of 2 M H2SO4. The optical density (OD) of each well was read at 450 nm using a microplate reader (BioTek). Antigen and serum dilutions that yielded OD values with a big increase compared with the previous dilution, focusing on lower usage of antigen amount, were chosen as the optimum dilutions for each ELISA set.
Screening of serum samples
For determination of cutoff values, the study involved a total of 30 samples of cattle sera collected from nonvaccinated noninfected cows from Razi Vaccine and Serum Research Institute. Infected and vaccinated sera were collected from farms of Ghom, Karaj, and Shahriar cities. Cutoff values were calculated as the mean value of negative sera ±2 standard deviation. To estimate diagnostic sensitivity, sera of 42 infected cows (36 cows with serotype O and 6 cows with serotype A) were collected and tested by our in-house 3AB and 3D assays, and the diagnostic sensitivity was estimated using the following formula: (true positive/[true positive + false negative]) × 100. Diagnostic specificity was also calculated using serum samples of 30 healthy cows and 32 A15-vaccinated cows according to the following formula: (true negative/[true negative + false positive]) × 100. All samples were tested in duplicates.
Results
Cloning of 3D and 3AB partial sequences
The PCR products of 3AB (415 bp) and 3D (560 bp) partial sequences were observed on 1% agarose gel (Fig. 1). Fragments were successfully cloned into pET-21a (+) vectors, sequencing confirmed the identity of sequences, and accession numbers were assigned (MK814881 for 3D and MK814882 for 3AB). Multiple sequence alignment showed that 3AB gene sequence had the highest nucleotide identity (97%) with Saudi Arabia serotype A (KU127247.1) and Bangladesh serotype A (MK088171.1) and also showed 93% identity with United Arab Emirates serotype O (G983737.1). The highest similarity was observed for 3D gene sequence with Saudi Arabia serotype A (KU127247.1) (99%), Bangladesh serotype A (MK088171.1) (97%), and Bangladesh serotype O (KX712091.1) (93%).
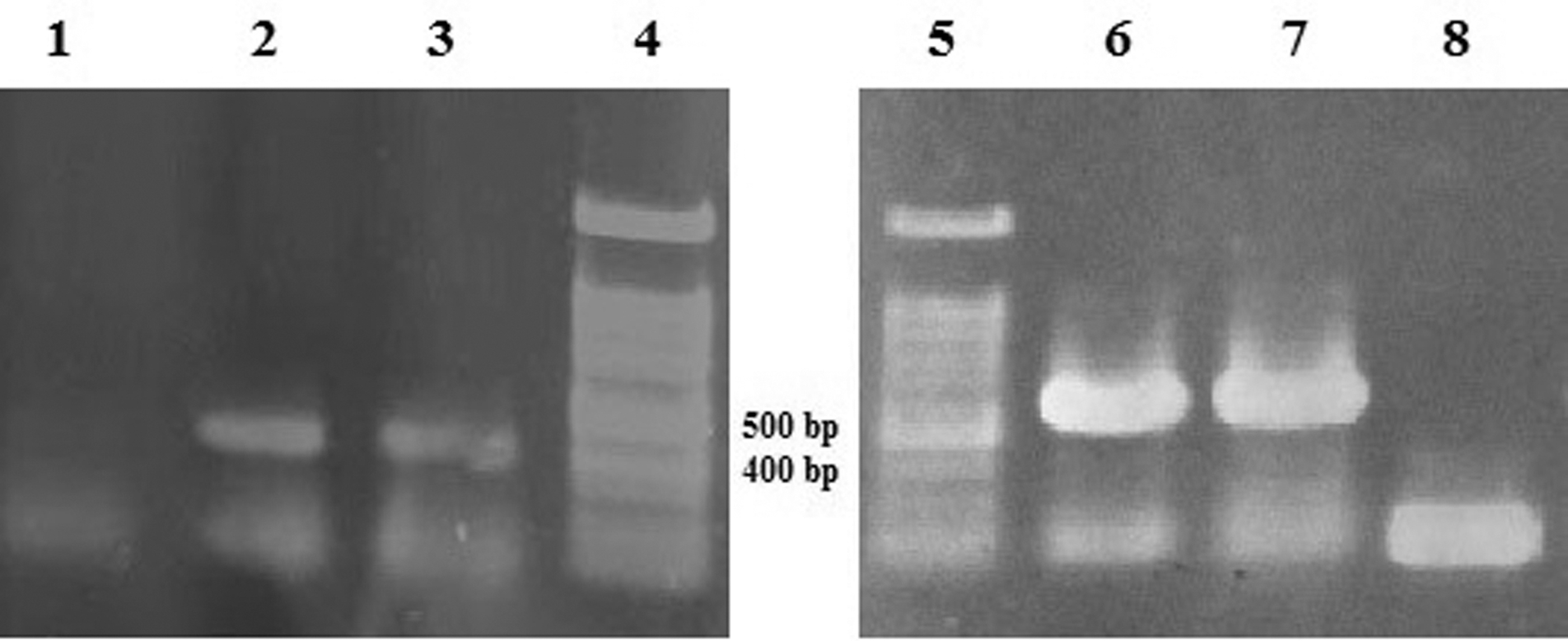
The PCR products of 3AB and 3D. Lane 1: negative control; Lanes 2 and 3: 415-bp fragments of 3AB; Lanes 4 and 5: 100-bp DNA ladder; Lanes 6 and 7: 560-bp fragments of 3D; Lane 8: negative control. PCR, polymerase chain reaction.
Expression of antigenic 3AB and 3D proteins
Protein expression was induced with 1 mM IPTG and screened at time intervals of 2, 3, and 4 h. SDS-PAGE (12% polyacrylamide gel) analysis of samples showed that the highest level of recombinant protein expression was achieved at 3 and 4 h postinduction for 3AB and 3D, respectively (Figs. 2 and 3). Estimated molecular weight was about 24 kDa for both antigenic recombinant proteins. The 3D protein was insoluble and purified under denaturing condition, but 3AB was extracted in bacterial supernatants, and this was somehow in accordance with percentage of hydrophilicity calculated for 3AB (0.45%) and 3D (0.27). However, expression and purification efficiency was high for 3D protein, and the purified protein concentration was 80 mg/L of induced culture, where 3AB was purified at the concentration of 15 mg/L of induced culture. Western blotting showed distinct protein bands and further confirmed the presence and the size of 6 × His-fused recombinant proteins (Fig. 4).

SDS-PAGE analysis of recombinant 3AB expression and purification. Arrow shows the recombinant protein.
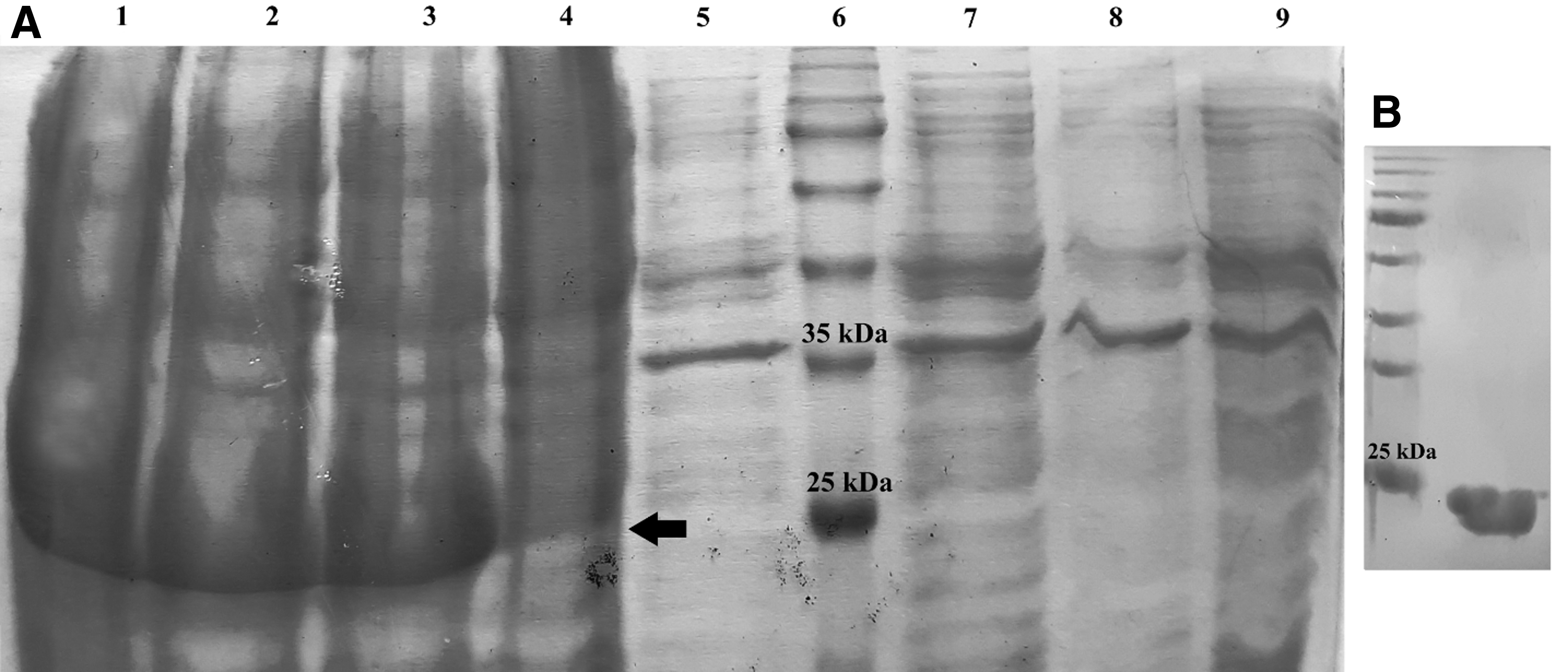
SDS-PAGE analysis of recombinant 3D expression and purification. Arrow shows the recombinant protein.
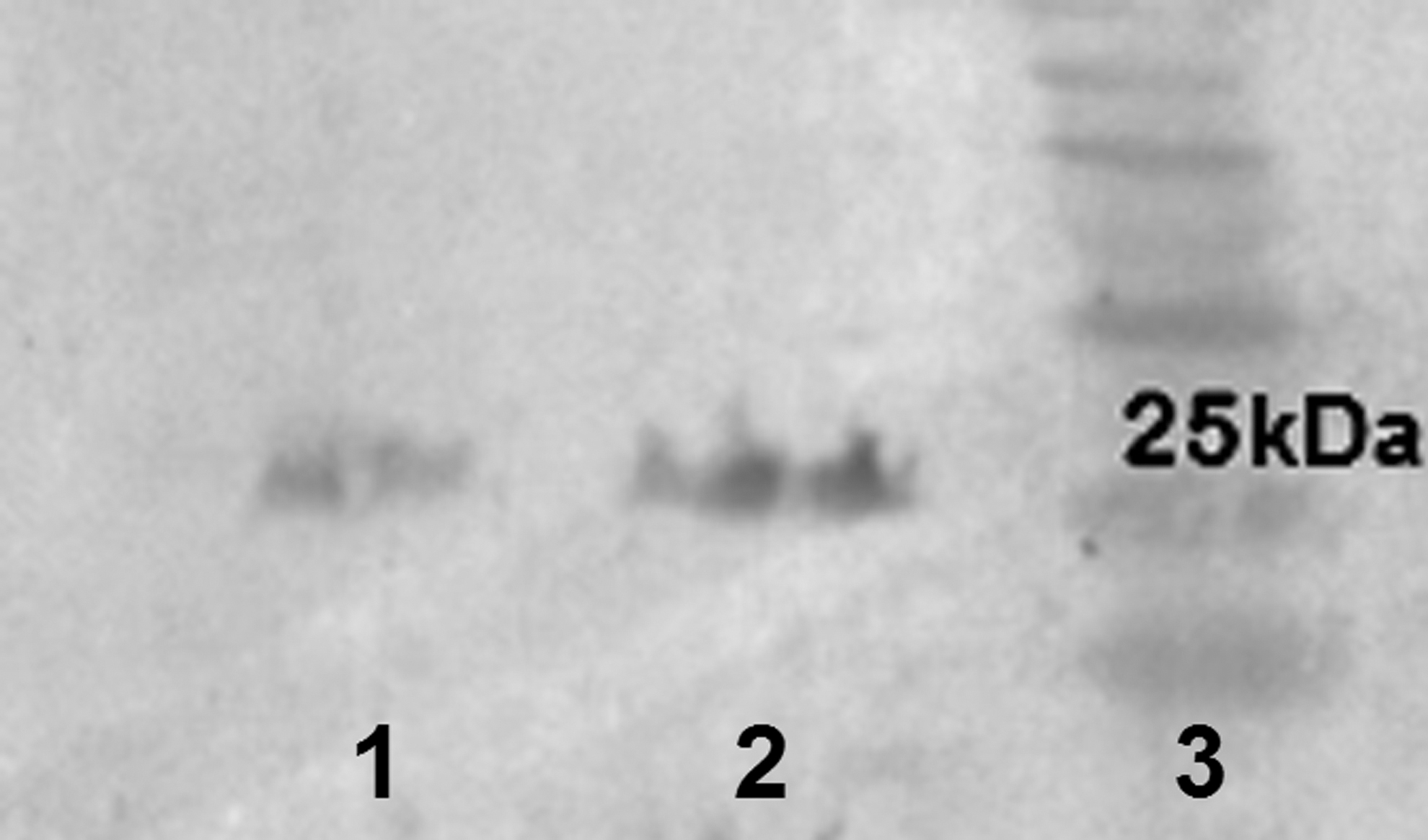
Western blot result. Lane 1: 3AB protein. Lane 2: 3D protein. Lane 3: 10–180 kDa protein ladder.
Indirect ELISA optimization and cutoff determination
The optimal antigen concentration and serum dilution for both 3AB-ELISA and 3D-ELISA were determined by checkerboard titration (Fig. 5). The results showed that the antigen concentration of 0.5 μg/mL and serum dilution of 1:400 were the optimal values for 3AB-ELISA. In addition, the antigen concentration of 2 μg/mL and serum dilution of 1:50 were selected as the optimal values for 3D-ELISA. These values were considered for detection of test sera.
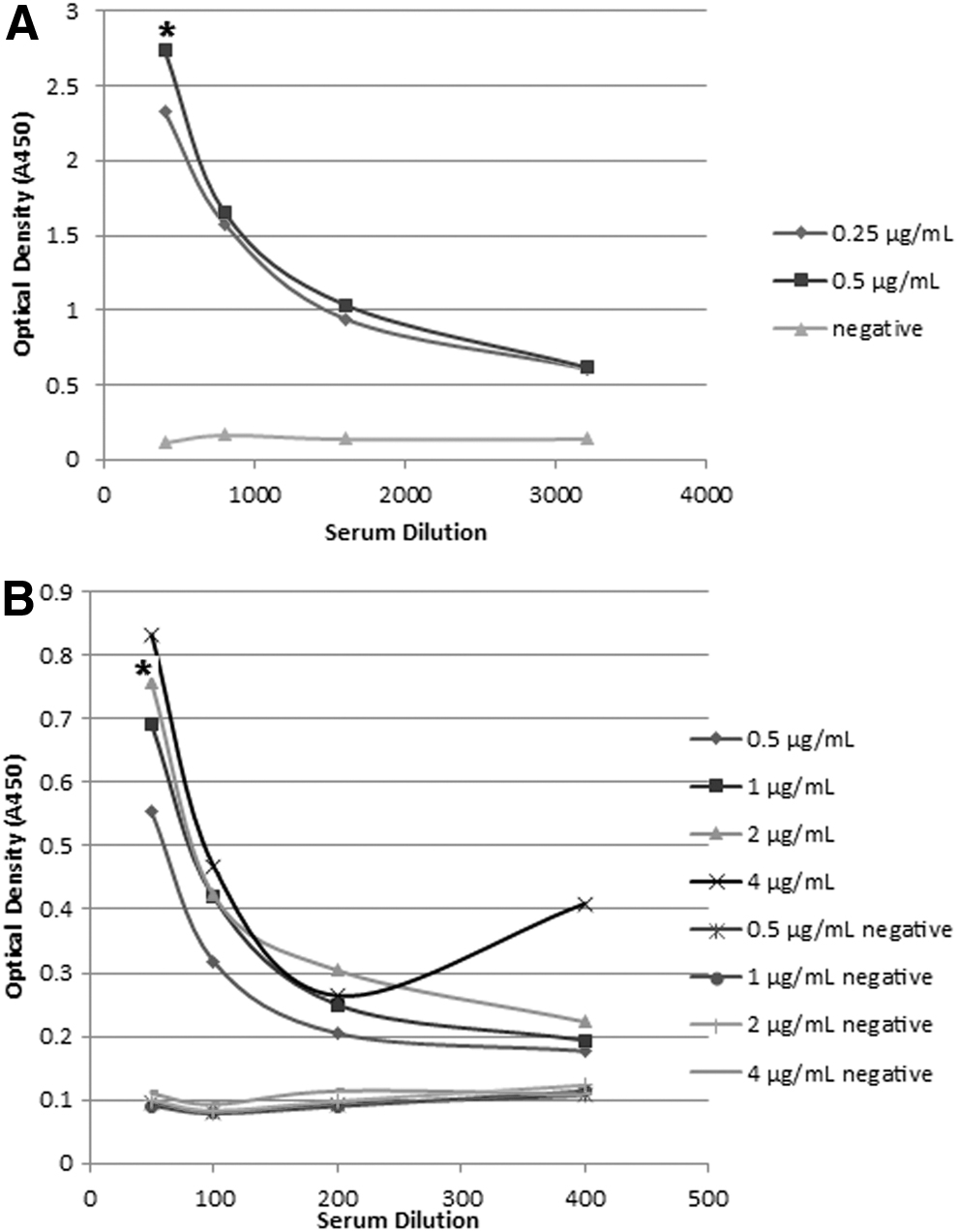
Reactivity of control sera at different dilutions with epitopic recombinant proteins 3AB
For determination of cutoff values, 30 negative sera were tested, and OD mean values of 0.182 (±0.025) and 0.125 (±0.035) were obtained for 3AB and 3D antigens, respectively. According to the data, cutoff values of 0.24 and 0.2 were considered for 3AB-ELISA and 3D-ELISA, respectively.
Antibody response against 3AB and 3D
At a similar antigen concentration and serum dilution, anti-3AB antibodies showed higher titer than anti-3D antibodies, obtained by OD value. Among 42 infected serum samples, 40 (34 O and 6 A) and 38 (32 O and 6 A) samples were positive for 3AB and 3D assays, respectively. In addition, among 62 negative (A15-vaccinated and normal) sera, 62 and 55 samples were detected to be negative for 3AB and 3D assays, respectively. Diagnostic sensitivity and specificity for 3AB assay were 95.24% and 100%, respectively. The sensitivity was 94.44% and 100% for serotypes O and A, respectively. For 3D-ELISA, diagnostic sensitivity and specificity were 90.48% and 88.71%, respectively. The sensitivity was 88.88% and 100% for serotypes O and A, respectively.
Discussion
Diagnostic assays based on NS proteins have not received much attention in Iran until recently. Early detection of FMD-infected animals has significant importance in sero-surveillance of the disease during the control program. There are several commercial ELISA Kits, most of which are based on 3ABC proteins. However, it has been suggested to use more than one NS antigen to increase diagnostic reliability (34).
In the present study, major epitopic sites of 3AB and 3D NS proteins were considered for recombinant protein production to find whether or not they are enough for FMD detection or DIVA. These epitopes are highly immunogenic and highly conserved in all seven FMDV serotypes (14,27). Anti-3D antibodies are produced as early as 5 days postinfection and remain in sera of infected animals until almost 600 days (22). The 3AB and 3ABC NS kits are frequently used for DIVA in FMD-free countries and also FMD endemic areas (17). In this study, anti-3AB antibody showed higher titer than anti-3D antibody at a similar assay condition. This may suggest that the selected antigenic sequence of 3AB is more immunogenic than the selected sequence of 3D in cattle. In addition, due to the denaturing condition of epitopic 3D protein, lower reactivity in ELISA assay could be expected.
It should be mentioned that shorter proteins with specific B cell epitopes would be better alternatives for whole proteins in detection assays, since they would reduce the chance of nonspecific interactions with antibodies or infections resulted by other Picornaviridae family members (20). Synthetic peptides based on bovine linear B cell epitopes and spanning antigenic sequences of NS proteins have been studied in peptide-ELISA for DIVA and have shown good results in detection of FMD-infected animals, irrespective of virus serotype (23,24).
In addition, carrier animals, generated by vaccination and consecutive infection, can be detected by peptide-based ELISA. Carriers that do not show FMD symptoms are a continuous threat for nonvaccinated animals. Application of an appropriate and sensitive method for detection of these animals is very important for early FMD diagnosis and control (14). Although synthetic peptides are simple to prepare and purify, the synthesis is expensive, and it seems that antigenic recombinant protein production would be a more cost-effective method for preparing antigens for FMD-ELISA Kits.
3ABC polyprotein has been the most reliable NS protein used for commercially available ELISA Kits. However, recombinant production of 3ABC in E. coli has shown expression of a truncated version of 3ABC with a molecular weight similar to 3A protein. This was due to the cleavage of 3ABC into each individual protein, probably by the 3C protein activity, which led to low efficiency of 3ABC expression (28). Moreover, no bovine B cell epitope has been found in 3C (14). Hence, we used E. coli host for expression of truncated epitopic 3AB recombinant protein and to disregard 3C protein.
Previous reports have shown that 3A, 3B, 3AB, and 3ABC proteins are expressed in insoluble state and need to be purified under denaturing condition (28,36). The antigenic 3AB protein produced here was purified completely in soluble and monomeric state. In addition, it contained all bovine DIVA associated B cell epitopes that have been previously reported (14) and so showed a high antibody response in a concentration as low as 0.5 μg/mL and a serum dilution of 1:400. However, it was not the same for the truncated 3D protein. The full 3D protein has been shown in previous studies to be purified mostly in soluble state (19,22,37). Shortening the protein to use only immunogenic amino acids in detection, and increase the antibody response, did not lead to expected results. The truncated 3D protein was purified under denaturing condition from bacterial lysates, and lower solubility may have affected the antibody detection, as higher antigen concentration (2 μg/mL) and lower-diluted sera in comparison with 3AB were needed to reach the detection threshold. It seems that the three-dimensional structure of the shortened 3D protein changed in the way that lower hydrophilic residues were exposed. Although the selected regions of 3D protein were linear epitopes, denaturation of the protein structure may have had an effect on its ability in detecting anti-FMD antibodies.
Different NS proteins have long been studied for FMD disease detection and differentiation. Mackay et al. (18) have declared that detection of antibody response to 3ABC, in addition to one of other NS proteins (L, 2C, 3A, or 3D), is a convincing evidence of previous infection with FMDV, although there are exceptions with multiply-vaccinated animals.
Classification of infected or vaccinated animals simplifies epidemiological studies. A full-length 3D protein has been expressed in insect cells and was used in indirect ELISA to assess the presence of 3D antibodies in herds. This recombinant 3D protein was introduced as a useful tool for screening the animals as an adjunct to other available NSP-based diagnostics for routine sero-surveillance of FMD (16). A 3D-indirect ELISA has been developed by Mahajan et al. (19), which showed lower specificity in uninfected vaccinated animals suggesting against its application in multiply vaccinated herds. However, due to its high sensitivity and longer persistence of anti-3D antibody after infection, it has been suggested for sero-epidemiological studies in unvaccinated regions.
In this study, it was noticeable that epitopic-3AB-ELISA assay showed higher diagnostic sensitivity and specificity than epitopic 3D. Lower specificity of truncated-3D-ELISA was anticipated according to previously published studies on full-length 3D protein ELISAs for multiply-vaccinated animals (19,32). However, lower sensitivity of epitopic 3D-ELISA faints its applicability as a complementary assay to 3AB-ELISA in screening of herds for FMD status. We suggest that by application of our in-house developed 3AB-ELISA assay, we would be able to develop routine sero-surveillance programs in endemic regions of Iran. Both 3AB- and 3D-ELISA assays showed 100% sensitivity for serotype A and lower sensitivity for serotype O. However, due to the fewer numbers of cattle infected with serotype A than with O, this would not be a good measure for comparison of the test efficacy between these serotypes.
Detection of FMD infection at early stages is indispensable for disease management and control, particularly in countries with vaccination strategy. Iran is an endemic region for FMD, and there is a continuous spread of serotypes O, A, and Asia1 in Iran. According to World Organisation for Animal Health (OIE), vaccination alone cannot control the disease, and serological surveys and surveillance systems aimed at the timely detection of FMD are very important for disease monitoring and control. Tests for antibodies to FMD viral NS proteins are available for detection of undisclosed virus circulation (5,12). Purchasing or designing assays for FMD detection is a prerequisite for control programs in Iran. This study showed that our in-house ELISA assay designed based on epitopic regions of 3AB NS protein has the ability to detect FMD virus infection and have the potential to be used for FMD diagnosis and DIVA strategy.
Footnotes
Acknowledgments
The authors would like to show their gratitude to Dr. Ali Mirjalili, Department of Genomics and Genetic Engineering, Razi Vaccine and Serum Research Institute, due to his kind advices in ELISA experiments.
P.M. is from Kharazmi University, and A.Z., M.B., and H.M. are from Razi Vaccine and Serum Research Institute, both where research and education are the primary functions.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was funded by Razi Vaccine and Serum Research Institute, Karaj, Iran (grant number: 2-18-18-021-960362).