
Abstract
Peripheral follicular helper T (pTfh) cells represent specialized CD4+ T cells that help B cells to secrete antibodies. Dengue infection appears to cause immune activation in a wide array of immune cells. Herein, we investigated the signatures of immune activation of circulating Tfh cells and mucosal-associated invariant T (MAIT) cells in adult subjects with confirmed acute clinical dengue virus (DENV) infection by multiparametric flow cytometry. The acute DENV infection induced a significant expansion of highly activated pTfh cells and circulating MAIT cells during acute febrile infection. We found a higher frequency of activated PD-1+ Tfh cells and CD38+ pTfh cells in clinical DENV infection. We also found similar activated and expanding phenotypes of MAIT cells in the patients tested. The total counts of activated pTfh cells and circulating MAIT cells were higher in dengue patients relative to healthy controls. We concluded that pTfh cells and circulating MAIT cells represent activated phenotypes in acute DENV infection.
Introduction
Dengue is a mosquito-borne disease of the tropical and subtropical world and is caused by at least four dengue virus serotypes (DENV1–DENV4) (3) causing a wide spectrum of symptoms ranging from a mild febrile DENV illness to more severe hemorrhagic fever (DHF) and shock syndrome (DSS). Estimates suggest that ∼96 million people are infected across the tropical and subtropical world, and the burden is especially alarming in Asia (3). Severe clinical dengue infection is characterized by coagulopathy, permeability, and leaky vasculature that eventually could progress to deadly hypovolemic shock (7). Dengue patients present with leukopenia and thrombocytopenia (12,30), and dengue hemorrhagic fever can often be fatal (23).
The immunology of DENV infection is highly complicated and the severity of DENV infection is dependent on mechanisms such as antibody-dependent cellular cytotoxicity (2) and cytokine storm (also called as macrophage activation syndrome or hypercytokinemia), resulting from antibody-dependent enhancement (ADE), and hence, cross-reactive antibodies to different serotypes of DENV infection play a critical role in DENV pathogenesis (25). CD4+ T cells remain the key orchestrators of host immune responses, and follicular T helper cells (Tfh) represent one of the important T cell types responsible for antibody-mediated immune responses in the host (26). During a virus infection, dendritic cells (DCs) induce Tfh responses and communicate with Tfh cells via the inducible T cell costimulator (ICOS) to engage the ICOS receptor on Tfh cells. Tfh cells express transcription factor B cell lymphoma-6 (Bcl-6) and programmed death-1 (PD-1). Virus-infected DCs communicate with Tfh cells via ICOS to engage the ICOS receptor on Tfh cells. Bcl-6 induces the expression of CXCR-5 on Tfh cells that interacts with CXCL-13 produced by the stromal cells of B cell follicles in the regional lymph node (17). Subsequently, Tfh cells resident in the T cell zones move to the B cell zones to secrete their signature cytokines, notably IL-21. These cytokines necessitate immunoglobulin (Ig) class switching in B cells (17,26).
Mucosal-associated invariant T (MAIT) cells are a population of innate-like T cells that represent 1–10% of circulating T cells in healthy individuals, and are also found in the gut, lungs, blood, liver, and several mucosal tissues (5,22). MAIT cells express CD8, CD161, and an invariant TCR Vα7.2 and recognize bacterial-derived riboflavin metabolites presented on an MR-1 ligand expressed on antigen-presenting cells (11,32), and can also be activated by IL-12 and IL-18. The role of MAIT cells has been explored in HIV, HBV, and HCV infections where their frequencies are reportedly decreased (1,21,24,31).
Ex vivo analyses of T cell subsets especially peripheral follicular helper T (pTfh) cells and MAIT cells in acute DENV infection have seldom been performed. We used the multicolor flow cytometry approach to investigate the levels of pTfh cells and MAIT cells in acute clinical DENV infection. We report significant activation of pTfh cells during the febrile phase of acute DENV infection. Tfh cell activation is defined by the coexpression of PD-1 and CD38 or high expression of PD-1. We also report the high expression of Ki67 and CD38 on MAIT cells. Our study shows that both pTfh cells and MAIT cells are activated during the febrile phase of acute DENV infection.
Materials and Methods
Ethical approval and consent
The pilot investigation was conducted following approval of the study protocols involving human subjects by the Medical Ethics Committee (MEC) of the Government Theni Medical College and Hospital, Theni, India (Ref. No. 2443). The study was carried out in accordance with the International Conference on Harmonization Guidelines and Declaration of Helsinki. Written informed consent forms were approved by the MEC and signed by the participants.
Clinical assessment of DENV infection
Whole blood samples (5 mL) were collected by venipuncture in K2 blood collection tubes from individuals admitted with dengue fever (n = 6) at the Government Theni Medical College, Theni, during the months of October to December 2019. Patients were diagnosed with DENV infection using an NIV (National Institute of Virology, Pune, India) IgM capture enzyme-linked immunosorbent assay (ELISA) to detect dengue-specific IgM (optical density [OD] >2.1 times P/N ratio, that is, P refers to a known positive control and N the known negative control). DENV-specific IgG was detected using a Panbio Dengue IgG capture ELISA kit (IgG >22 U). The 2011 World Health Organization (WHO) dengue diagnostic criteria were followed to determine clinical DENV severity (23). Based on the criteria, we were able to classify six patients as having dengue fever. Fourteen healthy controls were also recruited in the investigation to study the frequency and phenotypes of pTfh and MAIT cells.
Immunostaining
Two panels were used for immunostaining to evaluate CD4+, CD8+, pTfh cells and MAIT cells for immune activation by multiparametric flow cytometric determination in the study participants. The pTfh panel consisted of APC-conjugated anti-CD3 (clone UCHT1), PerCP-conjugated anti-CD4 (clone OKT4), APC-780-conjugated anti-CD8 (clone RPA-T8) (all from EBioscience), FITC-conjugated anti-CXCR5 (clone J252D4), PE-conjugated anti-PD-1 (clone EH12.2H7), and PE-Cy7-conjugated anti-CD38 (clone HIT2) (all from BioLegend, UK). The MAIT cell panel consisted of the following fluorescent-conjugated antibodies: FITC-conjugated anti-CD3 (clone OKT3), PerCP-Cy5.5-conjugated anti-CD4 (clone OKT4), APC-780-conjugated anti-CD8 (clone RPA-T8), PE-Cy7-conjugated anti-CD161 (clone HP-3G10) (all from EBioscience), APC-conjugated anti-TCR Vα7.2 (clone 3C10) (BioLegend), and PE-conjugated anti-Ki67 (clone 20Raj1) (EBioscience). Immunostaining was performed as per standard staining procedures described previously (21,32). Permeabilization buffer (1 × ) (Cat. No. 00-8333-56; EBioscience) was prepared by mixing one part of 10 × concentrate with nine parts of distilled water.
Multiparametric flow cytometry
Flow cytometry was performed as described previously (32). Briefly, 100 μL of whole blood was taken in an fluorescence activated cell sorter (FACS) tube and vortexed briefly, to which the antibody cocktail was added and incubated in the dark at room temperature (RT) for 30 min. One milliliter of 1 × Fix/Lyse solution (Cat. No. 00-5333-54; EBioscience) was added, vortexed gently, and incubated at RT for 1 h. The stained cells were centrifuged at 500 g for 5 min, and the pellet was washed twice with 1 × wash buffer at 500 g for 5 min. The final pellet was resuspended in 200 μL of FACS buffer. Samples were acquired on a CYTOFLEX −6 color flow cytometer (Beckman Coulter). For immunostaining for MAIT cell proliferation, the centrifuged pellet was resuspended in 100 μL of permeabilization buffer, and 8 μL of PE-tagged Ki67 was added and incubated at RT for 1 h in the dark, to which 2 mL of 1 × permeabilization buffer was added and centrifuged at 500 g for 5 min at RT. The above step was repeated and the pellet was resuspended in 100 μL of FACS buffer before acquiring the cells on a CYTOFLEX −6 color flow cytometer (Beckman Coulter). Flow cytometry data were analyzed using FlowJo version 9.0 (TreeStar, Ashland, OR).
Statistical analyses
Analysis was performed to compare the percentages and mean fluorescence index (MFI) of the proliferation marker (Ki67), immune activation markers, CD38 and PD-1 in CD4+ and CD8+ T cells, MAIT cells, and pTfh biomarkers between DENV-infected individuals and healthy controls. Comparisons were made using a nonparametric Mann–Whitney U test. Two-tailed p < 0.05 was considered statistical significance for all the tests performed, and p-value <0.05, <0.01, <0.001, and <0.0001. All statistical analyses were done using GraphPad Prism software version six (La Jolla, CA).
Results
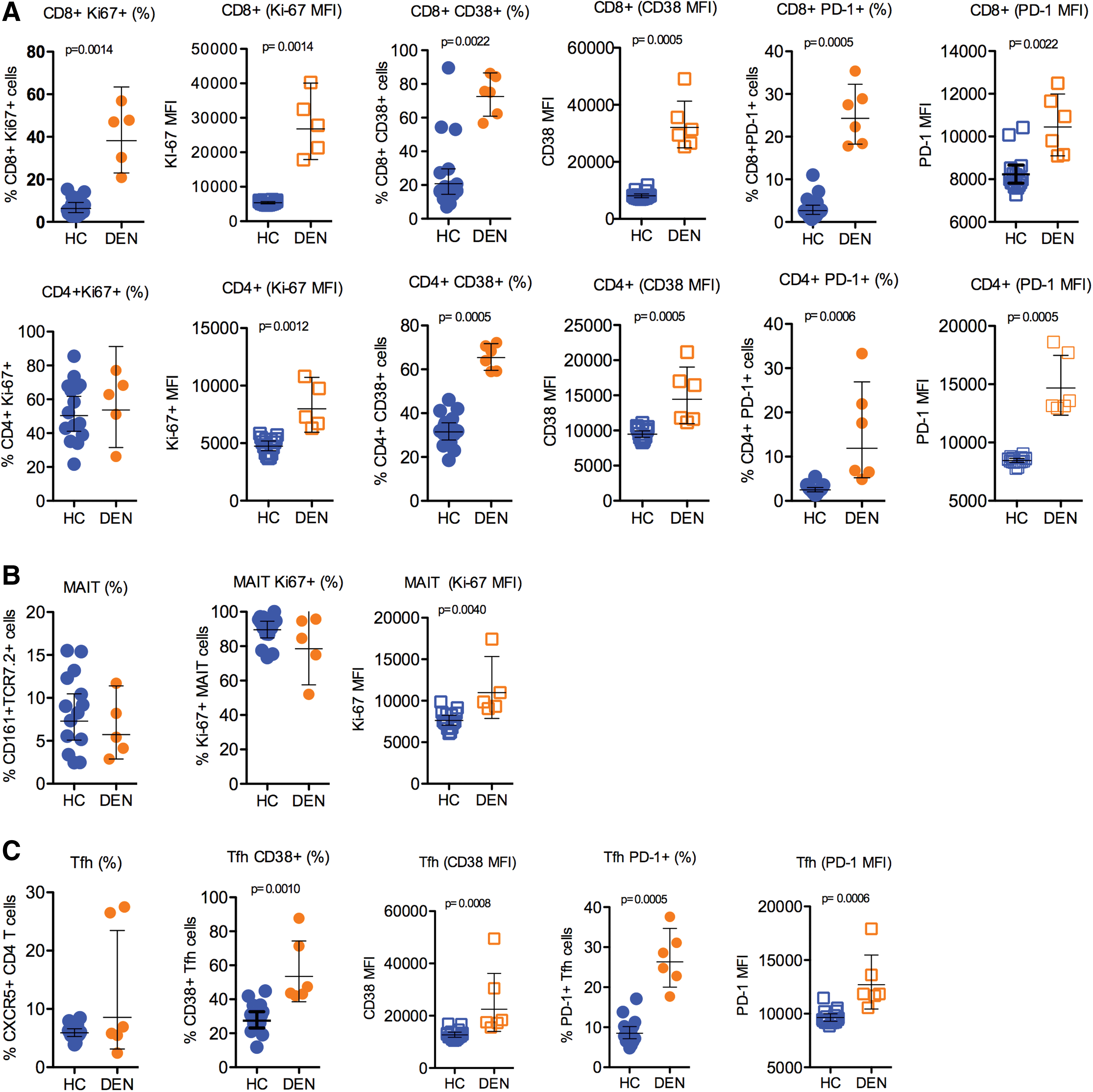
We conducted a study of patients with acute DENV infection that included six patients with confirmed acute DENV infection. Blood was collected at enrollment (typically 3–4 h after admission). Blood specimens were also collected from healthy donors (n = 14). Most acute viral infections induce proliferation and activation of T cells detected by an increase of Ki67 or CD38 markers. Given the speculation that acute DENV infection could impact the proliferation and activation status of T cells, we looked at the level of expression of proliferation and activation markers, Ki67, CD38 and PD-1 on MAIT cells and pTfh cells (CXCR5+CD4+ T cells) (15,17,27). The gating strategies used for studying the frequencies of MAIT cells and pTfh cells are shown in Figure 1. First, we studied the frequency of expression of the proliferation and activation markers Ki-67 and CD38 on both total CD8+ and CD4+ T cells. There was a significant increase in Ki67+, and CD38+ on total CD8+ and CD4+ T cells. Similar to the frequencies, the T cells from acute DENV-infected patients showed significantly elevated expression of Ki67 and CD38 (see corresponding MFI values in Fig. 2A) indicating that the cells were highly activated phenotypes (Fig. 2A). In addition to proliferation and activation markers, we also studied the expression of PD-1; although PD-1 is considered an exhaustion marker during chronic infection, it is regarded as a marker of activation during acute infection (28). There was a significant increase in PD-1+ total CD8+ and CD4+ T cells. In addition, the acute dengue patients had significantly high levels (MFI) of PD-1 on both CD8+ and CD4+ T cells (Fig. 2A). Overall, these data indicate that broad T cell activation, potentially driven by bystander activation and/or homeostatic proliferation in addition to antigen-driven activation of virus-specific CD8+ and CD4+ T cells, occurred.
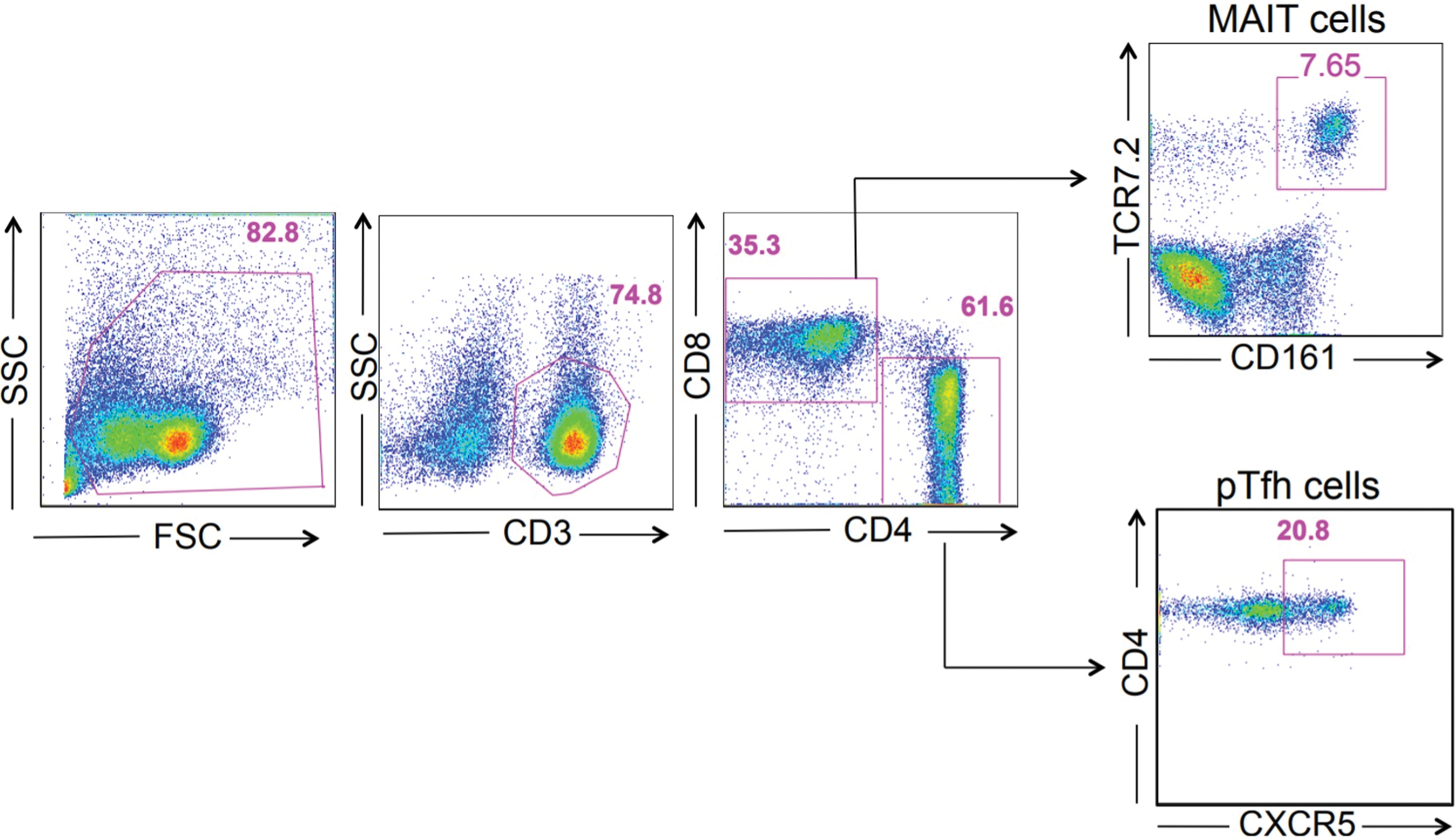
Representative FACS plots showing the gating strategies, distribution of CD3+, CD4+, CD8+ T cells, TCR Vα7.2+ MAIT cells, and CD4+ CXCR5+ pTfh cells in clinical DENV infection. The activation markers (Ki67 and CD38) and acute activation marker (PD-1) were measured from T cell subpopulations of DENV-infected patients. DENV, dengue virus; FACS, fluorescence activated cell sorter; MAIT, mucosal-associated invariant T; PD-1, programmed death-1; pTfh, peripheral follicular helper T. Color images are available online.
Distribution of CD4+ T cells, CD8+ T cells, TCR Vα7.2+ MAIT cells, and CD4+ CXCR5+ pTfh cells in clinical DENV infection.
MAIT cells are a distinctive innate-like T cell subset with antimicrobial activity. These cells are also shown to be activated in acute viral infections (20). MAIT cells are phenotypically identified as T cells coexpressing TCR Vα7.2 and CD161 in humans (1,13,16). Hence, we also studied the levels of circulating MAIT cells (CD3+CD8+161+TCR Vα7.2+ cells), and our results indicated that the frequency of MAIT cells was not different between the healthy controls and DENV-infected patients, although their proliferative status in terms of Ki-67 expression was higher (Fig. 2B).
Finally, we examined CD4+ T cells; given the potential role of antibodies in the response to DENV infection, we analyzed circulating Tfh cells. These cells are characterized by the expression of the follicular homing marker CXCR5 in the blood (14,27). In addition, we also studied the activated pTfh cells CXCR5+CD4+ T cells expressing CD38 and PD-1; the latter could be more reflective of recent antigen encounter and emigration from the germinal center (10). These analyses revealed that there was no difference in the frequency of CXCR5+ CD4+ T cells, although there was a significant difference in the frequency of both CD38+ and PD-1+CXCR5+CD4+ T cells, both in cell frequency and MFI significantly (CD38+CXCR5+CD4 T cells), p < 0.001 and (PD-1+CXCR5+CD4 T cells) p < 0.0001, respectively (Fig. 2C). Altogether, these data highlight hyper T cell activation in febrile DENV patients/DENV disease similar to what has been observed in other acute infections.
Discussion
Improved understanding of the role of T-cell activation and proliferation is urgently required to develop therapeutic modalities against acute DENV infections. Herein, we report the first analysis of pTfh cell and MAIT cell activation during acute DENV infection from PBMCs of adult subjects in a cross-sectional investigation. Previous studies have supported the current findings of activated CD8+ T cells (4,6,8,22,24,25) and also pTfh cells (9). One study assessed the expression of CD38 to mark T cell activation during the febrile stages of DENV infection (9). Advancing our understanding on the quality of immune cells in acute DENV infections is important for detecting key targets for potential therapeutic applications.
We found that the expansion and activation of CD8+ T cells were significantly high compared with CD4+ T cells. Of the CD8+ T cells, we were particularly interested in CD161+ TCR Vα7.2+ MAIT cells, and among the CD4+ T cell phenotypes, CXCR5+ Tfh cells (15). The phenotypic analysis was carried out on both pTfh and MAIT cells during the febrile phase of DENV infection, which is critical to determine the importance of systemic inflammation in necessitating immune cell activation. MAIT cells represented an activated phenotype revealing a significant difference (p < 0.01) in response to Ki67, the proliferation marker. Others have reported that MAIT cells had elevated coexpression of CD38, suggesting that acute DENV infection promotes immune activation (18). pTfh cells also showed an activated phenotype given that the cells showed significantly higher expression of CD38 and PD-1 molecules. Others have shown that IL-21, a signature cytokine of Tfh cells, was high in sera of patients during the early convalescent phase of DENV infection (29), suggesting that Tfh cells likely have a protective role in DENV infection by helping B cells to produce antibodies. Another recent investigation evaluated the levels of pTfh cells in acute DENV infection during the different phases of infection and found that high frequencies of activated pTfh cells occurred during the critical phase of dengue illness. They also reported that the number of pTfh cells was higher in secondary relative to primary DENV infection [31]. Reports on evaluation of pTfh cells in acute DENV infections are seemingly less, and the current study was successful in assessing the activation of pTfh cells as well as MAIT cells in DENV infection. Our study is in line with previously published studies on the activation of Tfh cells in DEVN infection (9, 33). Interestingly, the CD38+ Tfh cells are shown to correlate with functional antibody responses during influenza vaccination (10). In addition, a recent study has also shown that the CD38+ Tfh cells found in the blood can also occur in the lymph and can have characteristics similar to germinal center (GC) Tfh cells, suggesting a direct relationship between CD38+ pTfh and events in lymphoid tissue (26).
Although certain advances have been achieved from the current work, the study indeed has its own limitations; the foremost being the poor DENV sample numbers, which is attributed to the poor number of admitted during the study tenure. The second limitation is that the study failed to explore the roles of other immune activation molecules such as CD69 and HLA-DR, which might have added value to the concepts discussed in terms of quality of immune cells studied. The third limitation is the lack of functional studies on the cells investigated in acute DENV infection. The other major limitation is that the study unlike others (8,9) has not attempted to longitudinally investigate the kinetics of activated cells, especially B cells as immune activation is regarded often as a dynamic process that is known to gradually weaken the host's systemic immune responses, particularly involving antibodies (8,9). It is also believed that the pTfh and MAIT cell dynamics might change over the entire course of the illness, and therefore, a longitudinal study would have been preferred to a cross-sectional investigation. The quantification of IL-21 would also be beneficial to correlate the functional aspects of pTfh cells in clinical dengue disease.
Together, we concluded that pTfh cells and MAIT cells are significantly activated during the febrile stage of clinical dengue infection. Our findings suggest that activation of pTfh cells could serve as a novel marker for critical DENV illness, also supported by findings from other groups (9, 33). Since pTfh cells display highly activated and proliferative phenotypes in febrile DENV infection, further work is required to understand the dynamics of binding and neutralizing capacity of antibodies during the peak and memory phases of DENV infection. Thus, our work helps in understanding the simultaneous phenotypic analysis and frequency of distribution of circulating MAIT cells and pTfh cells during the febrile phase of dengue disease.
Footnotes
Authors' Contributions
E.M.S., A.M., and V.V. designed the study. S.U.P., E.M.S., and A.M. carried out the experiments. S.S., M.L., H.Y.T., and Y.K.Y contributed the reagents and materials. E.M.S., M.L., and S.U.P. wrote the article. All authors have read and agreed to the published version of the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
E.M.S. is supported by the Department of Science and Technology-Science and Engineering Research Board, Government of India (Grant Number CRG/2019/006096). M.L. is supported by: AI52731, The Swedish Research Council, The Swedish Physicians Against AIDS Research Foundation, The Swedish International Development Cooperation Agency, SIDA SARC, VINNMER for Vinnova, Linköping University Hospital Research Fund, CALF, and The Swedish Society of Medicine. Y.K.Y. is supported by a grant from Xiamen University Malaysia Research Funding (XMUMRF), XMUMRF/2018-C2/ILAB/0001. H.Y.T. is supported by a grant from Xiamen University Malaysia Research Funding (XMUMRF), XMUMRF/2020-C5/ITCM/0003. A.M. is supported by the Department of Health Research, Ministry of Health and Family Welfare (Grant No. VIR/66/2013/ECD-I) and (R.12020/04/2018-HR), Government of India. V.V. was supported by Emory University CFAR grant P30 AI050409 and NCRR/NIH base grants P30 RR00165, P51OD011132 (to Y.N.P.R.C.).