
Abstract
Avian influenza viruses (AIVs) infect a wide range of hosts, including humans and many avian species. Efforts have been made to control this pathogen in chickens using vaccination programs, but that has been met with varying degrees of success. Therefore, identification of more efficacious vaccination strategies is warranted. This study was undertaken to investigate the potential effects of probiotics on the immunogenicity of a beta-propiolactone-whole inactivated virus (WIV) vaccine of H9N2 subtype adjuvanted with the Toll-like receptor-21 ligand, CpG oligodeoxynucleotides 2007 (CpG). Eighty-four 1-day-old White Leghorn layers were allocated into six groups. Two out of six groups received a mixture of probiotic Lactobacillus spp. (PROB) biweekly from days 1 35 of age. Chickens were intramuscularly vaccinated with WIV either alone or adjuvanted with AddaVax™ (WIV+Add) or CpG (WIV+CpG), and one group received saline (phosphate-buffered saline). Primary and secondary vaccinations occurred at days 14 and 28 of age, respectively. The results revealed that the group that received probiotics and was vaccinated with CpG-adjuvanted WIV H9N2 vaccine had higher hemagglutination inhibition titers than the other treatment groups at days 14 and 21 postprimary vaccination. Probiotics did not induce higher IgM or IgY titers in chickens receiving the WIV vaccine only. Concerning their effect on cell-mediated immune responses, probiotics enhanced interferon-gamma (IFN-γ) gene expression and significantly increased secretion of IFN-γ protein by splenocytes in chickens vaccinated with CpG-adjuvanted WIV H9N2. Together, these findings suggest the use of probiotics to enhance the immunogenicity of CpG-adjuvanted WIV H9N2 vaccines. Additional studies are required to better understand the specific interactions between probiotics and the gut microbiota and different types of cells of the gastrointestinal tract to decipher the underlying mechanisms of how probiotics modulate immune responses to vaccines.
Introduction
Avian influenza virus (AIV) H9N2 is a subtype of the influenza A virus species and belongs to the family Orthomyxoviridae. It is the most widespread AIV subtype in poultry worldwide (18). Development of new vaccines and enhancement of vaccine-induced immunity against H9N2 AIV may play an important role in controlling AIV infection in chickens. Although mild clinical signs are observed during infection with H9N2 AIV in chickens, a deterioration of eggshell quality and a gradual decrease in egg production are, indeed, among the most common signs of infection (22). Coinfections with other respiratory pathogens can aggravate the clinical signs and economic losses caused by this virus (14). In addition, recent evidence indicates that H9N2 AIV could have a key role in a future influenza pandemic due to the fact that it provides internal genes to other AIV, including a novel H7N9 virus (15). Vaccination against H9N2 AIV has been used as a method for controlling infections in birds, and vaccination programs have been adopted in endemically affected countries (11,21). However, H9N2 viruses are somewhat neglected because vaccines efficacy is not assessed regularly against antigenically drifted viruses resulting in H9N2 to continue causing disease in vaccinated poultry, whereas it is better to control H9N2 viruses to prevent emergence of new zoonotic viruses (21). Therefore, due to limitations of existing vaccines, other complementary strategies should also be envisaged.
Various natural or synthetic adjuvants have been used to enhance the immunogenicity of inactivated virus vaccines. Toll-like receptor (TLR) ligands, which are a group of structurally conserved molecules present in or on microbes and known as pathogen-associated molecular patterns, have shown potential to serve as vaccine adjuvants in a variety of species, including chickens (28). TLRs are a class of pattern recognition receptors expressed by various leukocytes and are located on the cell surface or in the membranes of endosomes. Ligation of TLRs leads to intracellular signaling cascades resulting in increased expression and production of cytokines and costimulatory molecules essential for initiating innate responses and subsequent adaptive immune responses (33).
TLR ligands were used successfully as vaccine adjuvants resulting in enhanced immunogenicity of vaccines and a subsequent reduction of virus shedding in chickens (17,25 –27,29). Recent studies have demonstrated that CpG oligodeoxynucleotides (ODN) 2007 synthetic oligonucleotide, which is recognized by TLR21 in chickens, can be used as an adjuvant and showed the ability to enhance the efficacy of various vaccines in chickens (3,27,28). In addition, AddaVax™, a squalene-based oil in water emulsion adjuvant, has a formulation similar to MF59®. MF59 has been licensed in Europe as an adjuvant in flu vaccines. These adjuvants can elicit both cellular and humoral immune responses (8,22).
Probiotics have emerged as a useful tool to enhance immune responses. Oral administration of probiotics in chickens has been shown to increase the elicitation of antibody responses (2,5,14,15). In addition, administration of probiotic lactobacilli has modulatory effects enhancing the expression of immunological genes in vivo and in vitro. In a study in which gut microbiota was disrupted, immunity to infection of chickens with AIV was negatively impacted, resulting in higher viral shedding (38). Moreover, administration of probiotic lactobacilli to antibiotic-treated chickens restored type I interferon (IFN) and interleukin (IL)-22 mRNA levels in the gastrointestinal and respiratory tracts compared with those observed in untreated chickens (40). Using the same gut microbiota-depletion model with antibiotics, oral administration of probiotic lactobacilli enhanced antibody-mediated responses to a H9N2 AIV inactivated vaccine in chickens (39).
Studying the possible effects of probiotics on the immunogenicity of a H9N2 AIV vaccine when combined with TLR ligands as adjuvants may provide a platform for the development of more effective immunization strategies. In view of this, this study was undertaken to characterize the immune responses in chickens treated with probiotic lactobacilli and vaccinated with a beta-propiolactone (BPL)-inactivated H9N2 whole inactivated virus (WIV) vaccine adjuvanted with CpG oligodeoxynucleotides 2007 (CpG ODN 2007).
Materials and Methods
Chickens
A total of 84 one-day-old specific pathogen-free (SPF) White Leghorn layers were purchased from the Canadian Food Inspection Agency (CFIA; Ottawa, Canada) and were housed in the isolation facility at the Ontario Veterinary College at the University of Guelph. All experimental procedures were approved by the University of Guelph Animal Care Committee and conducted according to specifications of the Canadian Council on Animal Care.
Experimental design
Chickens were divided into six groups: group 1 was a negative control group receiving phosphate-buffered saline (PBS) only; group 2 received inactivated H9N2 WIV vaccine alone; group 3 received vaccine adjuvanted with AddaVax as a positive control adjuvant (WIV+Adda); group 4 was treated with probiotics (PROB) and then received the vaccine (PROB+WIV); group 5 received vaccine with CpG as an adjuvant (WIV+CpG); and group 6 was treated with probiotics and received the vaccine with CpG as an adjuvant (PROB+WIV+CpG). Chickens treated with probiotics received 108 colony-forming units (CFUs)/mL lactobacilli cocktail by oral gavage to the crop using a 1 mL syringe on day (D) 1 posthatch (ph) and treatment was repeated twice a week until D35 ph. Vaccinated chickens received 15 μg/dose of the whole inactivated AIV adjuvanted with AddaVax (half total volume of vaccine dose) or CpG (2 μg/dose) intramuscularly at D14 and D28 ph. Cell-mediated immune responses in five chickens per group were measured at D5 postsecondary vaccination (psv). Sera were collected from nine chickens per group weekly, starting from D7 postprimary vaccination (ppv) (Fig. 1).
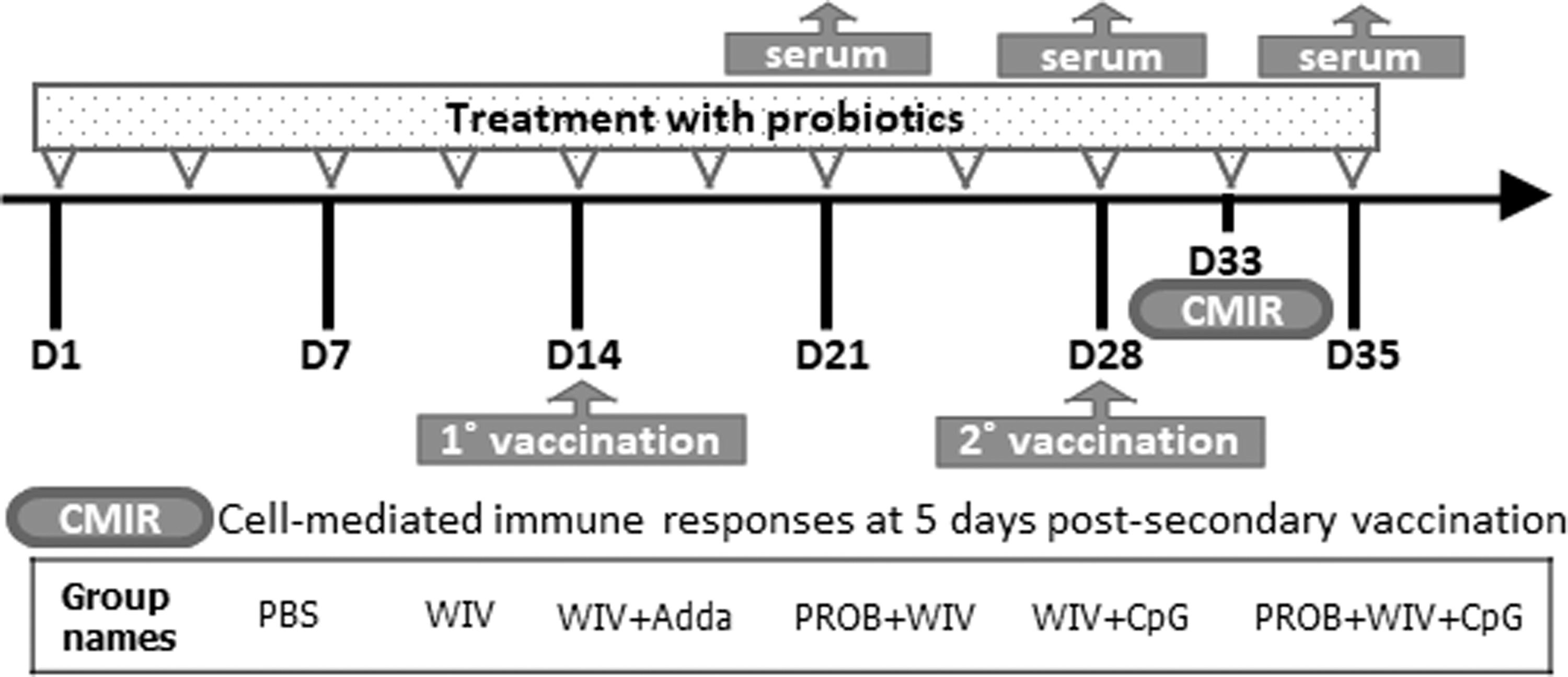
Experimental outline and groups. Chickens (n = 14/group) were administered a cocktail of probiotics twice weekly for 35 days and vaccinated when 14 (1°, primary) and 28 (2°, secondary) days old. Serum samples (n = 9) were collected on days 7, 14, and 21 postprimary vaccination for serology assays. Spleens (n = 5) were collected to assess cell-mediated immune responses at day 5 postsecondary vaccination. PBS, phosphate-buffered saline; WIV, 15 μg whole inactivated H9N2 virus/vaccine dose; Adda, half the vaccine dose of AddaVaxTM; PROB, 108 colony forming units/mL of probiotic lactobacilli; CpG, 2 μg/vaccine dose of CpG oligodeoxynucleotides 2007.
Preparation of probiotic bacteria
Lactobacilli strains were used in this study (Lactobacillus salivarius, L. johnsonii, L. reuteri, L. crispatus, and L. gasseri) were isolated from intestinal contents of broilers by Brisbin et al. (5). Lactobacilli cultures were prepared and administered to chickens, as previously described (40). In brief, different lactobacilli cultures were prepared separately from one another anaerobically in De Man, Rogosa, and Sharpe medium overnight at 37°C and subcultured twice before harvesting and washing with PBS. Optical density (OD) of each culture was measured, and 108 CFU/mL of each culture were mixed in equal proportions up to a total of 1 mL per bird.
Virus propagation and vaccine preparation
The virus used in this study was A/TK/IT/13VIR1864-45/2013 (H9N2), and it was kindly provided by the Istituto Zooprofilattico Sperimentale delle Venezie (IZSVe), Italy. In brief, the virus was inoculated into 10-day-old SPF embryonated-chicken eggs followed by incubation for 72 h at 37°C. Embryos were monitored every 24 h, and dead embryos were discarded until 72 h of incubation. Eggs were then refrigerated at 4°C overnight followed by collection of allantoic fluid, which was then centrifuged at 400 g for 5 min at 4°C and stored at −80°C until further use. The propagated virus titer was determined with the 50% tissue culture infectious dose (TCID50) method.
Inactivation of the vaccine preparation was performed using BPL, as described previously (3). In brief, one part of 0.5 M disodium phosphate was added to 38 parts of virus-containing allantoic fluid followed by dropwise addition of one part 2% BPL solution with continued stirring and incubation for 30 min on ice. The solution was then placed in a 37°C water bath for 2 h with 15 min mixing intervals. The pH of the mixture was adjusted to 7.3–7.4 using 7% sodium bicarbonate solution and stored at 4°C until confirmation of inactivation using both embryonated eggs and Madin–Darby canine kidney (MDCK) cells. Total protein concentration was determined for the inactivated virus using a Pierce bicinchoninic acid protein assay kit (Thermo Scientific, Rockford, IL) following the manufacturer's recommendations. Chickens were vaccinated intramuscularly with 15 μg of the WIV.
TLR ligands
The synthetic class B CpG ODN 2007 (5′-TCGTCGTTGTCGTTTTGTCGTT-3′) (a TLR21 agonist, Sigma) with a phosphorothioate backbone was used in this study at 2 μg per bird. This ligand was chosen for its ability to stimulate cytokine expression in chicken mononuclear cells and to enhance the immunogenicity of a H9N2 AIV vaccine in chickens (3,32,33).
Serology
Hemagglutination inhibition assay
A twofold serial dilution was performed with 50 μL serum samples followed by 30 min of incubation with 50 μL (8 hemagglutinin units) of H9N2 virus at room temperature in 96-well V-bottom plates (Corning Inc., Corning, NY). Chicken red blood cells (RBCs) at a concentration of 0.5% were then added, and plates were further incubated for 30 min. Hemagglutination inhibition (HI) titers were determined as the reciprocal of the highest sample dilution resulting in complete HI of the chicken RBCs.
Virus neutralization assay
Virus neutralization (VN) assays were performed using serum samples from D7 psv, as described previously (27,39), with some modifications according to the microneutralization assay protocol of the World Health Organization (35). In brief, sera were first treated with receptor destroying enzyme (RDE) at a ratio of 3:1 of RDE to serum for 12–18 h at 37°C. The RDE was then inactivated at 56°C for 30 min. Twofold serial dilutions of sera (starting dilution, 1:10) were made in 96-well flat-bottom plates using Dulbecco's modified Eagle's medium containing 50 μg/mL of gentamicin, 100 U/mL of penicillin, 100 μg/mL of streptomycin, 0.2% bovine serum albumin, 25 mM (4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid), and 2 μg/mL of L-1-tosylamido-2-phenylethyl chloromethyl ketone (TPCK)-treated trypsin. Serially diluted sera were then incubated with an equal volume (50 μL) of diluted virus containing 50 TCID50 of H9N2 virus for 1 h at 37°C, 5% CO2. One hundred microliter of 70–95% confluent MDCK cells at 2.5 × 105 cells/mL were added to all wells. Plates were then cultured for 72 h at 37°C, 5% CO2. Each plate included the following controls: serum from unvaccinated chickens as a negative control, positive control with the H9N2 virus, and a cell-only control. The reciprocal of the highest serum dilution to inhibit cytopathic effect was used to calculate the VN titer and expressed as log2.
Measurement of anti-H9N2 IgY and IgM by enzyme-linked immunosorbent assays
Virus-specific IgY and IgM titers in serum were determined by indirect enzyme-linked immunosorbent assays (ELISAs) following methods described previously with minor modifications (39). In brief, 96-well ELISA plates (Nunc MaxiSorp™, Thermo Fisher Scientific Inc., Mississauga, ON) were coated overnight at 4°C with whole heat-inactivated H9N2 AIV using an optimally determined concentration of 800 ng/100 μL of carbonate-bicarbonate buffer, pH 9.6. After washing the plates three times with 300 μL of PBS containing 0.05% Tween 20 (PBS-T), 100 μL of blocking solution (PBS-T containing 0.25% fish gelatin [Sigma-Aldrich]) were added, and the plates were incubated for 1 h at room temperature. After removing the blocking solution, 100 μL of chicken serum (diluted in PBS containing 1.5% Tween-20 and 0.29 M NaCl) were added to the wells. The plates were incubated for 1 h at room temperature. After washing three times, 100 μL of horseradish peroxidase-conjugated goat antichicken IgY or IgM (1:5,000) were added into each well, followed by incubation of the plates for 1 h at room temperature. Subsequently, plates were washed three times. The horseradish peroxidase substrate (ABTS peroxidase substrate system, Kirkegaard and Perry Laboratories Gaithersburg, MD) was added, and plates were incubated in the dark at room temperature for 20 min. Colour development was stopped with 1% sodium dodecyl sulfate, and absorbance at 405 nm was determined using a plate reader (Bio-Tek Instruments, Winooski, VT). Relative antibody titers were calculated in relation to a serially diluted high titer serum sample that was used on every plate using methods described previously (3,25).
Antibody avidity
Serum IgY antibody avidity indices were determined through resistance to 8 M urea, as described previously (18,28). Urea dislodges protein–protein bonds in low-affinity antibodies, whereas high-affinity antibodies remain bound to the antigen-coated plates. The antibodies with epitope binding sites of greater affinity show a higher avidity index. To measure IgY antibody avidity indices, the ELISA conditions described earlier were used with modifications: (1) twofold serial dilutions were used, (2) before the addition of 100 μL of goat antichicken IgY (Fc-specific) antibody conjugated to horseradish peroxidase, the plates were washed with 100 μL/well of either PBS-Tween or 8 M urea in PBS-Tween and incubated at room temperature for 5 min, followed by washing as described earlier. The avidity index was calculated as the average OD405 of urea-treated wells divided by the average OD405 of PBS-Tween-treated wells × 100. Sera with index values of >50% were designated as being of high avidity. Those with index values of 30–50% were designated as having intermediate avidity, and those with index values of <30% were designated as being of low avidity (28).
Cell-mediated immune responses
At 5 days psv, splenic mononuclear cells were prepared from unpooled five chickens per treatment, as described previously (5). Cells were seeded in 48-well flat-bottom plates at a density of 5 × 106 cells/well in 500 μL of RPMI-1640 (Invitrogen, Burlington, ON) supplemented with 10% fetal bovine serum, 200 U/mL of penicillin, 80 μg/mL of streptomycin, and 50 μg/mL of gentamicin and cells were then incubated at 41°C and 5% CO2. Mononuclear cells were then stimulated with 1 μg/mL of BPL-inactivated H9N2 AIV. At 12 and 24 h poststimulation, cells were collected for extraction of RNA. At 48 and 72 h poststimulation, supernatants were collected to assess IFN-γ protein levels using a chicken IFN-γ sandwich ELISA kit as per the manufacturer's recommendations (Invitrogen™).
RNA extraction, cDNA synthesis, quantitative real-time polymerase chain reaction, and IFN-γ gene expression analysis
Extraction of RNA and cDNA synthesis were performed as previously described (34). Quantitative real-time polymerase chain reaction using SYBR Green was performed on diluted cDNA using a LightCycler 480 II (Roche Diagnostics GmbH, Mannheim, Germany) as previously described (34). Each reaction started with a preincubation at 95°C for 10 min, followed by 45 cycles of denaturation (95°C for 10 sec), annealing (60°C for 5 sec), and extension (72°C for 10 sec). Subsequent melting curve analysis was performed by heating to 95°C for 10 sec, cooling to 65°C for 1 min, and heating to 97°C. Primers were synthesized by Sigma Aldrich (Table 1). The IFN-γ gene expression was calculated relative to the housekeeping gene β-actin using software on the LightCycler® 480 II (Roche Diagnostics GmbH), as described previously (4). Data represent the mean of five biological replicates (chickens) ± standard error.
Primer Sequences Used in Quantitative Real-Time Polymerase Chain Reaction
IFN-γ, interferon-gamma.
Statistical analyses
Analysis of multiple treatments for all data (including HI titers, VN, IgY, and IgM titers in serum, antibody avidity, relative expression of IFN-γ mRNAs, and concentration of IFN-γ protein) were performed with one-way analysis of variance using SAS® software (SAS Institute, Inc., Cary, NC). Means were considered significantly different when p < 0.05. Results for multiple comparisons using Duncan's multiple range test were considered significant at p < 0.05.
Results
HI and virus neutralization antibody titers
The negative control group (PBS) showed no detectable agglutination inhibition at any time points. At D7 ppv, there were no significant differences in HI titers among all groups. At D14 ppv, the PROB+WIV+CpG group showed a significantly higher HI titer compared with WIV only and PROB+WIV groups (p < 0.05). Treatment with PROB+WIV+CpG showed significantly higher HI titer than all other groups (p < 0.05) at D21 ppv (Fig. 2).
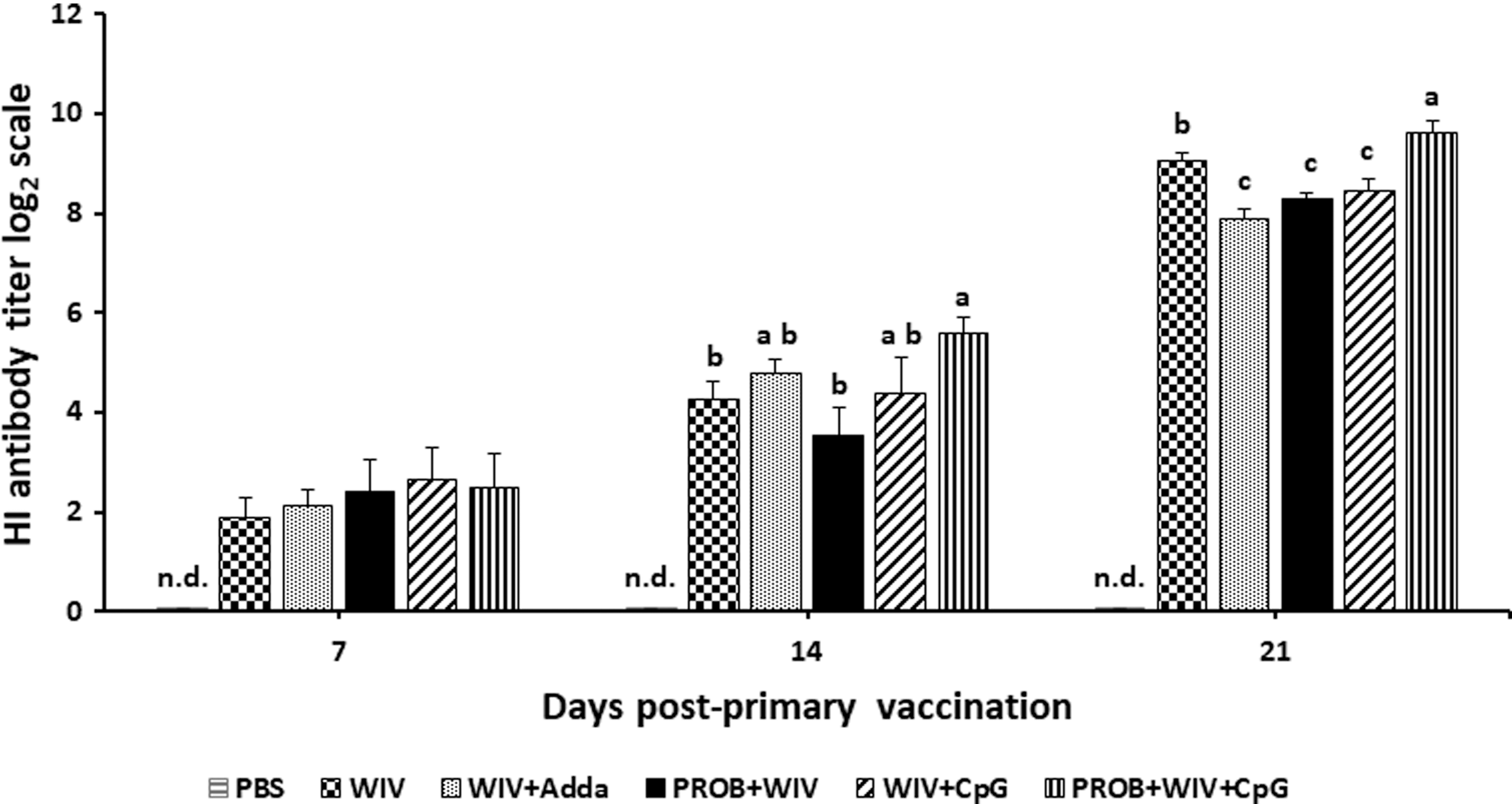
HI antibody titers. Average serum HI antibody titers in chickens from various treatments (n = 9/group) are shown at days 7, 14, and 21 postprimary vaccination. Error bars represent standard errors of the means. Data were analyzed in SAS® software using Duncan's multiple range test. Bars with different letters at a time point differ significantly from each other (p < 0.05). WIV vaccine (15 μg); Adda, AddaVax; PROB (108 CFU/mL); CpG, CpG ODN 2007 (2 μg). HI, hemagglutination inhibition; n.d., not detected.
Virus neutralization assay measured at D7 psv (D21 ppv) showed higher neutralizing antibody titers in sera of chickens vaccinated with the CpG-adjuvanted WIV vaccine, regardless of whether or not they were pretreated with probiotic lactobacilli, but the difference was not statistically significant when compared with other groups (Fig. 3).
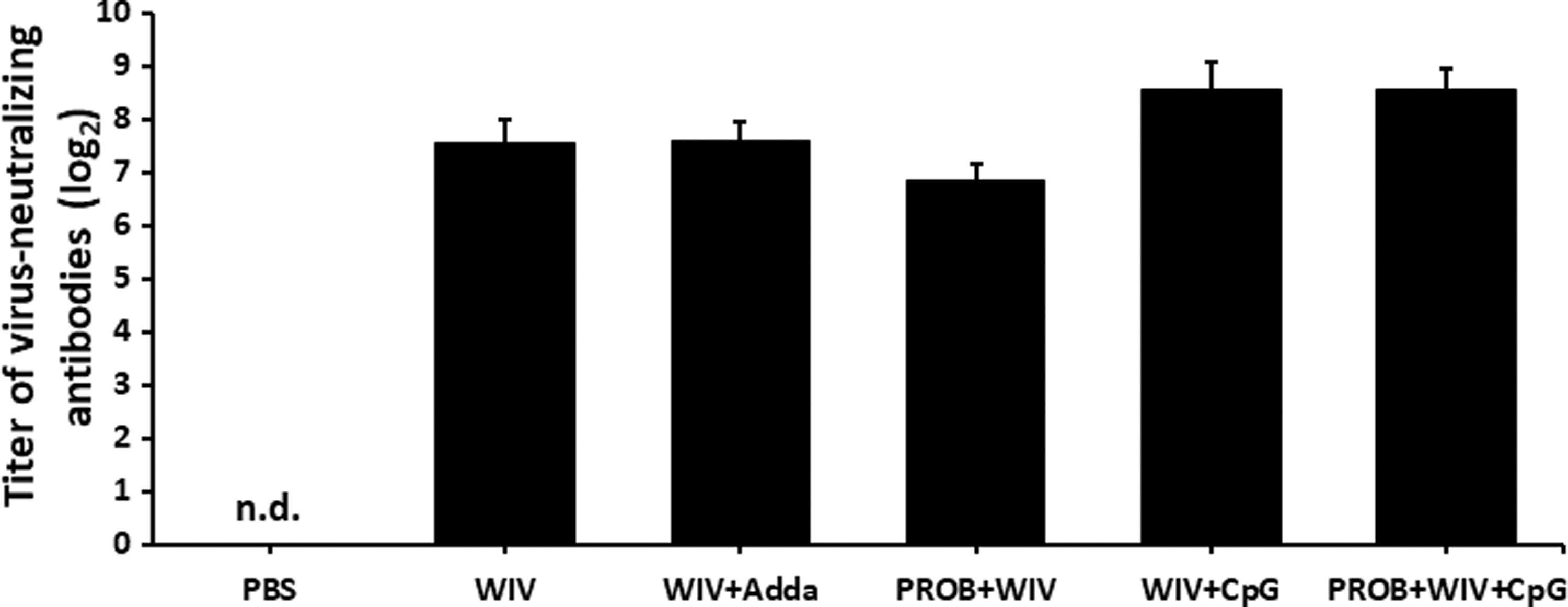
Virus-neutralizing antibody titers. Virus-neutralizing antibody responses are shown in chickens (n = 9/group) on day 7 psv. Error bars represent standard errors of the means. WIV vaccine (15 μg); Adda, AddaVax; PROB (108 CFU/mL); CpG, CpG ODN 2007 (2 μg). n.d., not detected.
H9N2 virus-specific IgM and IgY responses and IgY antibody avidity
At all time points, all treatment groups had significantly higher IgM titers than the negative control group that received only PBS. At D7 ppv, significantly higher IgM titers were observed in WIV+Adda and WIV+CpG groups than in the other groups except for the PROB+WIV+CpG (p < 0.05). No significant differences were observed when comparing these two groups with the PROB+WIV+CpG group. The PROB+WIV+CpG group had significantly higher IgM titers than the PROB+WIV group but not statistically different from the WIV only group. The WIV group had higher, but nonsignificant, IgM titers compared with the PROB+WIV group. At D14 ppv, the WIV+Adda group showed a significantly higher IgM titer compared with the WIV and PROB+WIV groups. At D21 ppv, WIV+CpG and PROB+WIV+CpG groups showed significantly higher IgM titers among all groups (Fig. 4A).

Serum IgM
Significantly higher IgY titers were observed in all groups than the PBS group at all time points (p < 0.05). At D7 ppv, no significant differences were observed among other groups. At D14 ppv, WIV+Adda, WIV+CpG, and PROB+WIV+CpG groups were significantly different from WIV and PROB+WIV groups, which did not significantly differ from each other. At D21 ppv, WIV+CpG and PROB+WIV+CpG groups showed significantly higher IgY titers compared with all other groups. WIV+Adda, WIV, and PROB+WIV groups were not different from each other at D21 ppv (Fig. 4B).
At D7 psv (D21 ppv), high avidity IgY antibodies (>50%) were generated by chickens in all study groups except for the PBS group. The serum IgY antibody avidity index for the PROB+WIV group was 68.1%. Although this avidity index was not statistically different from other groups, it was the highest antibody avidity index. The lowest IgY antibody avidity index was observed in the PROB+WIV+CpG group (55.5%), whereas moderate IgY antibody avidity was observed in the other groups: WIV+Adda (59.6%), WIV+CpG groups (59.8%), and WIV (62.4%) (Fig. 5).
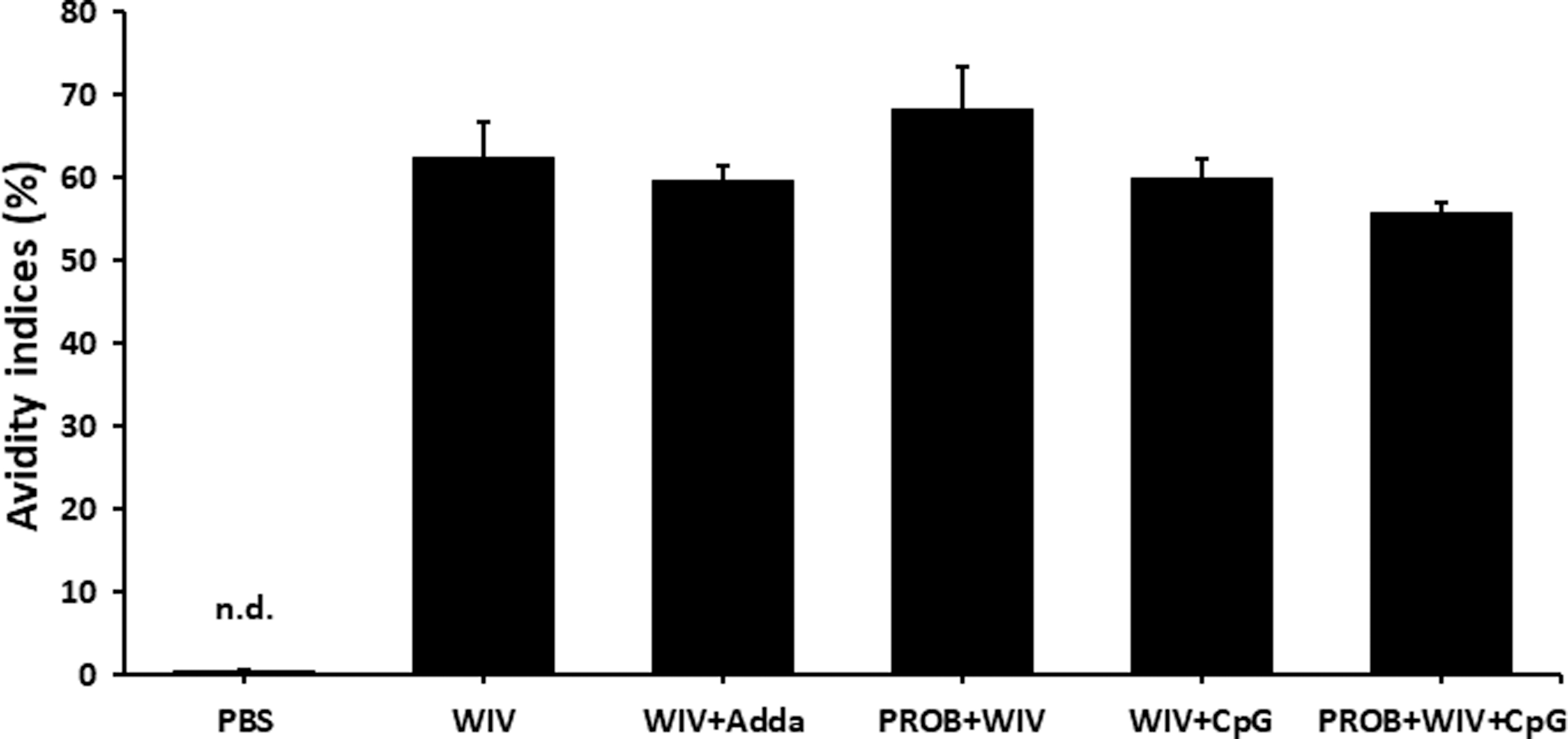
Serum IgY antibody avidity indices. Serum IgY antibody avidity indices of sera from chickens (n = 9) at day 7 psv were determined through resistance to 8 M urea. Error bars represent standard errors of the means. The avidity index was calculated as the average OD405 of urea-treated wells divided by the average OD405 of PBS-Tween-treated wells × 100. An avidity index of >50% represents antibodies of high avidity; values between 30% and 50% represent those of intermediate avidity, and values of <30% represent low avidity antibodies. WIV vaccine (15 μg); Adda, AddaVax; PROB (108 CFU/mL); CpG, CpG ODN 2007 (2 μg). OD, optical density. n.d., not detected.
Cell-mediated immune responses
Chickens were euthanized 5 days psv, and spleens were harvested to analyze cell-mediated immune responses. Splenocytes were stimulated with the vaccine antigen (whole inactivated H9N2 AIV). Analysis of IFN-γ expression showed upregulation of IFN-γ mRNAs in the PROB+WIV+CpG group at 12 and 24 h poststimulation, but expression was not significantly different among the groups at either time point (Fig. 6A). Analysis of IFN-γ protein production using ELISAs reflected the initial observations of IFN-γ gene expression, but the difference was statistically different. Splenic mononuclear cells from the PROB+WIV+CpG group produced significantly higher concentrations of the IFN-γ protein at 48 and 72 h poststimulation compared with all other groups (Fig. 6B).
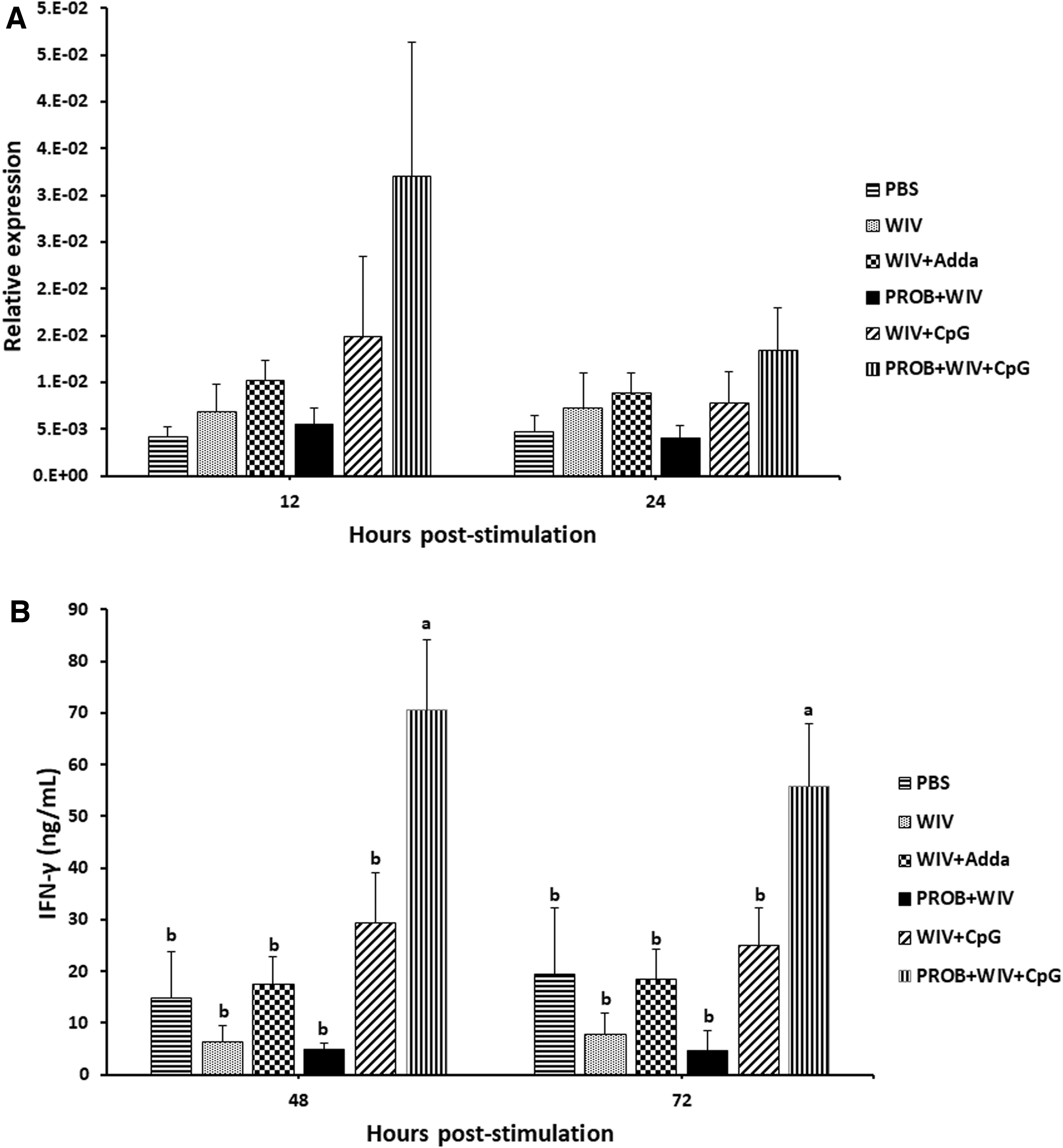
IFN-γ production by splenocytes. Two groups of chickens were administered with probiotic lactobacilli cocktail by oral gavage twice weekly for 35 days. Except for PBS group, all chickens were intramuscularly vaccinated with WIV H9N2 vaccine with different formulations of adjuvants for both vaccinations. Primary and secondary vaccinations were administered at days 14 and 28 ph, respectively. Splenocytes from immunized chickens (n = 5) were restimulated ex vivo with whole inactivated influenza virus subtype H9N2 5 days psv.
Discussion
The subtype H9N2 AIV is the most prevalent low pathogenic AIV in poultry. Aside from its economic impact on the poultry industry, H9N2 AIV poses a threat to public health. It has shown the capability to cross barriers to infect humans and donate its internal genes to emerging zoonotic influenza viruses (1,12,23). Vaccination can be an effective method to control influenza in chickens. However, it is also important to consider more strategies for the enhancement of vaccine efficacy. Indeed, various adjuvants have been investigated to enhance AIV vaccines. It has recently been shown that TLR ligands can serve as AIV vaccine adjuvants with demonstrated ability to augment vaccine immunogenicity (3,27). Furthermore, the utilization of probiotic bacteria has been demonstrated to enhance immunogenicity of influenza vaccines (36,39). Probiotic bacteria influence immune responses directly by their close interaction with intestinal epithelial cells and leukocytes such as dendritic cells (DCs), and indirectly by modulation of intestinal commensal microbiota and by driving the production of metabolites such as short-chain fatty acids (36). To this end, this study was undertaken to assess the combined effect of the CpG ODN 2007 and probiotic lactobacilli on the immunogenicity of a BPL-inactivated H9N2 AIV vaccine.
In general, irrespective of the time point, the results of this study revealed that administration of a probiotic cocktail enhanced immunogenicity of intramuscularly administered BPL-inactivated H9N2 AIV, adjuvanted with CpG ODN. This was marked by increased serum HI antibody titers and IFN-γ production by chicken splenocytes. Recent studies have shown that coadministration of CpG ODN with an inactivated H9N2 AIV vaccine to chickens induces higher HI antibody responses when compared with administration of positive-control adjuvants with the vaccine (3,28). Another study compared the potential of various TLR ligands to serve as adjuvants for AIV vaccines, and the results demonstrated that CpG induces higher levels of HI titers than the other TLR ligands, poly I:C and lipopolysaccharide (LPS) (29).
In an attempt to use probiotics as a vaccine adjuvant in humans, Davidson et al. demonstrated that administration of 1010 CFU of Lactobacillus rhamnosus GG twice daily to healthy human adults as an adjuvant to an influenza vaccine induces a significantly higher HI titer at day 28 postvaccination compared with participants received placebo and vaccine (10). Similar observations are made by Yitbarek et al. when probiotic lactobacilli were administrated biweekly to H9N2 AIV-vaccinated microbiota-depleted chickens, indicating the immunomodulatory potential of the probiotic bacteria (39).
Consistent with these observations, this study demonstrated that administering probiotic lactobacilli to chickens vaccinated with CpG-adjuvanted AIV induced a significantly higher HI titer than all other treatments. A previous study described a positive correlation between the gene expression of TLR5, which recognizes bacterial flagellin, and the magnitude of the HI response leading to the suggestion of links between innate responses and antibody-mediated responses to influenza vaccination (21). Moreover, stimulation of chicken macrophages with probiotic lactobacilli strains has shown significant antiviral and immunostimulatory effects in vitro (26).
Together, these studies suggest the possible role of lactobacilli when combined with CpG to stimulate innate responses and ultimately enhance adaptive immune responses. We speculate that the administration of the probiotic cocktail and CpG in this study may have a role in modulating innate responses through activation of TLR signaling pathways, which results in increased production of chemokines and inflammatory cytokines followed by the recruitment of immune system cells such as DCs and T cells, resulting in B cell activation and antibody production.
Although IgM and IgY antibodies and HI titers were increased in the group that received a combination of probiotics and CpG with the vaccine, this was not associated with an increase in antibody avidity or virus-neutralizing antibody titers in this group compared with other group. However, there was indeed evidence for significant induction of virus-neutralizing antibodies in all groups. It should be noted that although HI antibodies have neutralizing capacity, they only target the hemagglutinin (HA) antigen, whereas virus-neutralizing antibodies may target other antigens of AIV, such as the neuraminidase (NA) antigen. Therefore, the lack of correlation between HI and virus-neutralizing titers in the combined treatment group may be related to the fact that although these types of antibodies have similar functions, they target different antigens.
In addition to the notable increase in HI titers, treatment with probiotics also enhanced cell-mediated immune responses in CpG-AIV-vaccinated chickens, as demonstrated by the increased expression of IFN-γ production in splenocytes isolated from immunized chickens and restimulated ex vivo with the vaccine antigen compared with all other groups. IFN-γ is a cytokine with a major role in innate and adaptive immunity and functions as the primary activator of macrophages and plays a crucial role in host defence against viral infections (11). IFN-γ is also a stimulator of natural killer cells. In healthy mice, oral administration of a probiotic lactobacilli strain enhances the production and secretion of cytokines, including tumour necrosis factor-α, IFN-γ and IL-10, resulting in improved immune responses against Salmonella (9). In chickens, oral treatment with probiotics significantly increases gene expression and secretion of IFN-γ proteins by splenic mononuclear cells in response to Newcastle disease virus vaccine (5). Probiotic-mediated activation of cell-mediated immune responses may suggest one potential mechanism by which protection conferred by vaccines could be improved.
In conclusion, the results of this study revealed that probiotic lactobacilli could enhance both cell- and antibody-mediated responses when administered with a CpG ODN-adjuvanted vaccine. However, further investigations are needed to determine the mechanisms by which lactobacilli and CpG interact to enhance immune responses induced by the AIV vaccines and to investigate whether different treatment modalities and combinations with other TLR ligands may result in higher magnitude and/or better-quality immune responses.
Footnotes
Acknowledgments
We thank the staff of the isolation unit at Ontario Veterinary College at the University of Guelph for their help with the care and housing of chickens.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
Funding was partly provided by Agriculture and Agri-Food Canada, the Ontario Ministry of Agriculture Food and Rural Affairs, NSERC, Egg Farmers of Canada, Chicken Farmers of Saskatchewan, and the Canadian Poultry Research Council. This research is supported in part by the University of Guelph's Food from Thought initiative, thanks to funding from the Canada First Research Excellence Fund.