
Abstract
This study aimed to explore the effect of the immunopotentiator CVC1302 on foot-and-mouth disease (FMD) vaccination in animals placed under oxidative stress. We established oxidative stress models using porcine circovirus type 2 (PCV2)-infected PK-15 cells and mice model both in vitro and in vivo, respectively. The efficacy of CVC1302 on PK-15 cells or in addition to the FMD vaccine was evaluated by quantitative real-time polymerase chain reaction, histopathological and enzyme-linked immunosorbent assay (ELISA) analysis. CVC1302 affected apoptosis of PCV2-infected PK-15 cells and significantly inhibited PCV2 replication, while it had no effect on the viability for blank PK-15 cell in vitro test with varying dilutions of CVC1302. Results showed that PCV2 induced a strong oxidative stress response in mice. CVC1302 reduced the viral load in spleen of PCV2-infected mice and ameliorated the pathological injury of spleen. Furthermore, CVC1302 significantly increased IgG antibody titer, cytokine expression, superoxide dismutase activity, catalase concentrations, and glutathione content in mice immunized with FMD vaccine. In conclusion, CVC1302 inhibits PCV2 replication and regulates oxidative stress in PCV2-infected mice, which can improve the immune efficacy of the FMD vaccine, providing a safe and effective immune enhancement.
Introduction
Oxidative stress is a state that often exists in animals. Changes in dietary nutrition, environment, feeding mode, and bacterial and viral infections can all cause oxidative stress in animals (15,19). Oxidative stress can lead to the peroxidation of phospholipids, proteins, and cell membranes, as well as cause damage to DNA (29). Lots of free radicals are abruptly produced due to abnormal metabolism, when animals are subjected to stress stimulation or virus infection. The excessive free radicals will exceed the reduction capacity of the antioxidant system, which leaves the body in a state of oxidative stress, resulting in damage to organs and tissues (3,28). Vaccination under oxidative stress is likely to lead to immune failure and thus affect the growth and development of the body, causing varying degrees of harm (18).
Several studies have reported that porcine circovirus type 2 (PCV2) infection can destroy the immune system, cause oxidative stress in immune cells, and reduce the immunity of the animal (12,14). PCV2 infection can induce the apoptosis of B lymphocytes and reduce the number of antigen-presenting dendritic cells. T cells cannot therefore recognize antigens, thus causing immunosuppression, poor immune effect, or even failure of other vaccines, ultimately reducing the immune protection rates of vaccines (8,30). In our previous study, an immunopotentiator CVC1302 was determined to be an agonist for toll-like receptors, which successfully improved the efficacy of serotype O foot-and-mouth disease virus (FMDV)-inactivated vaccine, generating long-lived antibody responses to provide protection against virus infection (10). The healthy animal model was researched in our previous study, but we did not examine the immune response of animals under oxidative stress. The purpose of this study was to establish oxidative stress models of PCV2 in vitro and in vivo, using PCV2-infected PK-15 cells and mice, respectively, to verify the effect of CVC1302 on FMD vaccination in mice under oxidative stress caused by PCV2 infection. This study provides a more powerful basis for the promotion and use of the immunopotentiator CVC1302, and to better resolve the practical problems in production.
Materials and Methods
Cell culture, virus, immunopotentiator, and vaccines
Porcine kidney epithelial 15 (PK-15) cells were obtained from the Institute of Veterinary Immunology & Engineering, Jiangsu Academy of Agricultural Sciences, Nanjing, China, and maintained in Minimal Eagle's medium (MEM; Gibco) supplemented with heat-inactivated 10% fetal bovine serum (Gibco), penicillin (100 U/mL), and streptomycin (100 μg/mL) at 37°C in a humidified atmosphere containing 5% CO2.
PCV2-NJ strain was provided by the Institute of Veterinary Immunology & Engineering, Jiangsu Academy of Agricultural Sciences, and amplified in PK-15 cells. PCV2 titers were determined to be 106.0 TCID50/mL using the Reed-Muench assay and diluted with culture medium to 2 × 105.0 TCID50 for subsequent experiments.
CVC1302 was prepared as an adjuvant for a water-in-oil emulsion formulation. In brief, the immunopotentiator CVC1302 is composed of muramyl dipeptide, monophosphoryl lipid A, and β-glucan, which were purchased from InvivoGen (San Diego, CA). All components were simultaneously dissolved in sterile water as the aqueous phase, and then mixed thoroughly with the oil phase. The recipe for the adjuvant was as described in the Chinese patent license with the registered number 201310042983.0.
Commercial serotype O FMDV-inactivated vaccine was purchased from the China Agricultural Vet. Bio. Science and Technology Co. Ltd., Lanzhou, China, lot no.: H170203J. One volume of adjuvant was fully mixed with nine volumes of FMDV-killed vaccine before injection. The FMDV vaccine mixed with CVC1302 was named FMD-CVC1302.
Cell viability determination by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide
PK-15 cells (3 × 104/well with PCV2; multiplicity of infection [MOI] = 1) were cultured for 24 h in 96-well plates and replaced D-glucosamine medium. Simultaneously, different dilutions of CVC1302 (1:10, 1:100, 1:500) were added for an additional 24 h before being subjected to colorimetric 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay (Sigma-Aldrich, St. Louis, MO). Absorbance was measured at 490 nm with a secondary wavelength of 650 nm. All tests were performed in quadruplicate. Cell viability was calculated as % of control cells.
Experimental animals and experiment design
Five-week-old BALB/c female mice were purchased from Yang Zhou University (Yangzhou, Jiangsu, China). The study and protocol were approved by the Science and Technology Agency of Jiangsu Province, approval ID NKYVET 2015-0066, granted by the Jiangsu Academy of Agricultural Sciences Experimental Animal ethics committee. All animal studies were performed in strict accordance with the guidelines of Jiangsu Province Animal Regulations (Government Decree No. 45).
In total, 54 mice were randomly divided into three groups with 18 mice per group. Mice were inoculated with 1 mL of PCV2 virus (106 TCID50) and PCV2 virus dilution (2 × 105 TCID50) through intraperitoneal injection. Phosphate-buffered saline (PBS) instead of virus dilution was used in the control group. Postchallenge, the clinical symptoms of all animals were recorded daily. On days 7, 14, and 21 postchallenge (dpc), three mice were sacrificed in each group. Spleens were collected immediately and stored at −80°C until use.
To establish PCV2-induced oxidative stress in BALB/c mice followed by CVC1302 treatment, 120 mice were randomly divided into four groups with 30 mice per group. Mice were inoculated with 1 mL of PCV2 virus dilution (2 × 105 TCID50) through intraperitoneal injection termed PVC2. PBS instead of virus dilution was used in the control group. On day 7 postchallenge, two groups of mice were subcutaneously vaccinated with the FMD vaccine, with or without CVC1302 (100 μL/per mouse), termed PCV2-FMD-CVC1302 and PCV2-FMD.
Quantification of viral load in spleen tissue
Spleen tissue samples were randomly obtained from infected mice on different days after challenge. The forward and reverse primers for PCV2 quantitation were 5′-CCGCGGGCTGGCTGAACTT-3′ and 5′-ACCCCCGCCACCGCTACC-3′, respectively. Total DNA was isolated from spleen tissue using DNA mini kits (Takara, Dalian, China) following the manufacturer's instructions. The PCV2 gene fragment was amplified using polymerase chain reaction (PCR), and a segment of 1154 bp was detected through electrophoresis using 1% agarose gels containing 0.5 μg/mL ethidium bromide.
Fluorescence assay for reactive oxygen species
Spleen tissue samples were randomly obtained gnotobiotically and pressed through a steel strainer using the flat end of a plunger from a 1 mL syringe. The strainer was rinsed with 10 mL Roswell Park Memorial Institute (RPMI) 1640, and the filtered solution was centrifuged at 1,500 rpm for 5 min at 4°C. The supernatant was carefully decanted, and the pellet resuspended in precooled Tris-NH4Cl for 2 min, then centrifuged again at 1,500 rpm for 5 min at 4°C. The supernatant was decanted, and the pellet was resuspended in RPMI 1640 twice to wash away residual media. The cells were stained with trypan blue, and live cells were counted. Viability was >90%, and cell concentration was adjusted to 2 × 106/mL. The cells were grown in black 96-well plates with transparent bottoms for reactive oxygen species (ROS) analysis using the fluorescent probe, dichlorodi-hydrofluorescein diacetate (DCFH-DA) (Sigma). After removing the culture medium, the cells were washed with PBS three times. Then, 50 μL of DCFH-DA (10 μM/L) was added to each well and incubated in the dark for 30 min at 37°C. The cells were washed with PBS three times, and fluorescent intensity was measured at 485 nm for excitation and 530 nm for emission on an EnSpire automatic microplate reader (PerkinElmer, Waltham, MA).
Quantitative real-time PCR
PK-15 cells (3 × 104/well) infected with PCV2 (MOI = 1) were cultured for 24 h in 96-well plates and replaced D-glucosamine medium. Simultaneously, different dilutions of CVC1302 (1:10, 1:100, and 1:500) were added for an additional 24 h before detection of cell apoptosis and virus replication. RNA and DNA were extracted from PK-15 cells, and subjected to real-time PCR using a Prime Script TM II Strand cDNA Synthesis kit (Takara), according to the manufacturer's protocol. Real-time PCR was performed for each DNA sample in duplicate using Bright Green 2 qPCR Master Mix (ABM, Richmond, Canada) in a Roche Light Cycler 480 (Roche, Basel, Switzerland). The thermal cycling program was provided as follows: an initial 30 sec at 95°C, followed by 40 cycles of 5 sec at 95°C and 20 sec at 60°C. The relevant primers are shown in Table 1.
Primer Sequences for Real-Time Polymerase Chain Reaction
GADPH, glyceraldehyde-3-phosphate dehydrogenase; PCV2, porcine circovirus type 2.
The virus copy numbers in three randomly selected mice from each group were measured by real-time quantitative PCR on different dpc. DNA was extracted from spleens, and real-time quantitative PCR was performed for each DNA sample in duplicate using Bright Green 2 qPCR Master Mix in a Roche Light Cycler 480. The thermal cycling program was provided as follows: an initial 30 sec at 95°C, followed by 40 cycles of 5 sec at 95°C and 20 sec at 60°C. The primers for PCV2 are shown in Table 1.
Histopathology
At necropsy, spleens were collected and fixed in 10% neutral buffered formalin. After fixation and embedding in paraffin wax by standard methods, sections (4 μm) were cut and stained with hematoxylin and eosin for light microscopic examination.
Serum FMDV-specific liquid-phase blocking enzyme-linked immunosorbent assay antibody and cytokine assays
Serum samples from individual mice were collected at 28 days postimmunization (dpi) and evaluated for serotype O FMDV-specific liquid-phase blocking (LPB) antibody titers using liquid-phase blocking enzyme-linked immunosorbent assay (ELISA) (LPBE) detection kits (LVRI, Lanzhou, China) in accordance with the manufacturer's instructions. In brief, 50 μL of twofold serial dilutions of serum samples and 50 μL of FMDV antigen (1:20 dilution) were added to a U-bottomed 96-well plate and incubated for 1.5 h at 37°C. Then, the mixtures were transferred into a 96-well ELISA plate precoated with rabbit anti-FMDV polyclonal antibody and incubated for 1 h at 37°C. After five washes, plates were incubated with 50 μL of guinea pig antiserum against serotype O FMDV for 30 min at 37°C. Then, 50 μL of rabbit antiguinea pig IgG/horseradish peroxidase (HRP) was added to the 96-well plate after a washing step (total of five washes with phosphate buffered solution) and incubated for 30 min at 37°C. The plate was washed, and 50 μL of the substrate/chromophore mixture (H2O2/O-phenylenediamine [OPD]) was added to each well. The plate was then incubated for 15 min at 37°C in the dark. Finally, 50 μL of stop solution was added to each well, and absorbance at 492 nm was measured within 15 min of stopping the reaction.
Serum samples from individual mice were collected at 0, 10, 20, 28 dpi, and evaluated for secreted interferon gamma (IFN-γ) and interleukin-4 (IL-4) using commercially available ELISA kits (Angle Gene Technologies, Nanjing, China). The assays were performed in accordance with the supplier's instructions.
Analysis of superoxide dismutase, catalase, malondialdehyde activities and glutathione, oxidized glutathione content in serum
Analysis of superoxide dismutase (SOD), catalase (CAT), and malondialdehyde (MDA) activities, glutathione (GSH) and oxidized glutathione (GSSG) content in serum, which were collected at 3 dpi, was performed.
The activities of SOD, CAT, and MDA were evaluated using commercial kits (Nanjing Jiancheng Bioengineering Institute, Nanjing, China) following the manufacturer's instructions.
Sera from mice were prepared for GSH and GSSG content analysis using commercial kits (Nanjing Jiancheng Bioengineering Institute) following the manufacturer's instructions. The concentrations of both GSH and GSSG were determined by standard curves of GSH and GSSG.
Statistical analysis
The experimental data were statistically analyzed using SPSS (v20.0; IBM, Armonk, NY) through two-way analysis of variance. The results are expressed as the mean ± standard deviation. p < 0.05 (*) was considered statistically significant; p < 0.01 (**) and p < 0.001(***) were considered to indicate a high degree of significance.
Results
The influence of CVC1302 on PCV2-infected PK-15 cells
To evaluate the effects of CVC1302 on PCV2-infected and healthy PK-15 cells, we examined cell viability, viral replication, and apoptosis (Fig. 1). CVC1302 had no effect on viability of healthy PK-15 cells at any concentration tested. However, three concentrations of CVC1302 (1:10, 1:100, and 1:500) significantly decreased the virus copy numbers of PCV2. Apoptosis-related chemokine Bcl-xL and Bcl-2 were obviously upregulated by adding CVC1302 at any concentration, while the expression levels of Bax and caspase-3 were downregulated than the expression levels of PCV2-infected groups.

The influence of CVC1302 on healthy PK-15 cell or infected by PCV2.
Determination of PCV2 infection in mice
PCV2 viral load was determined in infected mice (Fig. 2). A specific fragment of 1,154 bp corresponding to virus-positive control was detected from mice infected with PCV2, whereas samples collected from the PBS control group showed negative results. The DNA from the same weight spleen tissue samples were extracted and amplified. Specific bands amplified from spleen samples of the PCV2 2 × 105 TCID50 group were strong compared with the PCV2 106 TCID50 group. With increasing duration of infection, the bands became gradually weaker. The optimum dose of virus challenge was 2 × 105 TCID50 per mouse.
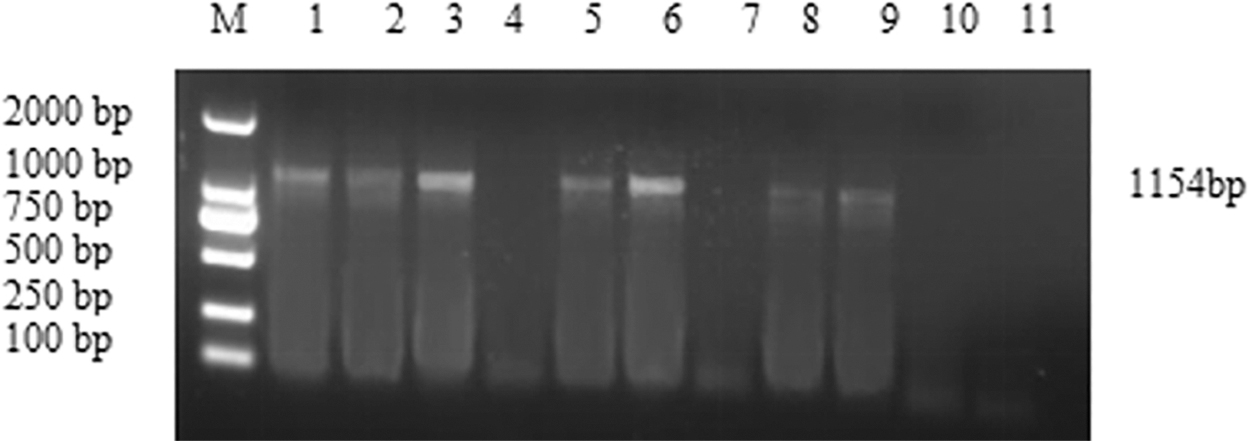
A specific nucleotide fragment of PCV2 was amplified through PCR method from the mouse spleen tissue. M: DL2000 DNA marker, (1) positive control, (2) 7 days PCV2 106 TCID50 group, (3) 7 days PCV2 2 × 105 TCID50 group, (4) 7 days PBS group, (5) 14 days PCV2 106 TCID50 group, (6) 14 days PCV2 2 × 105 TCID50 group, (7) 14 days PBS group, (8) 21 days PCV2 106 TCID50 group, (9) 21 days PCV2 2 × 105 TCID50 group, (10) 21 days PBS group, (11) H2O. PBS, phosphate-buffered saline.
Changes in the ROS production
We then investigated ROS production in spleens of PCV2-infected mice (Fig. 3). Compared with the PBS control group (3,825 ± 924.5, 3,887 ± 998.2), the 106 TCID50 PCV2 infection group (6,902 ± 1,010.3, 7,495 ± 1,176.9) and 2 × 105 TCID50 TCID50 PCV2 infection group (6,829 ± 1,065.1, 7,572 ± 1,016.9) induced significantly higher intracellular ROS production in splenic lymphocytes on 7 and 14 dpc (p < 0.001). The 106 TCID50 PCV2 infection group (4,906 ± 1,165.0) and 2 × 105 TCID50 TCID50 PCV2 infection group (4,885 ± 1,076.3) induced higher intracellular ROS production in splenic lymphocytes on 21 dpc than PBS groups (3,823 ± 945.9) (p < 0.05). Mice infected with 106 TCID50 PCV2 exhibited similar results to those infected at the lower dose of PCV2 2 × 105 TCID50.
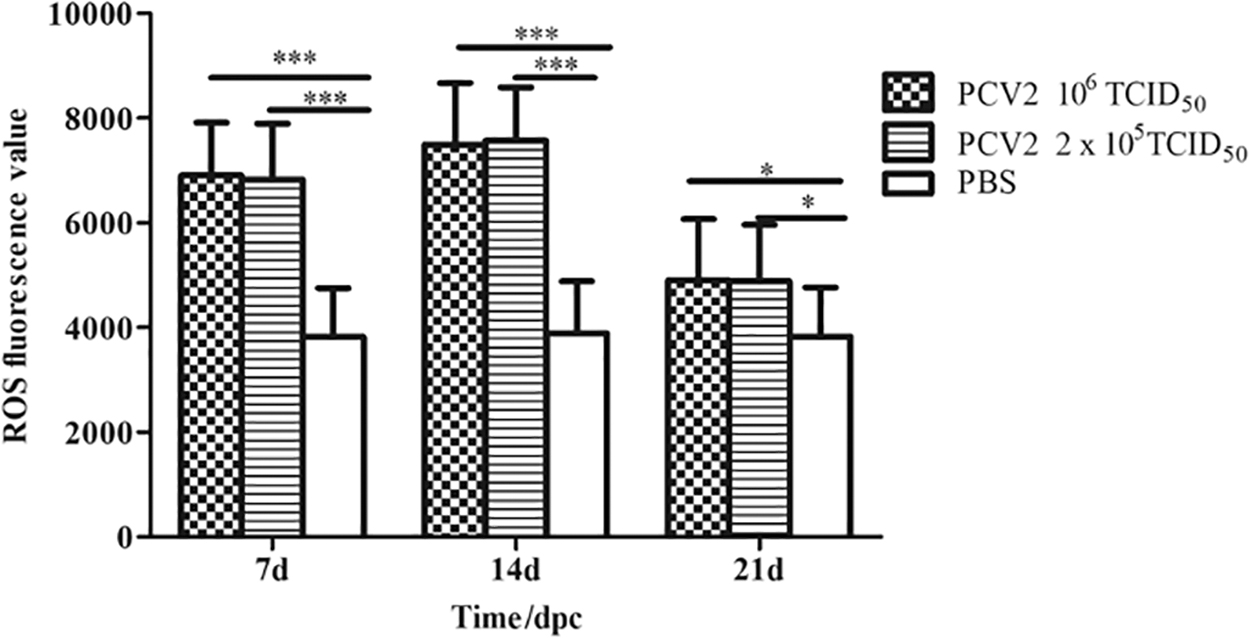
The ROS production in mice spleen lymphocytes induced by PCV2. Data were analyzed through ANOVA at 7, 14, and 21 dpc. p < 0.05 (*) was considered statistically significant; p < 0.001 (***) was considered highly significant. dpc, days postchallenge; ROS, reactive oxygen species.
Changes in PCV2-specific DNA copies
We then sought to establish the effect of CVC1302 treatment and FMD vaccination on PCV2 replication. Infected mice were vaccinated at 7 dpc with FMD vaccine with or without CVC1302. We established viral loads using real-time PCR. The copy numbers of PCV2-specific DNA in spleen samples at 0, 7, 14, 21, and 28 dpi (7, 14, 21, 28, and 35 dpc) are shown in Figure 4. At 7 and 14 dpi, the levels of PCV2-specific DNA in the PCV2-FMD-CVC1302 group, the PCV2-FMD group, and PCV2 challenge control were 4.5 ± 0.460 copies/0.1 g and 4.0 ± 0.376 copies/0.1 g; 6.9 ± 0.390 copies/0.1 g and 6.6 ± 0.298 copies/0.1 g; and 7.1 ± 0.460 copies/0.1 g and 6.62 ± 0.378 copies/0.1 g, respectively. The PCV2 virus copies in PCV2-FMD-CVC1302 were lower than those in the PCV2-FMD and PCV2 groups (p < 0.01). However, at 21 and 28 dpc, the levels of PCV2 DNA in all groups were similar.
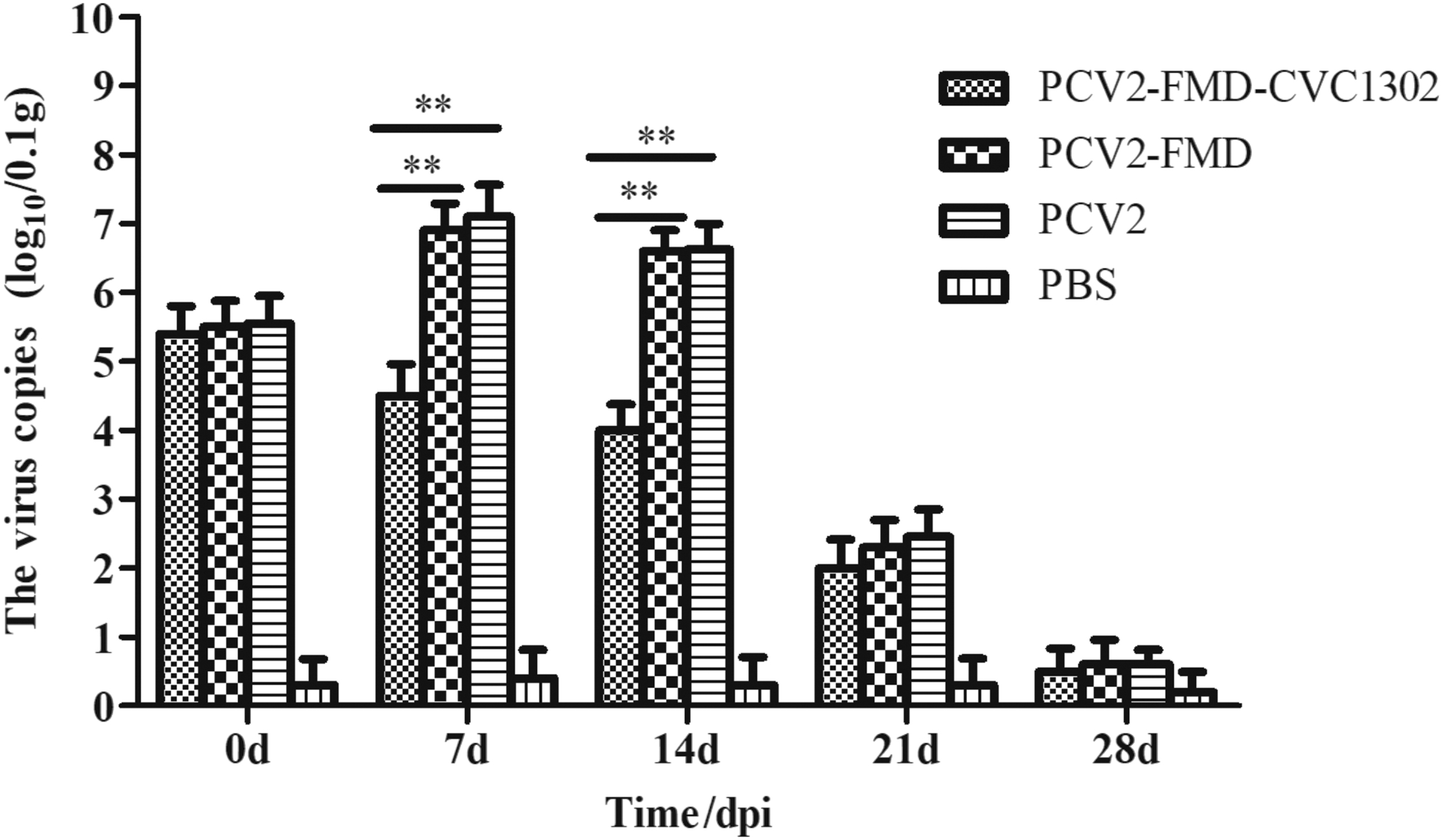
PCV2-specific RNA copies in spleen. Data are analyzed using ANOVA. p < 0.01 (**) was considered highly significant.
Histopathological analysis
We further investigated histopathological lesions in spleens of infected mice (Fig. 5). The structure of spleen nodules in the PBS group showed no significant histopathological changes. In contrast, severe macrophage infiltration and karyorrhexis of spleen cells were observed in the PCV2 and PCV2-FMD groups. Furthermore, the histopathological changes in the PCV2 group were more serious than those in PCV2-FMD group. Conversely, mild microscopic lesions were observed in the PCV2-FMD-CVC1302 group.
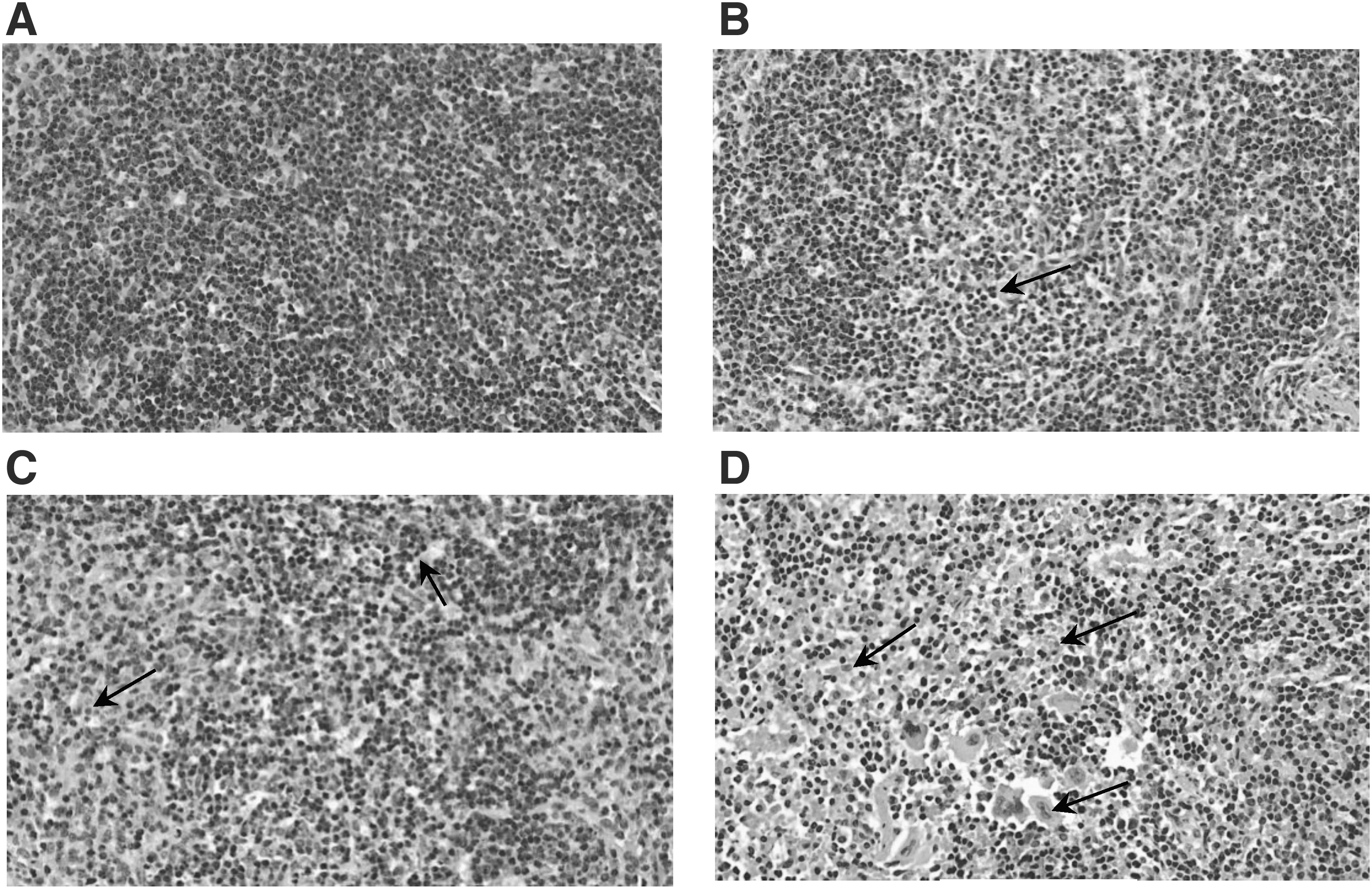
Microscopic lesions in spleen at 28 dpc (HE stain, 400 × ).
Changes in serum LPB-ELISA antibody titers
Serotype O FMDV-specific humoral responses were characterized in mice inoculated with PCV2-FMD-CVC1302 and PCV2-FMD. The levels of serotype O FMDV-specific LPB IgG antibody titers were observed between the two groups at 28 dpi (Fig. 6); the LPB-ELISA antibody titer in PCV2-FMD-CVC1302 group was 6.5 ± 0.428 log2, which was significantly higher than that observed in the PVC2-FMD group, 3.2 ± 0.285 log2 (p < 0.001).

The serotype O FMDV-specific LPB antibody responses induced by FMD-CVC1302 and FMD. Serum samples were collected at 28 dpi. Data were analyzed through t-test. p < 0.001 (***) was considered highly significant. FMDV, FMD virus; dpi, days postimmunization; LPB, liquid phase blocking.
Changes in serum cytokine concentration
Serum concentrations of IFN-γ and IL-4 are investigated in Figure 7. At 28 dpi, the concentrations of the two cytokines peaked. In the PCV2-FMD-CVC1302 group, the IFN-γ concentration was 206.28 ± 14.70 pg/mL, which was significantly higher than that in the PCV2-FMD group (130.4 ± 12.68 pg/mL) (p < 0.01) (Fig. 7A), while the IL-4 concentration was 44.1 ± 1.79 pg/mL, also significantly higher than that in the PCV2-FMD group (30.5 ± 1.56 pg/mL) (p < 0.01) (Fig. 7B). In the PBS and PCV2 groups, the levels of these two cytokines in the sera were low or undetectable (data not shown).
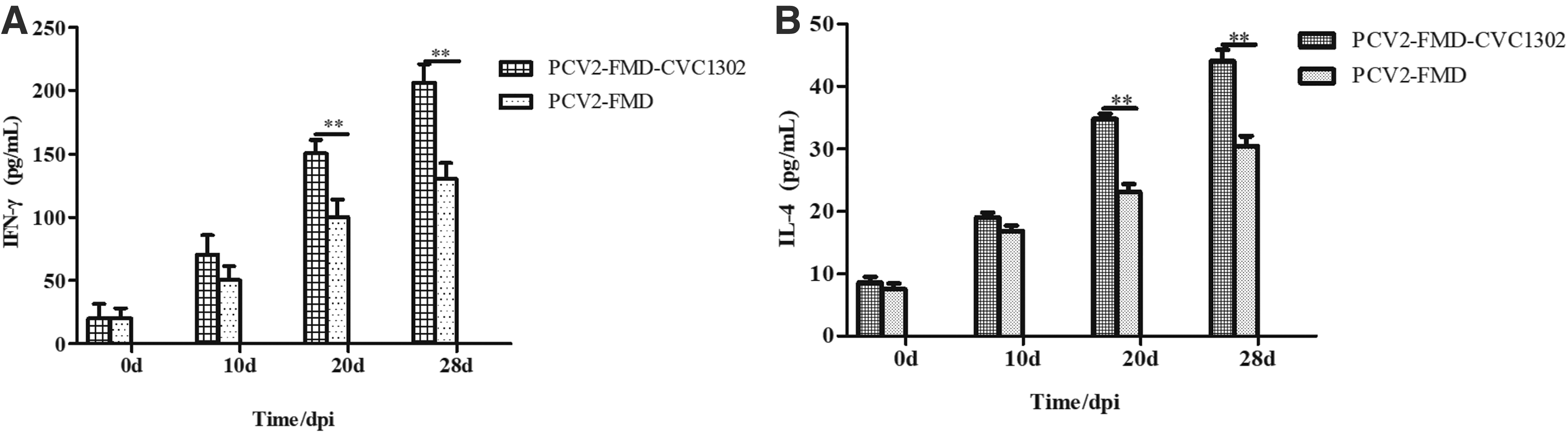
Cytokine levels in serum induced by FMD-CVC1302 and FMD.
MDA, SOD, and CAT activities
PCV2 infection caused the changed of MDA activities are shown in Figure 8, but the level of MDA in the PCV2-FMD-CVC1302 (5.45 ± 1.17 nmol/L) and PBS groups (5.12 ± 1.50 nmol/L) was significantly lower than that in the PCV2-FMD group (40.67 ± 1.92 nmol/L) (p < 0.05). Compared with the PCV2-FMD group (188.52 ± 20.41 U/mL), SOD activities were significantly higher in the PCV2-FMD-CVC1302 groups (290.8 ± 21.65 U/mL) (p < 0.001). The CAT activity in the PCV2-FMD-CVC1302 group was 307.99 ± 19.21 U/mL, which was significantly higher than that in the PCV2-FMD group, 184.33 ± 18.45 U/mL (p < 0.001).

MDA activity, SOD activity, and CAT activity of serum in PCV2-infected mice. Data are presented as the mean ± SD and statistically analyzed using ANOVA. p < 0.05 (*) was considered statistically significant; p < 0.001 (***) was considered highly significant. CAT, catalase; MDA, malondialdehyde; SOD, superoxide dismutase.
GSH and GSSG content
PCV2 infection induced significant decreases in GSH levels and the GSH to GSSG ratio (Fig. 9); the GSH content in PCV2-FMD-CVC1302 group was 20.42 ± 2.825 μmol/L, which was similar to that in the PBS group, 21.81 ± 2.95 μmol/L. The GSH content in PCV2-FMD group was 12.81 ± 1.990 μmol/L, significantly lower than that in the PCV2-FMD-CVC1302 group (p < 0.001) (Fig. 9A). The ratio of GSH to GSSG in PCV2-FMD-CVC1302 group was 4.68 ± 1.00, which was significantly higher than that in PCV2-FMD group, 1.39 ± 0.85 (p < 0.01) (Fig. 9B). Furthermore, the ratio of GSH to GSSG in PCV2-FMD-CVC1302 group showed a similar trend to the PBS group.

GSH concentration, GSSG content
Discussion
PCV2-infected animals are often in a state of chronic oxidative stress that could lead to immunosuppression by accumulation of free radicals caused by oxidative stress (17). The damage caused by viral challenge generates extra ROS, which in turn aggravates oxidative stress status (4,24). In this study, the PCV2 virus copies in PK-15 cell were 10.32 ± 0.068 log10 and specific bands amplified from mice spleen samples of the PCV2 2 × 105 TCID50 group. These results showed that a PCV2-challenge model was successfully constructed in vitro and in vivo. Two PCV2 infection groups induced significantly higher intracellular ROS production in splenic lymphocytes on 7 and 14 dpc. The results demonstrate that PCV2 infection caused oxidative stress in mice.
Infection with PCV2 can significantly reduce body weight, enhance sensitivity to various other infectious diseases, resulting in increased likelihood of mixed infections (2,7). Importantly, a state of oxidative stress can lead directly to immune failure. Therefore, the effective reduction of the oxidative stress caused by the virus is of great significance for the prevention and control of disease (1,13,21). The results of this study showed that different concentrations of CVC1302 (1:10, 1:100, and 1:500) significantly decreased the virus copies numbers of PCV2. Bcl-xl and Bcl-2 are apoptotic inhibitors, while Bax and Caspase-3 are proapoptotic factors. The changes in their mRNA expression levels determine the apoptosis or survival of host cells (16). In this study, real-time PCR analysis confirmed that the expressions of Bax and Caspase-3 were significantly upregulated after PK-15 cells were infected with PCV2. Treatment with CVC1302, Bcl-xL, and Bcl-2 were upregulated, while Bax and Caspase-3 were downregulated. These results indicate that CVC1302 not only inhibits the replication of PCV2 in PK-15 cells but also inhibits the apoptosis of cells after virus infection. In vivo model also demonstrated that CVC1302 not only reduces the viral load in the spleen but also reduces the pathological damage caused by PCV2. Furthermore, CVC1302 had an effect on the clearance of PCV2 virus in vivo and in vitro. The effective elimination of virus has a beneficial effect on the improvement of immune level.
An ideal vaccine should elicit both a humoral immune response and a cellular immune response. Immunization is the main method for the prevention and control of FMD, and the antibody levels with LPB-ELISA are used for the direct evaluation of immune effect, which being the most internationally common evaluation method (22,25). In this study, at 28 dpi, the LPB-ELISA antibody titer in the PCV2-FMD-CVC1302 group was 6.5 ± 0.428 log2, significantly higher than that in the PVC2-FMD group, 3.2 ± 0.285 log2. The levels of IFN-γ and IL-4 in the PCV2-FMD-CVC1302 group were significantly higher than those in the PCV2-FMD group. This result was consistent with that of recently published papers (6,9,20,27). These published papers have indicated that the titer of LPB-ELISA has a direct correlation with challenge protection, and cytokines are effective in virus clearance at the same time. The results of this study showed that in mice exposed to oxidative stress, those treated with CVC1302, in addition to the FMDV vaccine, not only produced high levels of LPB-ELISA antibodies but also produced high levels of cellular immune responses. Thus, CVC1302 is an effective immune enhancer that improves humoral and cellular immune efficacy after immunization with the inactivated FMD vaccine, whether the animal is healthy or under oxidative stress.
MDA, SOD, CAT, and GSH are important indexes to evaluate oxidative stress (26,31). Previous studies have documented oxidative stress in spleens of mice infected with PCV2, and the results showed that total flavonoids of Spatholobus suberectus Dunn could significantly inhibit SOD activity, GSH levels, and GSH/GSSG compensatory reduction in spleen, while also inhibiting myeloper oxidase and xanthine oxidase activities and the compensatory increase in GSSG level to restore normal levels (5,11,23). The results of this study showed that CVC1302 could significantly reduce MDA levels in the sera of PCV2-infected mice after FMD vaccination, and significantly increase SOD and CAT activities. Meanwhile, GSH and GSH/GSSG content were significantly increased and similar to PBS group. The results indicate that CVC1302 could improve antioxidant levels and protect mice from oxidative stress damage induced by PCV2 infection. In summary, CVC1302 has a regulatory effect on oxidative stress in the body and may play a role in returning levels to normal.
Overall, CVC1302 is a safe and efficient immune enhancer. It can be used not only in healthy animals but also in oxidatively stressed animals, to adjust the immune status of the body and improve the immune efficacy of vaccines. The results of this study provide support for the application of CVC1302 in clinical trials.
Footnotes
Acknowledgments
All animal experiments were performed in accordance with protocols approved by the Institutional Animal Use Committee.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by grants from the National Natural Sciences Foundation of China (31802220).