
Abstract
The staphylococcal enterotoxins (SEs) are classified as superantigens due to their potent stimulation of the immune system resulting in T cell activation and prodigious cytokine production and toxicity. This study examined the ability of superantigens to induce prophylactic antiviral activity in vivo and in vitro and evaluated potential superantigen mimetic peptides. Prophylactic treatment of mice in vivo with intraperitoneal injections of SE superantigens SEA and SEB (both at 20 μg/day for 3 days) prevented encephalomyocarditis virus (EMCV)-induced lethality in 100% and 80% of mice, respectively, as compared with control saline-treated groups in which EMCV was lethal to all mice. Furthermore, SEA (2 μg/mL) and SEB (1 μg/mL) induced antiviral activity in mouse splenocytes to produce an antiviral factor since their supernatant prevented EMCV lysis of L929 cells in tissue culture. It was found that superantigens do not directly prevent EMCV infection, but rather indirectly through inducing interferon gamma (IFNγ) production in cells as the antiviral factor. Evaluation of various superantigen mimetic peptides showed that one peptide (SEA3) had superantigen-like activity by inducing IFNγ production in cells but without the cellular proliferation, as associated with superantigens. However, the induction of IFNγ activation by the SEA3 peptide was not as pronounced, and took a much higher peptide concentration, when compared with the parent superantigen. If the negative side effects of superantigens can be eliminated, their beneficial properties can be harnessed for prophylactic treatment of viral infections and other pathologies requiring a robust immune response.
Introduction
Antiviral agents are highly desirable both as a therapeutic and prophylactic treatment for viral infections, especially for those viruses that do not have vaccines available yet. The immune system produces natural broad-spectrum antivirals similar to the various types of interferons as first responders, as part of an innate immune response, and targeted humoral antibody and cellular T cell responses, as part of a specific adaptive immune response, to combat viral infections (17,20,29,38). However, this antiviral protection may not be strong enough or is slow to develop in some individuals, especially those with immunocompromised conditions (22,32). Superantigens are potent T cell mitogens and stimulators of the immune system that could offer a robust immune response to viral infection; however, superantigens, such as the staphylococcal enterotoxins A and B (SEA and SEB, respectively), can also induce serious side effects, such as symptoms of food poisoning and toxic shock syndrome among other pathologies (15,18,23,25,35,36). Our previous studies have shown that superantigens treatment can enhance an antigen-specific antibody response in immunized mice as well as enhance immune responses against melanoma tumors in treated mice (12,37). Superantigens stimulate as many as 20% of the T cells of the body as compared with 0.001% of the T cell stimulated with “typical” antigens (15,36). This large T cell activation results in prodigious cytokine production (15,25,36). Some of the cytokines produced by superantigen treatment of animals in vivo or cells in tissue culture include IL-1, IL-2, IL-4, IL-5, IL-6, IL-12, IL-17, IL-21, TNFα, and interferon-gamma (IFNγ), which has significant antiviral and immunomodulating properties (3,4,8,15,25,26,30,36).
Unlike normal antigens, superantigens such as the staphylococcal enterotoxins (SEs) act as whole molecules binding directly to the MHC/TCR complex along with B7/CD28 complex extracellularly (23,25,36). The superantigen/MHC/TCR complex activates predominately CD4+ T cells, as well as some CD8+ T cells, resulting in cell proliferation and cytokine release (8,15,18,21,25,36). Previous studies have shown peptide sequences in both SEA and SEB superantigens that are important for their activity and binding to their receptors (1,7,11,13,21,24). Some of these peptides have been shown to mimic superantigen activity (7,11). Thus, superantigen mimetic peptides could offer therapeutic potential in which they retain the immunoenhancing properties, especially the antiviral effects, of superantigens, but reduce the toxic side effects.
In this study, we determined if the SE superantigens have antiviral inducing properties to prophylactically protect healthy mice from encephalomyocarditis virus (EMCV) infection, and determined the mechanism of this protection. Furthermore, short mimetic peptides of SE superantigens were synthesized and evaluated for their antiviral inducing and mitogenic properties in tissue culture.
Materials and Methods
Mice, virus, cells, superantigens, interferon, and media
C57BL/6 and NZW female mice were obtained from Jackson Laboratories (Bar Harbor, ME) and kept at the animal research facilities at the University of Florida (Gainesville, FL). All animal protocols were approved by the Institutional Animal Care and Use Committee at the University of Florida before any in vivo experimentation. EMCV was obtained from ATCC (Manassas, VA). L929 fibroblast and HeLa cells (both from ATCC) were maintained in EMEM (from ATCC) containing 1% penicillin/streptomycin and 10% fetal bovine serum. Staphylococcal enterotoxin A and B superantigen were purchased from Toxin Technology (Sarasota, FL). Recombinant murine IFNγ was obtained from PeproTech, Inc. (Cranbury, NJ). Human blood was purchased from Innovative Research, Inc. (Novi, MI) and histopaque-1077 (Sigma-Aldrich, Inc., St. Louis, MO) was used as the density gradient to isolate the human peripheral blood mononuclear cells (HPBMCs) from blood through centrifugation. HPBMCs were then washed in RPMI 1640 media (ATCC) and used in the in vitro experiments described as follows.
Treatment of mice with superantigens and EMCV challenge
C57BL/6 mice were prophylactically pretreated through intraperitoneal injections for 3 days with 0.1 mL of phosphate buffered saline (PBS), mouse IFNγ (3,000 U/day), SEA (20 μg/day), or SEB (20 μg/day). PBS was used as the vehicle of transfer for all intraperitoneal injections. On the last day of treatment, mice were challenged with 50 plaque forming unit (PFU) of EMCV, again through intraperitoneal injection (0.1 mL). The number of surviving mice were recorded starting on the day of EMCV challenge (day 0) and presented as percent survival. Ten mice per treatment group were used and PBS was used as the control and vehicle of transfer. In other experiments, before SEA intraperitoneal injection (naive), 24, 48, and 72 h after SEA injection, NZW mice were bled from the tail and sera isolated from the blood cells. Sera were diluted (1:100) in PBS and used for western blot analysis for IFNγ detection.
Western blot assay for detection of IFNγ
Sera collected from NZW mice treated with the 0.1 mL of staphylococcal enterotoxins SEA and SEB (25 μg/mouse by intraperitoneal injection) were diluted 1:100 in PBS and run on an SDS-PAGE apparatus. After transferring onto nitrocellulose membrane, western blot analysis was performed for the detection of mouse IFNγ in each sample. After development, the blot was scanned, and the digital image was saved for further analysis. Image J software (NIH, Bethesda, MD) was used to measure gray values for the IFNγ dimer bands in the digital image to determine fold increase in IFNγ levels in sera from treated groups as compared with the control group (PBS treated).
Synthesis of superantigen peptide mimetics
Peptides were synthesized that encompass parts of the SE protein sequences, specifically for the superantigens SEA, SEB, SEC, and TSST. The amino acid sequences of the peptides were developed based on critical SE amino acid residue involved in MHC class II binding and T cell receptor binding. Table 1 lists the sequence, length, molecular weight, and design description along with the original references from which the peptides were developed from. Some peptide sequences were altered from their original sequence to improve their activity. The SEA4 peptide was used as control peptide as its sequence is a randomly scrambled version of the SEA2 peptide. The peptides for this study were synthesized at Peptides 2.0, Inc., (Chantilly, VA). Stock peptides were desalted, dissolved in DMSO at 5 mg/mL, aliquoted and kept in a −20°C freezer until use. The purity of the peptides obtained from Peptides 2.0, Inc., was >80%.
Superantigen Mimetic Peptide Sequences, Lengths, Molecular Weights, Sequence Description, and Reference
Some amino acids within each of the sequences have been changed from their original sequence to enhance activity or solubility. Other peptide sequences are a combination of critical amino acids involved in binding to the T cell receptor or MHC molecules.
Antiviral assay
A cytopathic-effect reduction assay using EMCV was used to deduce the antiviral effects of superantigen-treated cells as previously described (19). Mouse spleen cells or HPBMC were incubated with mimetic peptides for 72 h in 24-well tissue culture plates. After incubation, supernatants from each well was collected and preincubated with L929 fibroblast cells (ATCC, Manassas, VA) on a 96-well plate for 24 h. EMCV was added to the wells and incubated for an addition of 18–36 h. Cell lysis and death was determined by crystal violet staining (0.1% w/v concentration), methanol extraction, and absorbance reading at 550 nm on an MRX II Microplate Reader (Dynex Technologies, Chantilly, VA). In some experiments, stained plates were scanned and Image J software (NIH) was used to measure gray values of the cytopathic effect of the viruses.
Proliferation assay for determination of superantigen peptide stimulatory effects
HPBMC at 104 cells/well were incubated with various concentrations of SEA (100 to 1.1 ng/mL), peptides (30 μg/mL), or media (RPMI 1640; ATCC) in a 96-well plate and proliferation was quantified through incubation with WST-1 proliferation reagent kit (Clonetech Laboratories, Inc., San Francisco, CA) at either 72 or 96 h of incubation at 37°C in a CO2 incubator. Color development by WST conversion by proliferating cells was monitored after 30 min to an hour through measuring absorbance at 450 nm wavelength on an MRX II microplate reader (Dynex Technologies).
Enzyme-linked immunosorbent assay for IFNγ detection
Supernatant from proliferation assays involving superantigens and mimetic peptides as described earlier were taken at 72-h time point and evaluated for IFNγ production. An IFNγ enzyme-linked immunosorbent assay (ELISA) kit (ThermoScientific, Inc., Rockford, IL) utilizing a rabbit polyclonal anti-human IFNγ detecting antibody and a horse radish peroxidase conjugated goat anti-rabbit secondary antibody was used to screen cell supernatants for human IFNγ. The kit was used as per the manufacturer's instructions. Color development and absorbance was measured at 450 nm in an MRX II microplate reader.
Mouse brain and heart tissue extraction and homogenization
C57BL/6 mice were pretreated for 3 days through intraperitoneal injections with 0.1 mL of PBS (control), SEA or SEB (both at 20 μg/day). On the last day of treatment, mice were challenged with 50 PFU of EMCV through intraperitoneal injection (0.1 mL). On day 6 post-EMCV infection, mice were sacrificed and equal amounts of central nervous system (CNS) (brain and spinal cord) and heart tissues were extracted, homogenized, and lysed by thawing/freezing method in EMEM as previously described (19). Sample were centrifuged at 1,000 rpms and supernatants were incubated at various dilutions with murine L929 cells that were plated to confluency in a 96-well plate for detection of EMC viral cytopathic effect through staining with 0.1% crystal violet solution. After washing plates with deionized water and overnight drying, cell viability was determined by measuring the gray values of the scanned plate using Image J software (NIH).
Statistical analysis
A paired two-tailed Student's t-test was used for comparing control groups with superantigen- or peptide-treated groups for both in vitro and in vivo studies. A p-value of 0.05 or less was considered statistically significant.
Results
Prophylactic protection of mice by SE superantigens against EMCV infection
We have previously shown that SE superantigens, such as SEA and SEB, can protect mice against melanoma in vivo in an animal model for melanoma cancer by enhancing an antigen-specific immune response (12,36). We were curious if a similar prophylactic protection can be conferred by superantigens against viral infections. Healthy C57BL/6 mice were injected intraperitoneally with superantigens SEA and SEB prophylactically 3 days prior intraperitoneal EMCV challenge. Previous studies by us and others have shown that EMCV infection leads to lethality in mice within 7 days postinfection (2,19). As shown in Figure 1, all control mice succumbed to the viral infection, whereas 100% and 80% of the SEA and SEB, respectively, treated groups were protected. As others have shown that IFNγ is protective against EMCV infection in this mouse model, we used IFNγ (3,000 U/days) as a positive control, and it showed 100% protection, confirming previous studies (2,19,33). Consistent with the protective effects of the superantigens SEA and SEB, mice from the superantigen-treated groups that were sacrificed on day 6 post-EMCV challenge showed no EMCV particles in any tissue, whereas mice from the control group (PBS treated) had significant level of EMCV in their heart and CNS tissues as shown in Figure 2. The EMCV induces lethality in mice by infecting and attacking heart and neuronal tissues as part of their pathogenesis (2,19). Thus, superantigen SEA- and SEB-treated mice were protected from the viral infection and had eliminated the virus from their tissues by a robust prophylactic immune response in vivo.
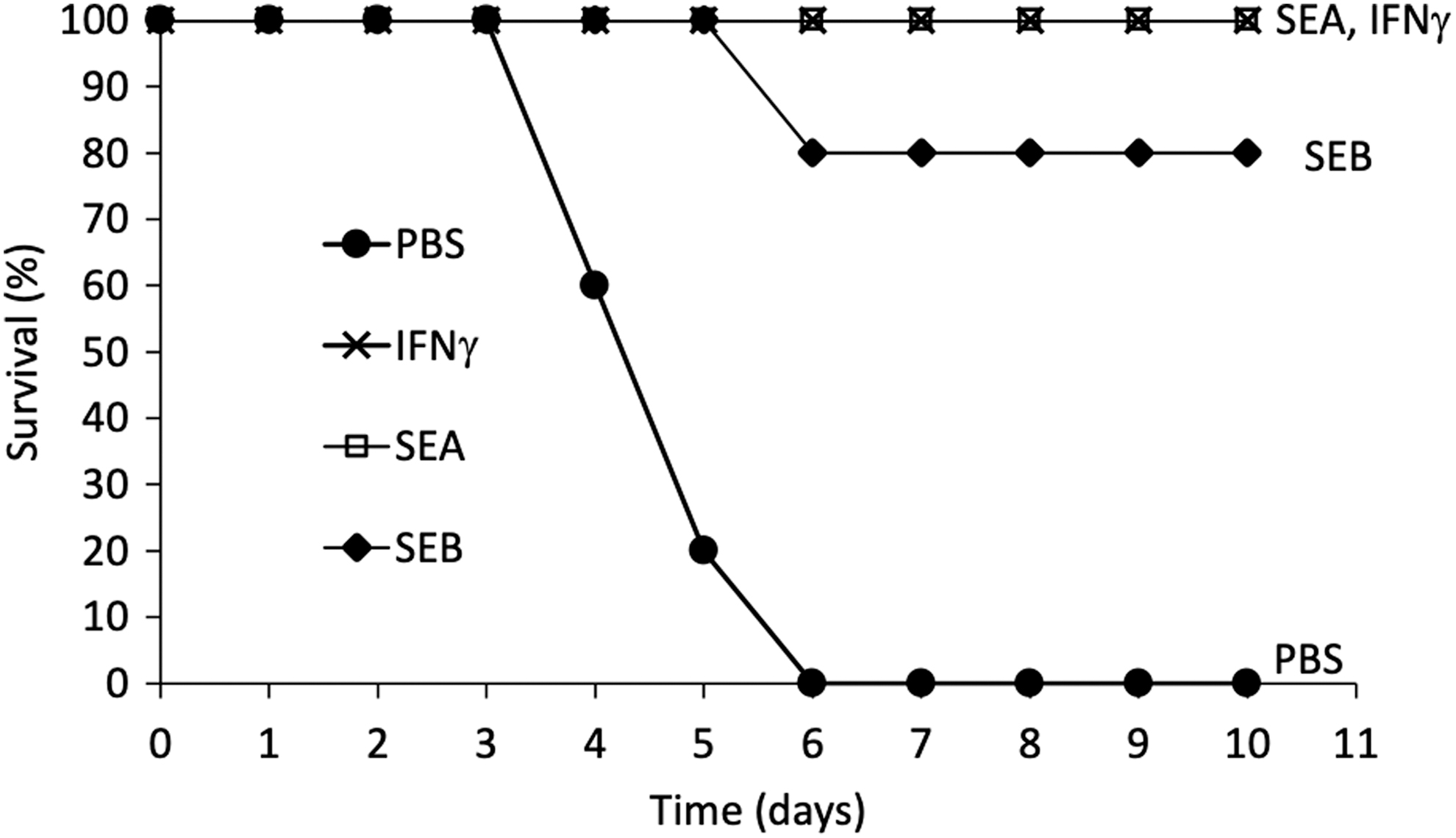
Protection of mice from EMCV challenge by prophylactic treatment with superantigens SEA and SEB. C57BL/6 mice were pretreated for 3 days through intraperitoneal injections with 0.1 mL of PBS, IFNγ (IFN, 3,000 U/day), SEA (20 μg/day), or SEB (20 μg/day). On the last day of treatment, mice were challenged with 50 PFU of EMCV through intraperitoneal injection. The number of surviving mice were recorded starting on the day of EMCV challenge (day 0) and presented as percent survival. Ten mice per treatment group were used. Both the SEA and IFNγ treatment groups had 100% survival rates for the duration of the experiment, and thus the symbols and lines overlap in the figure. EMCV, encephalomyocarditis virus; PFU, plaque forming unit.
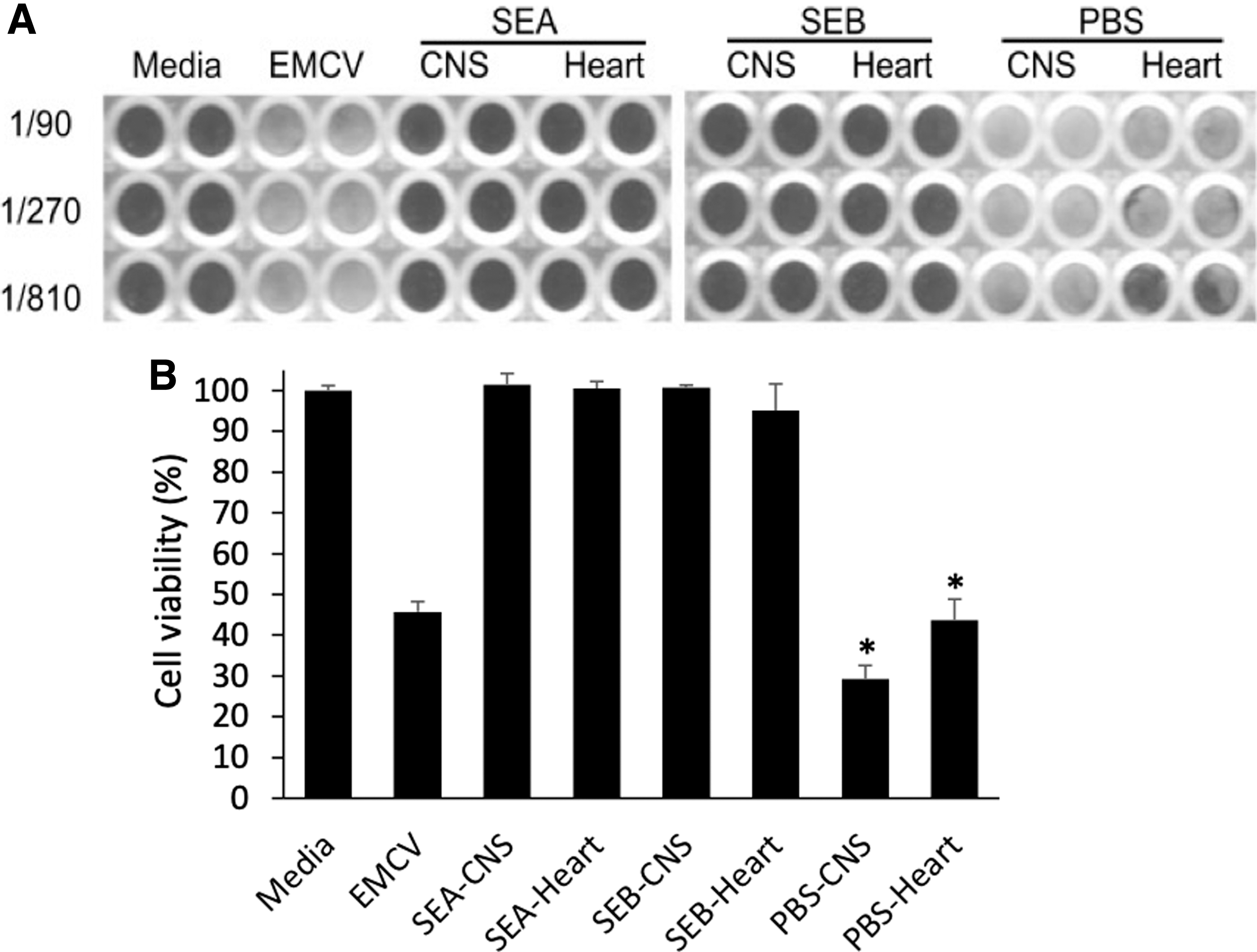
Absence of EMC viral particles from superantigen-treated mice. C57BL/6 mice were pretreated for 3 days through intraperitoneal injections with 0.1 mL of PBS, SEA or SEB (both at 20 μg/day). On the last day of treatment, mice were challenged with 50 PFU of EMCV. On day 6 post-EMCV infection, mice were sacrificed and equal amounts of CNS (brain and spinal cord) and heart tissues were extracted from each group, homogenized, and lysed by thawing/freezing. Samples were then centrifuged and supernatants were diluted and incubated on murine L929 cells plated to confluency in a 96-well plate for detection of EMC viral cytopathic effect after staining with crystal violet solution in
Superantigens induce mouse splenocytes in tissue culture to have antiviral-inducing properties
To replicate our in vivo antiviral results in vitro, naive mouse splenocytes were cultured with media, superantigens SEA or SEB (2 μg/mL) for 96 h, after which supernatants were collected and incubated with L929 mouse fibroblast cells grown to confluency. After 24 h, L929 cells were challenged with EMCV and incubated for another 24 h. Cells were stained with crystal violet for detection of cytopathic effect. As shown in Figure 3, supernatants taken from SEA and SEB splenocyte cultures had antiviral activity against the EMCV and protect the L929 cells from virus-induced cytotoxicity.

Induction of antiviral activity by SEA and SEB in vitro. NZW mouse splenocytes were treated in tissue culture with SEA (2 μg/mL), SEB (1 μg/mL), or media for 96 h after which supernatants were collected and incubated with murine L929 cells plated to confluency. After 24 h of incubation, EMCV (200 pfu/mL) was added to the wells and plates were stained with 0.1% crystal violet and picture taken as shown in
Superantigens stimulate IFNγ production to confer their antiviral activity
We next wanted to determine if this superantigen induction of antiviral effect in splenocytes is due to an intrinsic superantigen property or indirectly through induction of interferons, specifically IFNγ, which induce an antiviral state in cells (17,29,30,33,38). Superantigens are potent activators of IFNγ and other inflammatory cytokines (5,23,25,26,35). Thus, we tested antibodies against IFNγ in our viral coculture assay. As shown in Figure 4, antibodies against IFNγ neutralized the antiviral activity in supernatants collected from superantigen-treated splenocytes. Furthermore, mice that were injected intraperitoneally for 3 consecutive days with superantigens showed increasing levels of IFNγ in their sera as determined by a western blot analysis of IFNγ in mouse sera collected from tail veins (Fig. 5). As shown in Figure 1, and by others, IFNγ fully protects mice from EMCV challenge when given as a prophylactic treatment (19,33). Thus, the data here suggest that superantigens protected mice prophylactically against EMCV lethal effects by stimulating IFNγ in vivo and in vitro, and this IFNγ stimulating activity of superantigens can be offset by an IFNγ-neutralizing antibody as shown with the in vitro splenocyte assay.
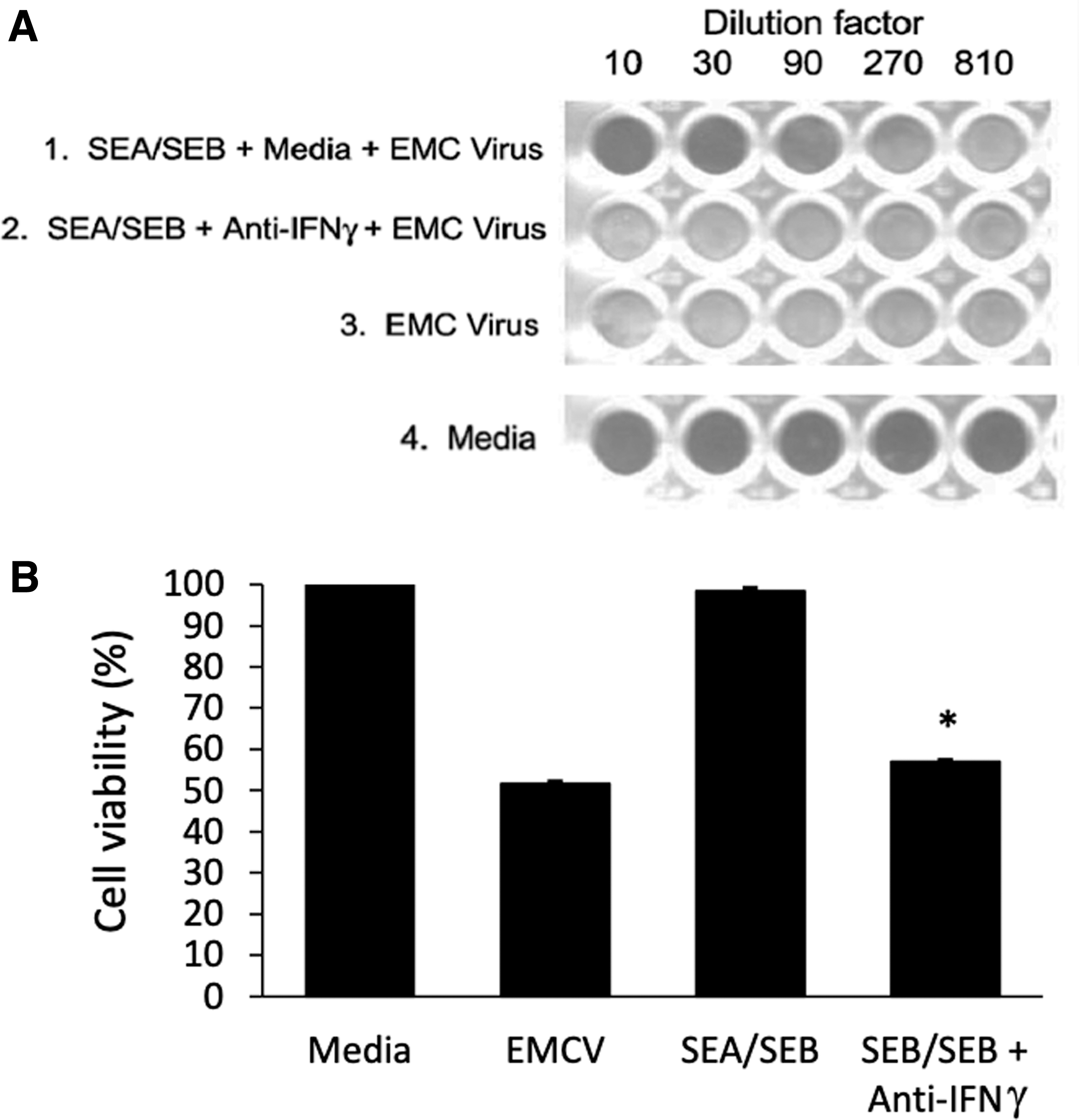
Neutralization of SEA-/SEB-induced antiviral activity by anti-IFNγ antibody. NZW splenocytes were treated with both SEA (2 μg/mL) and SEB (1 μg/mL) or media for 96 h after which supernatants were collected and incubated with media or anti-IFNγ antibody (1/200 dilution) for 2 h. Supernatants were then incubated with L929 cells plated to confluency at various dilutions shown. EMCV (200 pfu/mL) was added to the wells and cell lysis was determined by crystal violet staining. Picture was taken as shown in
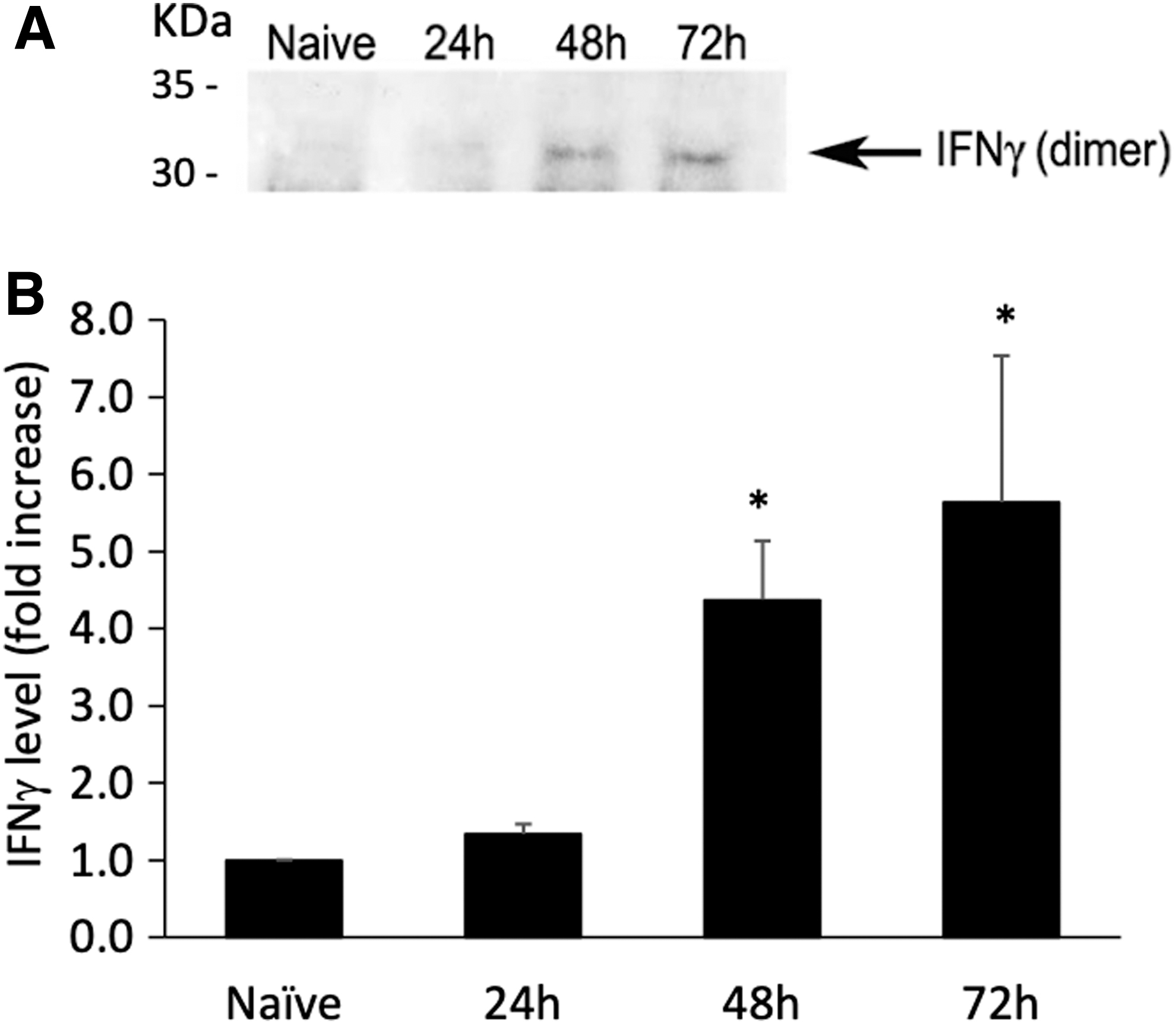
Increasing in vivo IFNγ levels detected in sera of mice after superantigen injection. NZW mice were injected with 25 μg of SEA intraperitoneally (0.1 mL). Before SEA injection (naive), 24, 48, and 72 h after SEA injection, mice were bled and sera isolated from the blood cells. Sera were diluted 1:100 in PBS and run on an SDS-PAGE apparatus. After transferring onto nitrocellulose membrane, western blot analysis was performed for the detection of mouse IFNγ dimer (detected between 30 and 35 kDa) in each sample as shown in
Evaluation of peptide mimetics of SEs for their IFNγ inducing and mitogenic activities
To reduce the enterotoxin's cellular toxicity and mitogenic activity, but keep its interferon-inducing activity, we developed and evaluated short peptide sequences that encompass various superantigens at their binding sites as shown in Table 1. Previous studies have shown sequences that are important for the SE's superantigen and receptor binding activity (1,7,11,13,21,24). Peptides were developed based on critical SE amino acid residue involved in MHC class II binding and T cell receptor binding (Table 1) and evaluated for their IFNγ stimulation in HPBMC (7, 11, 13, 14, 21, 24). Similar to results shown for superantigen induction of IFNγ in vitro in mice splenocytes, superantigens also induced IFNγ in HPBMC as determined through a human IFNγ ELISA kit (Fig. 6). Furthermore, only one mimetic peptide (SEA3) induced IFNγ production in HPBMC after 72 h of incubation as determined by the IFNγ ELISA as shown in Figure 6. However, this IFNγ induction by the SEA3 mimetic peptide was not as pronounced as that for the whole superantigen parent protein (SEA), and it took a high peptide concentration (30 μg/mL) to induce IFNγ in cells as compared with the SEA superantigen (10 ng/mL).
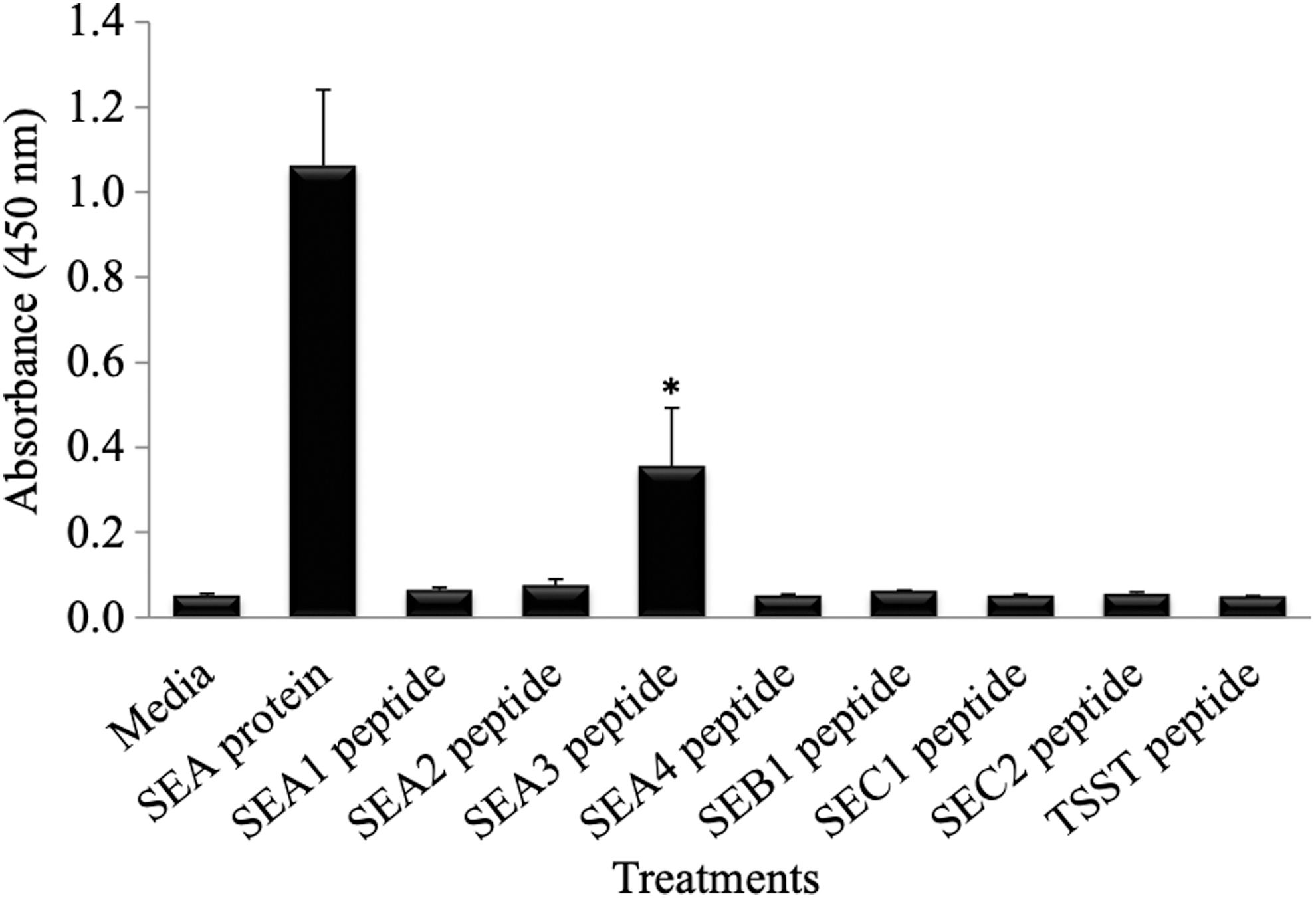
IFNγ production by HPBMC stimulated with the superantigen SEA and mimetic peptide, SEA3. HPBMCs were incubated at 104 cell/well with either SEA (10 ng/mL), mimetic peptides (30 μg/mL), or media. Supernatants were collected at the 72-h time point of incubation and evaluated for IFNγ production through an IFNγ ELISA kit. Absorbance was read at 450 nm. SEA whole protein was a positive control, and SEA4 (scrambled peptide) and media were used as the negative controls. Using a Student's t-test analysis, only the SEA3 peptide induction of IFNγ production was statistically significant (p < 0.05) when compared with media treatment. (*) means p < 0.05.
Superantigen confer their toxicity in part through their mitogenic activity by stimulating T cell proliferation, anergy, and eventual apoptosis (15,23,36). Unlike the whole SEA protein superantigen, SEA3 mimetic peptide did not induce cellular proliferation in HPBMC at 96 h as determined by a WST-1 incorporation (Fig. 7). Three of the mimetic peptides tested (SEA1, SEA2, and TSST) did have mitogenic activity and induced cellular proliferation (Fig. 7) but interestingly, did not stimulate IFNγ production (Fig. 6) in HPBMC. Thus, although superantigen whole protein was much more effective at stimulating IFNγ production, superantigens mimetic peptides may offer potential to reduce the mitogenic and toxic effects of superantigens, but still retain the IFNγ stimulating properties.
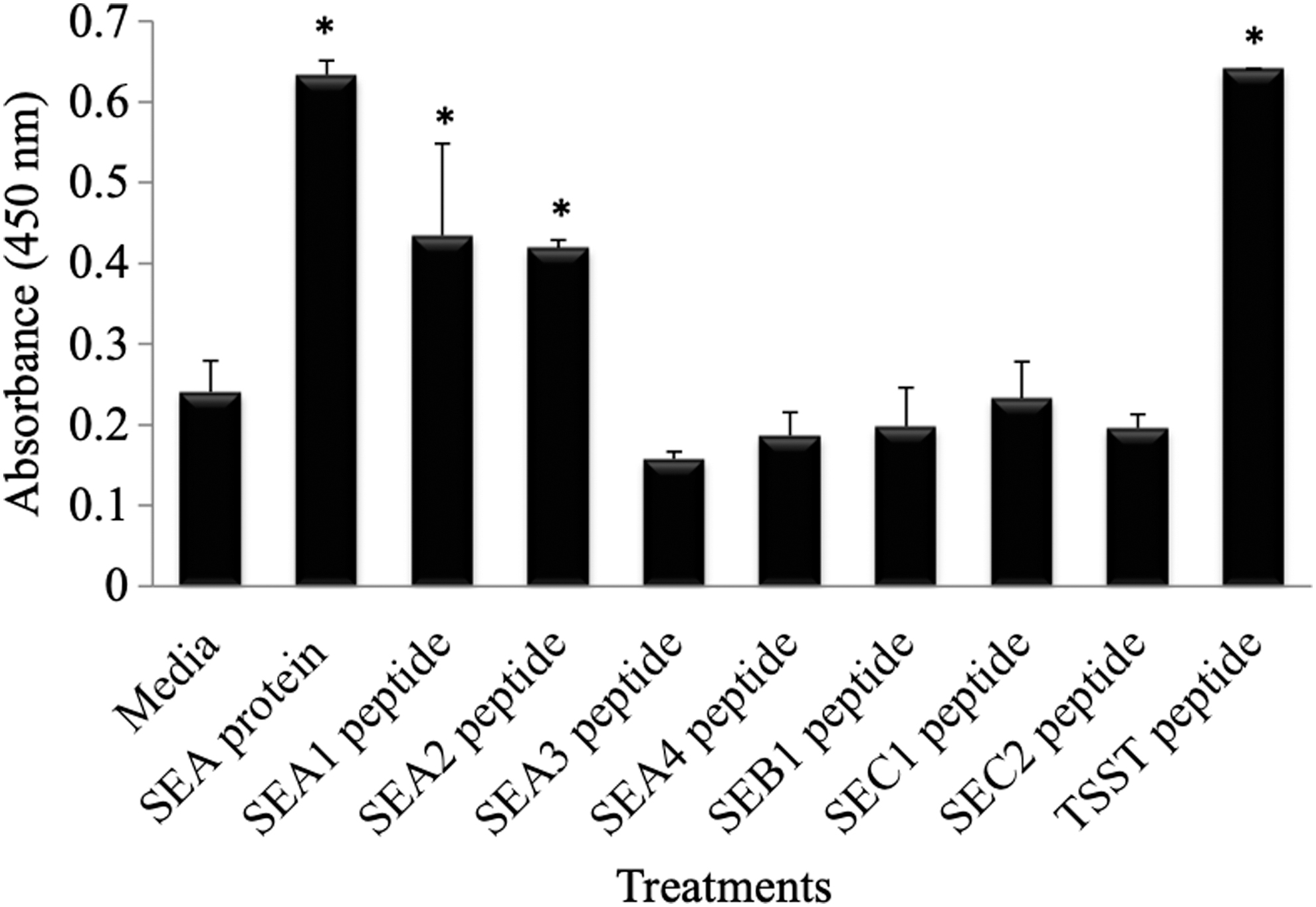
Induction of cellular proliferation by SEA superantigen and mimetic peptides. HPBMCs were incubated at 104 cell/well in a 96-well plate with either SEA (10 ng/mL), mimetic peptides (30 μg/mL), or media and proliferation was quantified through incubation with WST-1 reagent at 96 h and absorbance read at 450 nm. The SEA whole protein was positive control, and the SEA4 scrambled peptide and media were negative controls. Induction of proliferation by the SEA protein, SEA1 peptide, SEA2 peptide, and TSST peptide were statistically significant when compared with media control as determined by a Student's t-test (p < 0.05). (*) means p < 0.05.
Discussion
We have previously shown that superantigens enhance the immune response to melanoma in C57BL/6 mice (12,36). In this study, we show that the SE superantigens SEA and SEB can also be utilized to prophylactically enhance the immune system in preventing mortality in mice challenged with a lethal dose of EMCV after superantigen treatment. This protection of mice in vivo by superantigens did not occur directly by the inhibition of the viral agent, but rather indirectly through the activation of IFNγ production. IFNγ levels were upregulated in sera of mice and in tissue cultures of splenocytes upon superantigen treatment. IFNγ has been shown previously to have potent antiviral activity, including against EMCV (19,30,33). To dissociate superantigens from their mitogenic activity, and further cellular toxicity, and still retain their IFNγ inducing activity, short mimetic peptides of superantigens were developed. Of the eight mimetic peptides tested, one peptide (SEA3) induced IFNγ production to a higher level than the other peptides (Fig. 6) and did not induce cellular proliferation (Fig. 7). However, the induction of IFNγ production by the SEA3 peptide was less pronounced, and took a much higher peptide concentration, when compared with the SEA whole protein. Future studies will focus on the modification of the SEA3 peptide sequence to enhance the IFNγ inducing properties without any intrinsic mitogenic activity since superantigen-induced proliferation in T cells leads to anergy and eventual apoptosis (28,36).
IFNγ has direct antiviral activity on cells; however, it must be administered therapeutically by injection or transfusion as the protein will denature if given orally (16,19,27). Superantigens, similar to the SEs, stimulate IFNγ production, as shown in this study and by others, and can be administered by injection, orally, and/or by inhalation (9,10,36,39). They are not altered by acids, proteases, or temperature differences found in the digestive track (10,36). Other studies have shown that at very low concentrations the SEs might be tolerable with minimal associated toxicity (6,31,39). Furthermore, it has been shown that T cells that do become anergized by superantigens, continue to secrete IFNγ as well as the anti-inflammatory cytokine, IL-10, after a subsequent challenge with superantigen (5,14). Interestingly, we have shown previously that superantigens enhance the immune responses against melanoma in mice by expanding the specific T cell populations that have been already activated (12,36). A similar expansion of viral-specific T cell by superantigen might also occur if superantigens are used to treat active viral infections that have already initiated an adaptive immune response. Since the experiments outlined in our study were a prophylaxis evaluation of superantigen antiviral activity, future experiments will evaluate superantigens as a therapeutic where mice have active viral infections. Another limitation to our in vivo mouse model for evaluating superantigen's antiviral activity is that mice are more resistant to superantigen toxin exposure and resulting pathologies, whereas humans are exquisitely sensitive to them (3,34). Other studies have used lipopolysaccharide or d-galactosamine to induce animals to be more sensitive to superantigen activity, which could improve our mouse model in future experiments (3,4,34).
In conclusion, this study shows that staphylococcal superantigens protect mice prophylactically against EMCV-induced lethality, and this antiviral protection was due to the induction of IFNγ. If the immunomodulatory properties of superantigens can be harnessed and their toxic effects reduced, perhaps through production of superantigen mimetic peptides, then superantigens might be beneficial as a prophylaxis for viral infections and other pathologies requiring a vigorous immune response.
Footnotes
Authors' Contributions
Conceptualization, methodology, data curation, visualization, investigation, validation, project administration, original draft preparation, reviewing, and editing by M.G.M. Conceptualization, resources, review, and editing by H.M.J. Conceptualization, methodology, data curation, original draft preparation, visualization, investigation, and validation by J.M.P.
Acknowledgments
The authors thank our undergraduate research students, Chintak Patel, Ravi Patel, and Olga Zambrano, for their time and effort in assisting with the mouse and peptide experiments and maintaining the cell culture lines.
Author Disclosure Statement
The authors declare that they have no competing financial interests or personal relationships that could have appeared to influence the study reported in this article.
Funding Information
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.