
Abstract
CD100 is an important immune semaphorin that is a secreted and membrane bound protein involved in infectious diseases. However, CD100 expression profile and the regulation to innate immune system in hepatitis B virus (HBV)-associated acute-on-chronic liver failure (ACLF) was not previously reported. The aim of this study was to investigate CD100 level and modulatory function of CD100 to CD14+ monocytes in HBV-ACLF patients. Plasma-soluble CD100 (sCD100) level and membrane-bound CD100 (mCD100) expression on peripheral CD14+ monocytes was analyzed in HBV-ACLF patients. CD14+ monocytes-induced cytotoxicity and CD14+ monocytes-mediated T cell activation in response to CD100 stimulation was also assessed in direct and indirect contact coculture culture systems. HBV-ACLF patients had lower plasma sCD100 and higher mCD100 level on CD14+ monocytes compared with asymptomatic HBV carriers, chronic hepatitis B patients, and controls. CD14+ monocytes from HBV-ACLF patients induced limited target Huh7.5 cell death and secreted less interferon-γ (IFN-γ), tumor necrosis factor-α (TNF-α), and granzyme B in both direct and indirect contact coculture systems compared with controls. Recombinant sCD100 not only enhanced CD14+ monocytes-mediated Huh7.5 cell death and granzyme B secretion, but it also elevated CD14+ monocytes-induced IFN-γ/interleukin-17 production by CD4+ T cells as well as IFN-γ/TNF-α secretion by CD8+ T cells in HBV-ACLF patients. The current data indicated that severe inflammation induced sCD100/mCD100 imbalance to inactivate CD14+ monocytes response, which might be beneficial for the survival of HBV-ACLF patients.
Introduction
Acute-on-chronic liver failure (ACLF) is a progressive syndrome that is associated with rapid clinical worsening, acute decompensation, organ failure, and high short-term mortality (1,4). ACLF always occurs in a close relationship to a precipitating event, such as acute alcoholic, drug-induced, bacterial infection, or viral hepatitis (1). In China, hepatitis B virus (HBV) infection is still the most common severe exacerbation risk factor for development of ACLF (22). Importantly, intensive systemic inflammation is the hallmark of ACLF in the setting of immune dysfunction. Thus, HBV-associated ACLF displays a shift from low-grade inflammatory response to exaggerated inflammation (8,19). However, the inflammatory response and immune function corresponding to pathogenesis of HBV-ACLF was still not fully elucidated.
CD100 is also known as semaphorin4D, which belongs to the family of immune semaphorins and abundantly expressed on T cells, natural killer (NK) cells, and different antigen-presenting cells (7,10,12). The membrane-bound CD100 (mCD100) was digested and shed from membrane to soluble CD100 (sCD100) mainly by specific matrix metalloproteinases (24,27). CD100 plays a vital role in immune modulation by enhancement of immune cell function in various diseases, including infectious diseases (7,12,18,27) and cancers (17,24,25,28). sCD100 regulated cytokine production by monocytes as well as migration and function of antigen-presenting cells (2). CD100 overexpression closely related to tumor angiogenesis, and it prompted monocytes differentiation toward M2 macrophages in epithelial ovarian cancers (3). Moreover, there was an imbalance between mCD100 expression on T cells and serum sCD100 in regulating antiviral immune responses during HBV and hepatitis C virus (HCV) infection (12,27). Insufficient shedding of mCD100 contributed to CD8+ T cells and NK cells dysfunction in chronic viral hepatitis (7,12,27). However, expression profile of mCD100/sCD100 and regulation of CD100 to monocytes function was not previously reported in ACLF patients. Thus, we hypothesized that CD100 also promotes monocytes activity in HBV-ACLF patients. To test this possibility, sCD100 level and mCD100 expression on monocytes in HBV-ACLF patients was measured. Cytotoxicity and regulatory activity of monocytes from HBV-ACLF patients was also assessed in response to recombinant CD100 stimulation in vitro.
Patients and Methods
Ethical statement
The study was approved in compliance with the Declaration of Helsinki by the Ethics Committee at The Affiliated Hospital to Changchun University of Chinese Medicine, by the Ethics Committee at 964th Hospital of PLA, and by the Ethics Committee at Shaanxi Provincial People's Hospital. Written informed consent was obtained from all enrolled subjects.
Studied population
Eighteen asymptomatic HBV carriers (ASC) were enrolled from Shaanxi Provincial People's Hospital between June 2018 and December 2018. The diagnosis of ASC was made in accordance with HBV DNA and HBV surface antigen (HBsAg)-positive for more than 6 months with normal liver function. Thirty chronic hepatitis B (CHB) patients were enrolled from The Affiliated Hospital to Changchun University of Chinese Medicine and Shaanxi Provincial People's Hospital between July 2019 and November 2019. The diagnosis of CHB was made in accordance with HBV DNA and HBsAg-positive patients for more than 6 months and exhibition symptoms or signs of abnormal hepatic function. Twenty-eight ACLF patients were recruited from 964th Hospital of PLA and Shaanxi Provincial People's Hospital between March 2019 and November 2019. The diagnosis of HBV-ACLF was based on the underlying chronic HBV infection and the clinical criteria of recent development of jaundice (total bilirubin >170 μM) and a prothrombin activity <40%. The exclusion criteria were the following: patients with other chronic viral infections, patients with malignancies, patients with autoimmune diseases, patients with diabetes, and pregnancy. No patients received antiviral or immunomodulatory therapies before sampling. Further, 20 age- and sex-matched healthy controls were also included. The baseline characteristics of all enrolled subjects are shown in Table 1.
Clinical Characteristics of Studied Subjects
Data were shown as median and range.
ASC, asymptomatic HBV carriers; CHB, chronic hepatitis B; HBV-ACLF, hepatitis B virus-associated acute-on-chronic liver failure.
Isolation of plasma and peripheral blood mononuclear cells
Plasma were obtained by centrifuging anticoagulation peripheral blood at 1,000 g, at 4°C for 5 min. Peripheral blood mononuclear cells (PBMCs) were isolated with Ficoll-Hypaque density centrifugation reagent Histopaque-1077 (Sigma, St Louis, MO) according to the manufacturer's protocol.
Purification of CD14+ monocytes and CD3+ T cells
CD14+ monocytes were purified with CD14 MicroBeads UltraPure (Miltenyi, Bergisch Gladbach, Germany), and CD3+ T cells were purified with CD3+ T Cell Isolation Kit (Miltenyi) according to the manufacturer's protocol, respectively.
Cell culture
CD14+ monocytes were stimulated with 1 × lipopolysaccharide (LPS; eBioscience, San Diego, CA) in the presence or absence of recombinant sCD100 (final concentration: 5 μg/mL; PeproTech, Rocky Hill, NJ) for 12 h. Cells were washed twice, and 105 of stimulated CD14+ monocytes were cultured in direct contact or in a Transwell-based indirect contact with 5 × 105 of Huh7.5 cells for 48 h. In certain experiments, 105 of stimulated CD14+ monocytes were cultured in direct contact with autologous 105 of CD3+ T cells for 48 h. In the last 6 h of the culture, 2 μL of cell activation cocktail (Biolegend, San Diego, CA) and monensin (10 μg/mL) were added. Cells and supernatants were harvested for further experiments.
Quantification of sCD100 and cytokines
sCD100 level in the plasma, interleukin-1β (IL-1β), IL-6, interferon-γ (IFN-γ), tumor necrosis factor-α (TNF-α), and granzyme B level in the supernatants were measured by commercial enzyme-linked immunosorbent assay (ELISA) kits (CUSOBIO, Wuhan, Hubei Province, China) according to the manufacturer's protocol.
Flow cytometry
For CD100 staining, PBMCs were extracellular stained with fluorescein isothiocyanate (FITC)-CD14 (eBioscience), and phycoerythrin (PE)-CD100 (R&D Systems, Minneapolis, MN), and the appropriately conjugated IgG antibodies were used as isotype-matched controls, at 4°C in the dark for 20 min. The labeled cells were examined on an FACSCalibur flow cytometer (BD Bioscience, San Jose, CA). FACS results were analyzed with CellQuest Pro software (BD Bioscience) and FlowJo V10 software (Tree Star, Ashland, OR). To investigate the molecular features of CD14+ monocytes, cells were extracellularly stained with FITC-CD14 (eBioscience), allophycocyanin (APC)-Fas ligand (FasL) (eBioscience), and PE-TNF-related apoptosis-inducing ligand (TRAIL) (eBioscience) at 4°C in the dark for 20 min. The labeled cells were examined on an FACSCalibur flow cytometer (BD Bioscience). The FACS results were analyzed with CellQuest Pro software (BD Bioscience) and FlowJo V10 software (Tree Star). To investigate the molecular features of CD3+ T cells, expression of IFN-γ and IL-17 in CD3+CD4+ T cells and expression of IFN-γ and TNF-α in CD3+CD8+ T cells were assessed. Cells were first extracellularly stained with peridin chlorophyll protein (PerCP)-CD3 (BD Bioscience), PerCP Cyanine5.5-CD4 (BD Bioscience), and PE-CD8 (BD Biosciences). Cells were then treated with Cytofix/Cytoperm solution and Perm/Wash buffer (BD Bioscience), and they were intracellularly stained with APC-IL-17 (BD Bioscience), Brilliant Violent 711-IFN-γ (Biolegend), and FITC-TNF-α (Biolegend). The labeled cells were detected on a Beckman DxFLEX cytometer (Beckman Coulter, Brea, CA), and they were analyzed with CytExpert software (Beckman Coulter) and FlowJo V10 software (Tree Star).
Cytotoxic assay
Lactate dehydrogenase (LDH) level in the supernatants was measured by LDH Cytotoxicity Assay Kit (BioVision, San Francisco, CA) according to the manufacturer's protocol. The high level of LDH control was defined as the LDH level in Huh7.5 cells with Triton X-100 treatment, whereas the low level of LDH control was defined as LDH level in cultured Huh7.5 cells. The cytotoxicity was calculated by using the following formula: (high level LDH control–LDH level of sample)/(high level LDH control–low level LDH control) × 100%.
Statistical analysis
Data were presented as mean ± standard deviation, and they were statistically analyzed by using SPSS V19.0 software (Chicago, IL). One-Way ANOVA, SNK-q test, Student t test, and paired t test were used for comparison. A p-value <0.05 was considered statistically significant.
Results
sCD100 level was downregulated in HBV-ACLF and CHB patients
Plasma sCD100 level in all enrolled subjects was measured by ELISA. sCD100 expression in the plasma from both CHB patients (259.9 ± 60.48 pg/mL) and HBV-ACLF patients (223.1 ± 67.42 pg/mL) was downregulated when compared with ASC (302.8 ± 69.60 pg/mL) and controls (299.0 ± 53.55 pg/mL) (SNK-q test, p < 0.05, Fig. 1). sCD00 level was also reduced in HBV-ACLF patients when compared with CHB patients (SNK-q test, p = 0.033, Fig. 1). There was no remarkable difference of sCD100 level between controls and ASC (SNK-q test, p = 0.852, Fig. 1). There were no significant correlations between sCD100 level and clinical index (p > 0.05).
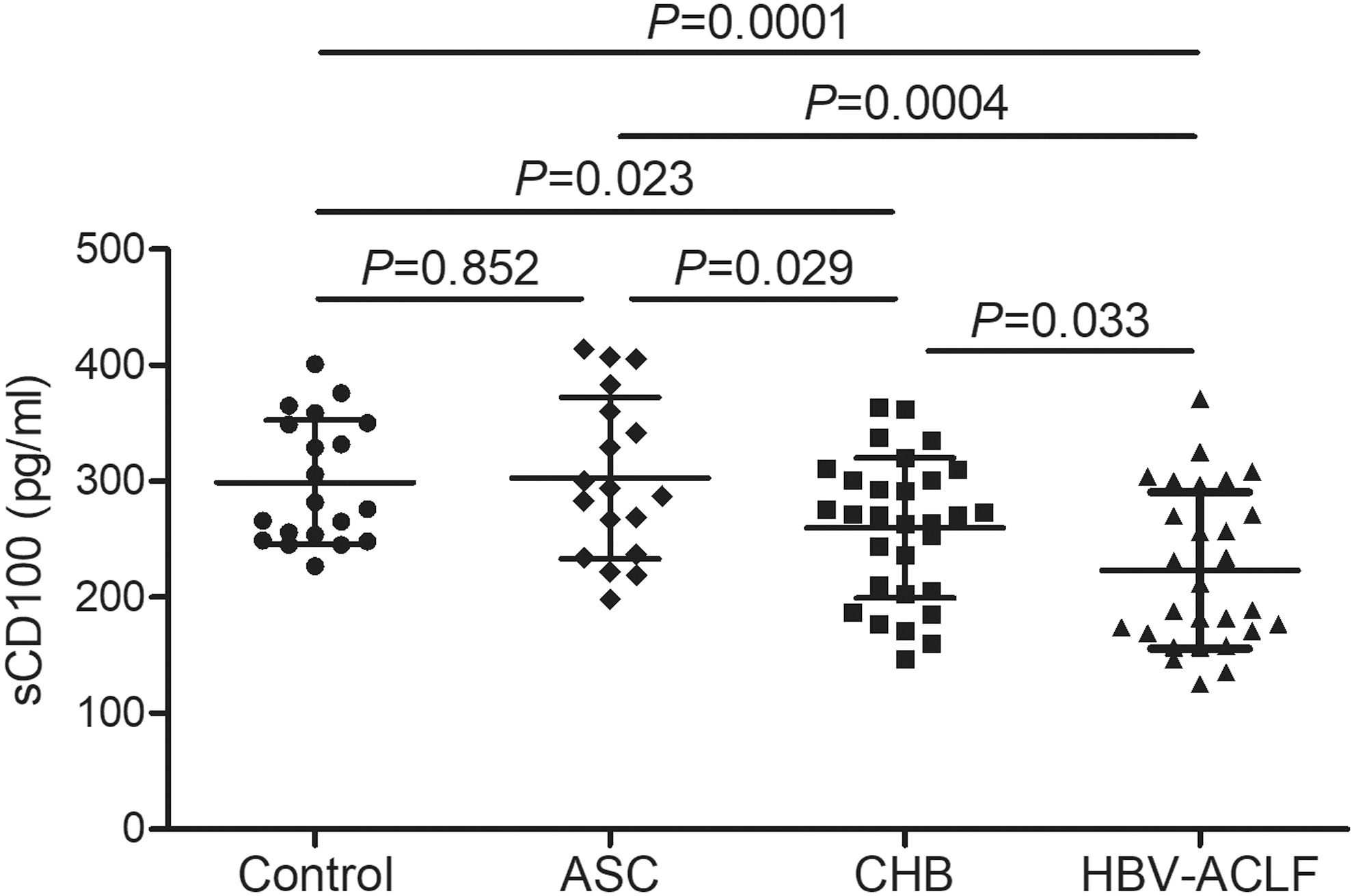
sCD100 level was downregulated in ASC, CHB patients, and HBV-ACLF patients. sCD100 level in plasma was measured by ELISA in healthy controls (n = 20), ASC (n = 18), CHB patients (n = 30), and HBV-ACLF patients (n = 28). Plasma sCD100 level was downregulated in CHB patients and HBV-ACLF patients when compared with ASC and controls. sCD100 expression was also reduced in HBV-ACLF patients when compared with CHB patients. Individual level of each subject was shown. The horizon line represented the mean, whereas the bar represented the standard deviation. One-Way ANOVA and SNK-q test was used for comparison among groups. ASC, asymptomatic HBV carriers; CHB, chronic hepatitis B; HBV-ACLF, hepatitis B virus-associated acute-on-chronic liver failure.
mCD100 level was upregulated in HBV-ACLF patients
The percentage of CD14+mCD100+ cells and the mean fluorescence intensity (MFI) of mCD100 in CD14+ monocytes was assessed by flow cytometry. The representative flow data for isotype control, mCD100 in CD14+ monocytes was shown in Figure 2A. The percentage of CD14+mCD100+ cells within CD14+ monocytes was upregulated in HBV-ACLF patients (79.51% ± 7.63%) when compared with controls (63.80% ± 11.11%, SNK-q test, p < 0.0001, Fig. 2B), ASC (62.94% ± 10.43%, SNK-q test, p < 0.0001, Fig. 2B), and CHB patients (63.43% ± 10.31%, SNK-q test, p < 0.0001, Fig. 2B). Similarly, MFI corresponding to mCD100 in CD14+ monocytes was also elevated in HBV-ACLF patients (331.6 ± 63.20) when compared with controls (224.6 ± 64.71, SNK-q test, p < 0.0001, Fig. 2C), ASC (250.3 ± 56.60, SNK-q test, p < 0.0001, Fig. 2C), and CHB patients (242.3 ± 55.38, SNK-q test, p < 0.0001, Fig. 2C). There were no significant differences of CD14+mCD100+ percentage or mCD100 MFI among ASC, CHB patients, and controls (SNK-q test, p > 0.05, Fig. 2B, C).
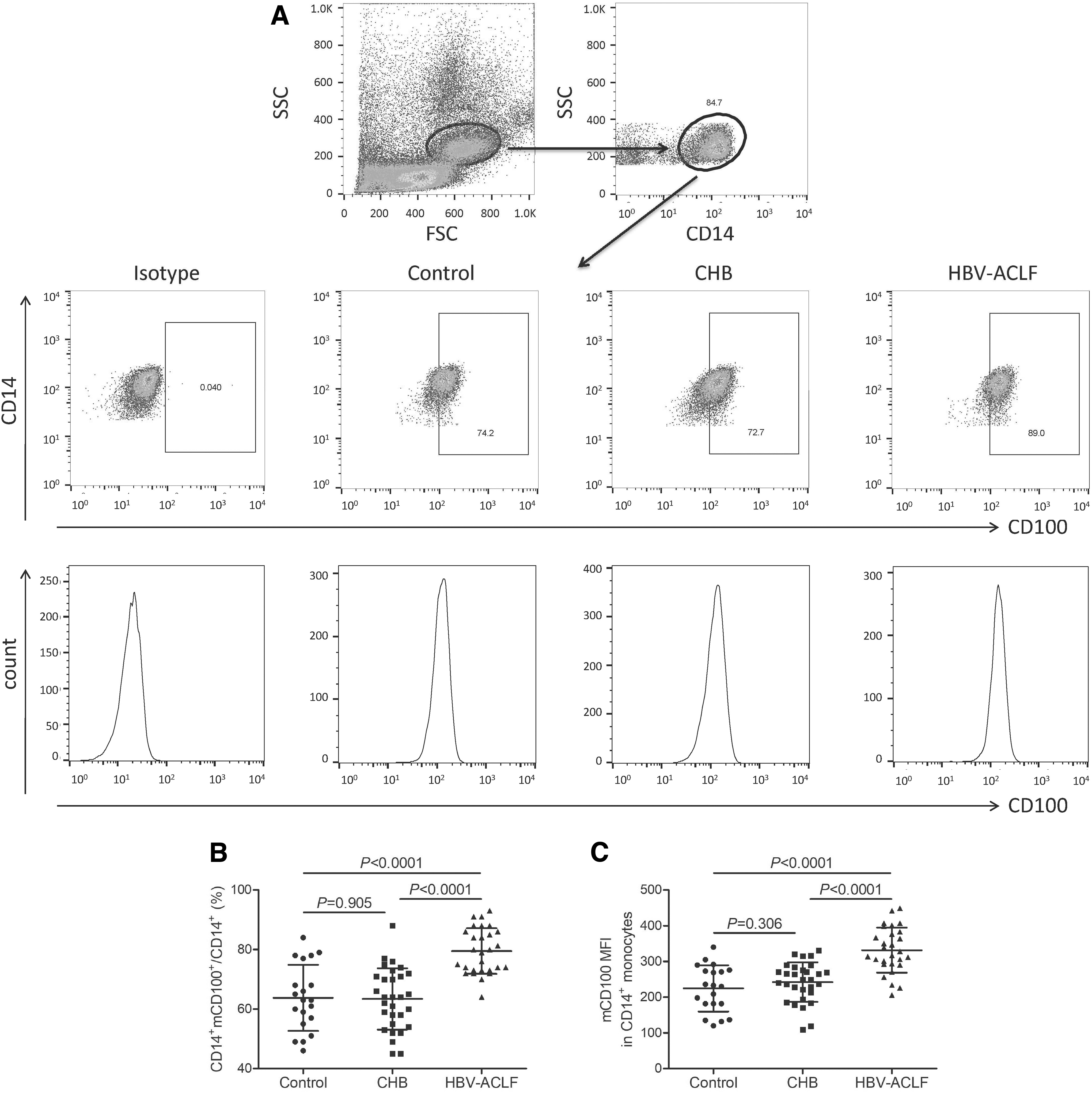
mCD100 in CD14+ monocytes was upregulated in HBV-ACLF patients. mCD100 expression in CD14+ monocytes was assessed by flow cytometry.
CD14+ monocyte function was exhausted in HBV-ACLF patients
Overall, 105 of purified CD14+ monocytes from 24 HBV-ACLF patients and 13 controls were cultured in direct contact or in Transwell-based indirect contact with 5 × 105 of Huh7.5 cells for 48 h. The percentage of HBV-ACLF-derived CD14+ induced target cell death was significantly decreased when compared with controls in both direct contact (13.95% ± 3.85% vs. 28.00% ± 5.43%, Student t test, p < 0.0001, Fig. 3A) and indirect contact systems (14.12% ± 3.88% vs. 25.15% ± 5.49%, Student t test, p < 0.0001, Fig. 3A). However, there were no remarkable differences in the percentage of target cell death between direct contact and indirect contact systems in either HBV-ACLF patients or controls (Student t tests, p = 0.879 and p = 0.196 respectively, Fig. 3A). IL-1β, IL-6, IFN-γ, TNF-α, and granzyme B level in the supernatant was measured by ELISA. There were no significant differences of IL-1β or IL-6 level between HBV-ACLF patients and controls or between direct and indirect contact systems (Student t tests, p > 0.05, Fig. 3B, C). Further, IFN-γ, TNF-α, and granzyme B level was notably decreased in HBV-ACLF patients when compared with controls in both direct contact and indirect contact systems (Student t tests, p < 0.05, Fig. 3D–F). Similarly, there were no statistically significant differences of IFN-γ, TNF-α, or granzyme B level between direct contact and indirect contact systems in either HBV-ACLF patients or controls (Student t tests, p > 0.05, Fig. 3D–F). The MFI corresponding to FasL and TRAIL within CD14+ monocytes was assessed by flow cytometry. Further, representative flow data for isotype control, FasL, and TRAIL in direct contact and indirect contact systems are shown in Figure 4A. There were no significant differences of FasL or TRAIL MFI in CD14+ monocytes between HBV-ACLF patients and controls or between direct and indirect contact systems (Student t tests, p > 0.05, Fig. 4B, C).
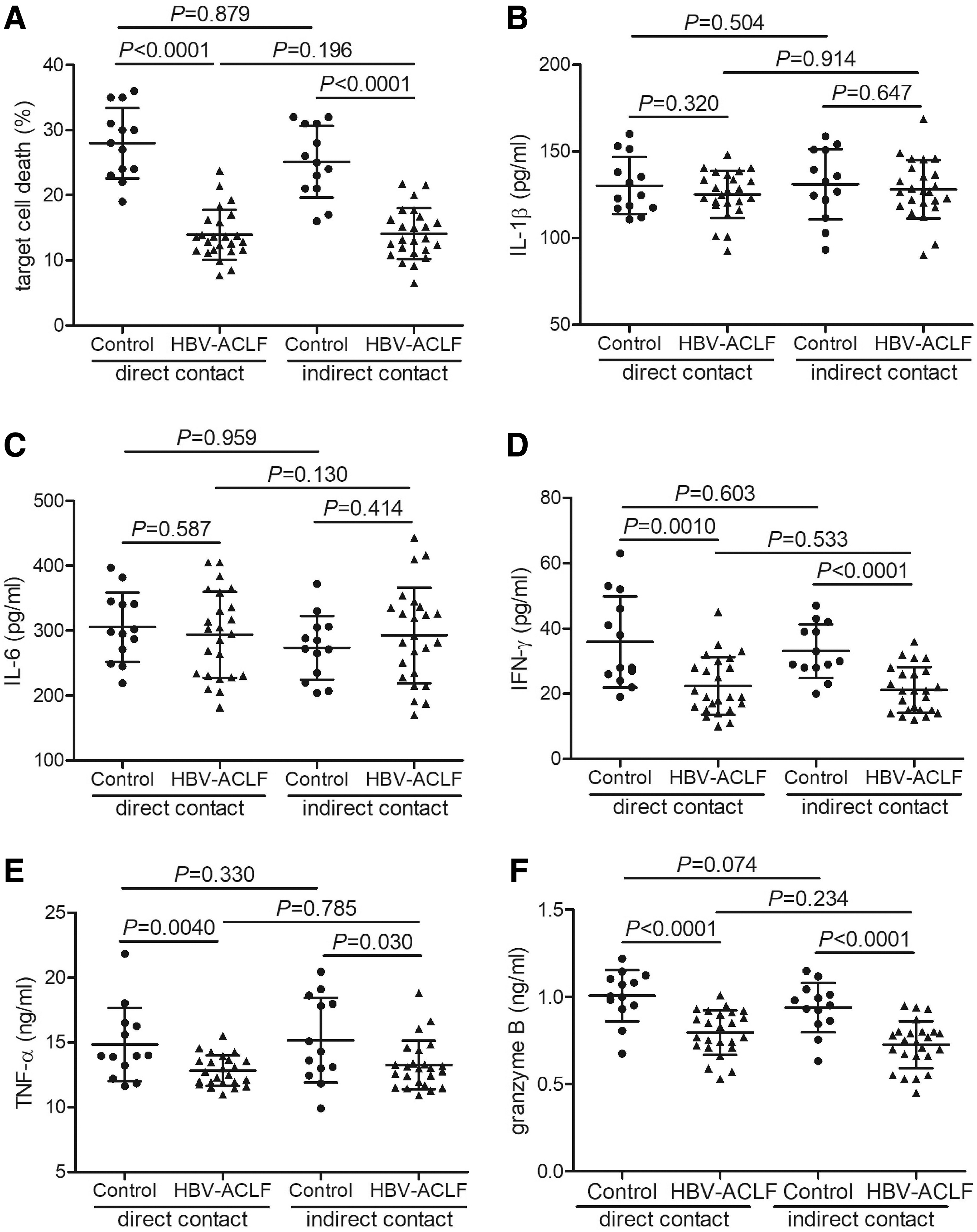
Target cell death, cytokine and granzyme B secretion by CD14+ monocytes was reduced in HBV-ACLF patients. CD14+ monocytes were purified from 24 HBV-ACLF patients and 13 healthy controls. CD14+ monocytes were stimulated with 1 × LPS for 12 h. Overall, 105 of CD14+ monocytes were cultured in a direct contact or in a Transwell-based indirect contact system with 5 × 105 of Huh7.5 cells for 48 h, and supernatants were harvested.
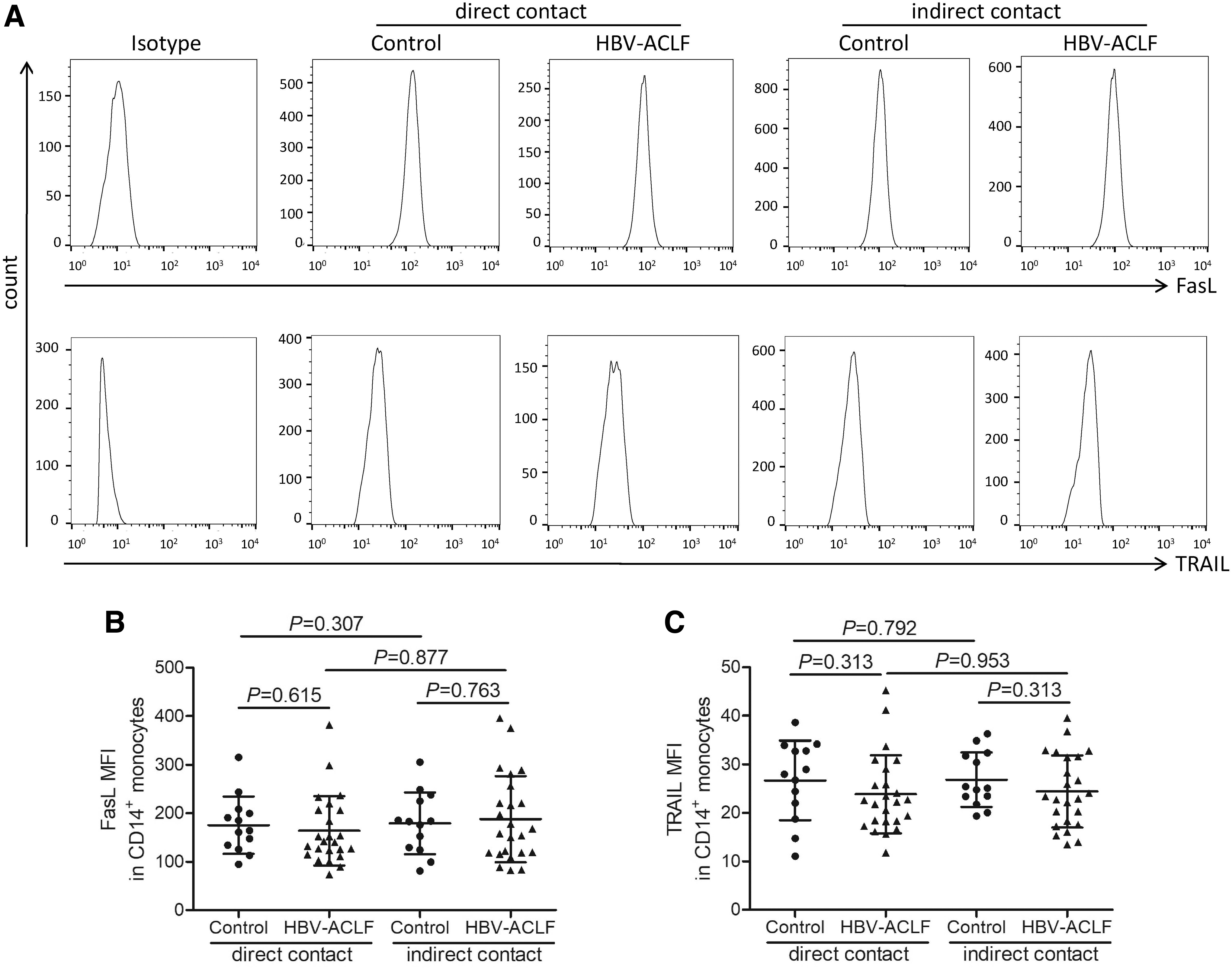
FasL and TRAIL MFI in CD14+ monocytes was comparable between HBV-ACLF patients and controls. CD14+ monocytes were purified from 24 HBV-ACLF patients and 13 healthy controls. CD14+ monocytes were stimulated with 1 × LPS for 12 h. Overall, 105 of CD14+ monocytes were cultured in a direct contact or a Transwell-based indirect contact system with 5 × 105 of Huh7.5 cells for 48 h, and cells were harvested. FasL and TRAIL level in CD14+ monocytes was assessed by flow cytometry.
Recombinant sCD100 enhanced CD14+ monocytes induced target cell death in HBV-ACLF patients
Purified CD14+ monocytes from 12 HBV-ACLF patients were stimulated with 1 × LPS in the presence or absence of recombinant sCD100 for 12 h. Overall, 105 of stimulated CD14+ monocytes were cultured in direct contact or in a Transwell-based indirect contact system with 5 × 105 of Huh7.5 cells for 48 h. The percentage of HBV-ACLF-derived CD14+ induced target cell death was remarkably increased in response to sCD100 stimulation in both direct contact (20.19% ± 4.42% vs. 13.48% ± 3.42%, paired t test, p = 0.0004, Fig. 5A) and indirect contact systems (18.15% ± 4.38% vs. 12.84% ± 2.26%, paired t test, p = 0.0012, Fig. 5A). There were no significant differences of IFN-γ or TNF-α level secreted by CD14+ monocytes between without CD100 and with CD100 stimulation or between direct and indirect contact systems (paired t tests and Student t tests, p > 0.05, Fig. 5B, C). However, granzyme B level was notably elevated in response to sCD100 stimulation in both direct contact (1.23 ± 0.15 ng/mL vs. 0.77 ± 0.15 ng/mL, paired t test, p < 0.0001, Fig. 5D) and indirect contact systems (1.19 ± 0.23 ng/mL vs. 0.76 ± 0.13 ng/mL, paired t test, p = 0.0012, Fig. 5D).
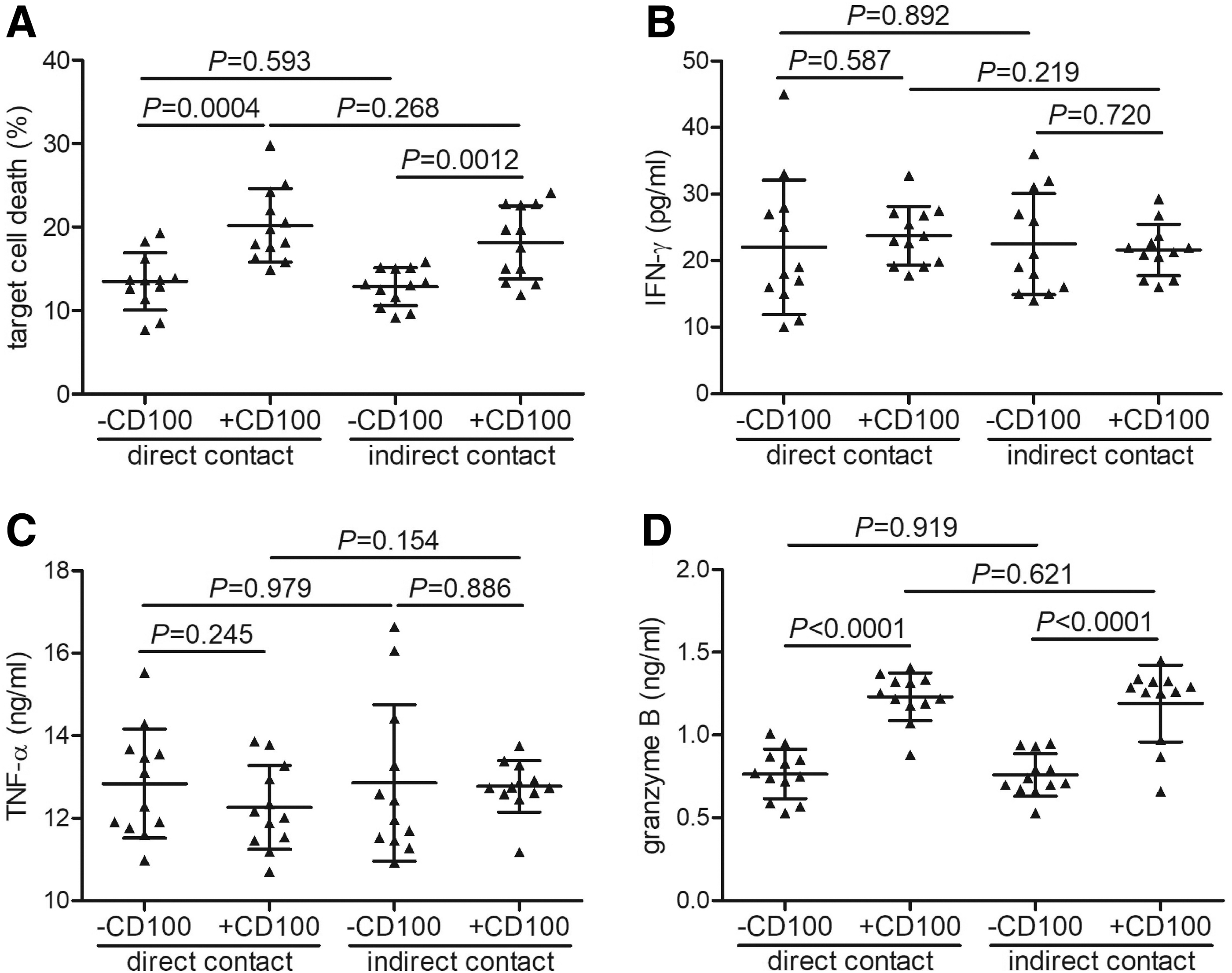
Target cell death, cytokine and granzyme B secretion by CD14+ monocytes was elevated in response to recombinant sCD100 stimulation in HBV-ACLF patients. CD14+ monocytes were purified from 12 HBV-ACLF patients, and they were stimulated with 1 × LPS in the presence or absence of recombinant sCD100 for 12 h. Overall, 105 of stimulated CD14+ monocytes were cultured in a direct contact or in a Transwell-based indirect contact system with 5 × 105 of Huh7.5 cells for 48 h, and supernatants were harvested.
Recombinant sCD100 enhanced CD14+ monocytes induced CD4+/CD8+ T cell activation in HBV-ACLF patients
Purified CD14+ monocytes from 10 HBV-ACLF patients were stimulated with 1 × LPS in the presence or absence of recombinant sCD100 for 12 h. Overall, 105 of stimulated CD14+ monocytes were in a direct contact system with autogolous 105 of CD3+ T cells for 48 h. IFN-γ and IL-17 secretion by CD3+CD4+ T cells and IFN-γ and TNF-α secretion by CD3+CD8+ T cells was measured by flow cytometry. The representative flow dots for isotype control and intracellular staining were shown. Recombinant sCD100 stimulation increased the percentage of CD4+IFN-γ + cells (1.49% ± 0.48% vs. 1.06% ± 0.16%, paired t test, p = 0.018, Fig. 6A) and CD4+IL-17+ cells (1.02% ± 0.17% vs. 0.74% ± 0.14%, paired t test, p = 0.0007, Fig. 6B). Similarly, recombinant sCD100 stimulation also elevated the percentage of CD8+IFN-γ + cells (1.73% ± 0.36% vs. 1.13% ± 0.11%, paired t test, p < 0.0001, Fig. 6C) and CD8+TNF-α + cells (5.10% ± 1.13% vs. 2.92% ± 0.59%, paired t test, p < 0.0001, Fig. 6D).
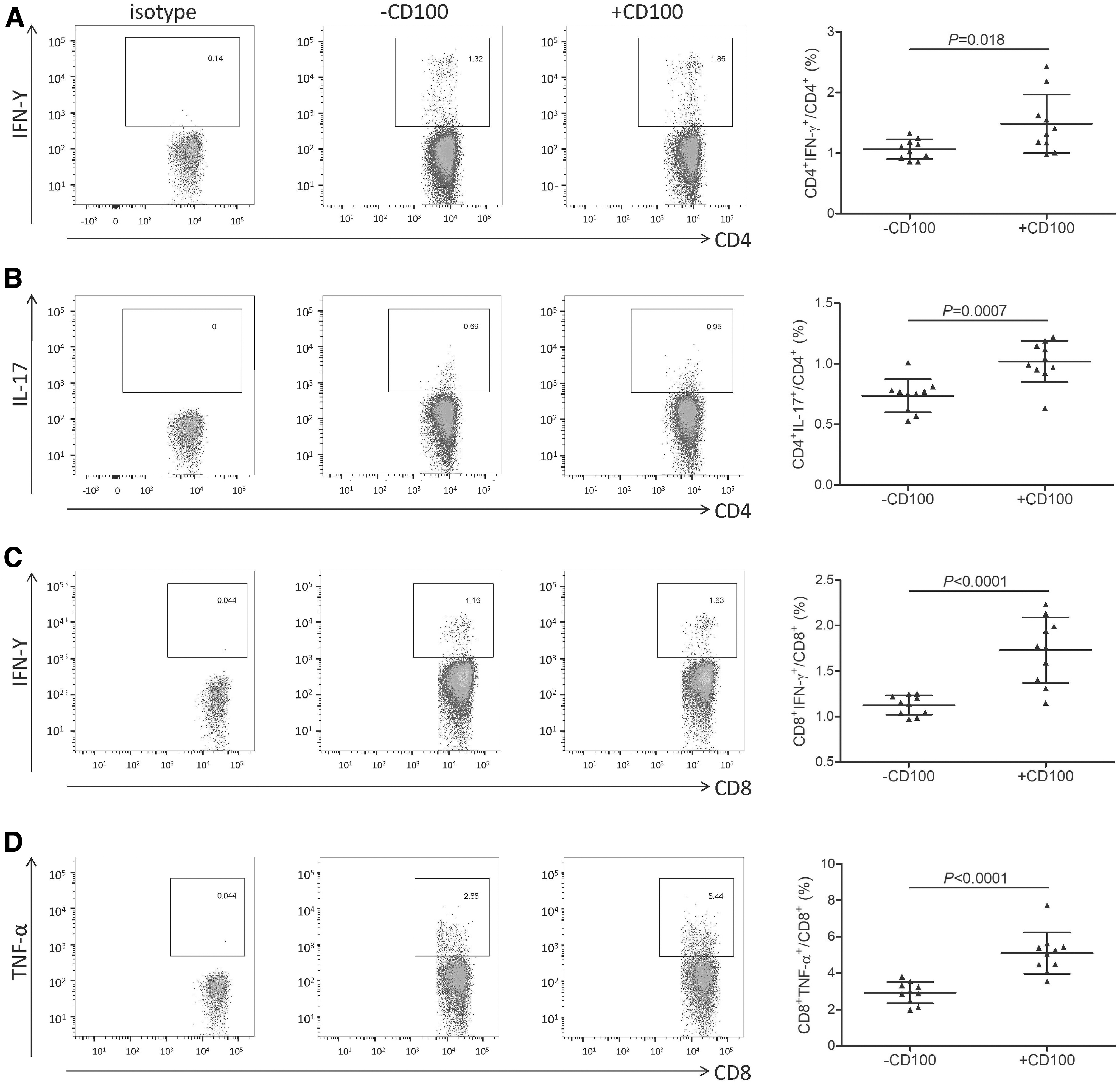
CD14+ monocyte-induced CD4+/CD8+ T cell activation was elevated in response to recombinant sCD100 stimulation in HBV-ACLF patients. CD14+ monocytes were purified from 10 HBV-ACLF patients, and they were stimulated with 1 × LPS in the presence or absence of recombinant sCD100 for 12 h. Overall, 105 of stimulated CD14+ monocytes were in direct contact with autologous 105 of CD3+ T cells for 48 h. IFN-γ and IL-17 secretion by CD3+CD4+ T cells and IFN-γ and TNF-α secretion by CD3+CD8+ T cells was assessed by flow cytometry. The representative flow dots for isotype and for intracellular staining were shown.
Discussion
To the best of our knowledge, this was the first article on sCD100/mCD100 balance and its regulation to CD14+ monocytes in HBV-ACLF patients. HBV-ACLF patients had lower peripheral sCD100 level and higher mCD100 expression on CD14+ monocytes even compared with chronic HBV-infected patients. HBV-ACLF-derived peripheral CD14+ monocytes revealed exhausted phenotype, which presented as reduced cytotoxicity to target cells and abatement of IFN-γ, TNF-α, and granzyme B secretion. Importantly, recombinant sCD100 stimulation enhanced CD14+ monocyte-induced cell death and CD14+ monocyte-mediated T cell activation in HBV-ACLF patients. The current results indicated that there was imbalance between sCD100 and mCD100 on CD14+ monocytes in HBV-ACLF patients, leading to the inactivation of innate immune response and being beneficial for the survival of HBV-ACLF patients.
CD100 contributed to the regulation of immune response in viral infections. However, the expression profile was varied in acute and chronic viral infectious diseases. A higher proteolytic release of sCD100 from CD4+ T cells was found in patients with neuronal human lymphotropic virus type 1 infection, and signaling through sCD100 and its receptor could activate axonal growth collapse pathways, resulting in neurodegenerative and inflammatory response in the central nervous system (20). The elevated plasma sCD100 was reported in patients with hemorrhagic fever with renal syndrome (HFRS), and it was associated with disease severity (15). Further, elevation of sCD100 in HFRS patients might be a result from the shedding of mCD100 on PBMCs (15). The shedding progression could lead to a novel CD8+ T cell subset, CD8lowCD100− T cells, which might be associated with viral clearance during Hantaan virus infection (16). Thus, acute viral infection always leads to elevated shedding of mCD100 from immune cells and increased sCD100 level, which was important for viral control and inflammation (18). In contrast, the immune status of chronic viral infection was completely different from acute infection. Serum sCD100 level was decreased whereas CD100 expression was increased on T cells and B cells in human immunodeficiency virus-1 (HIV-1) positive viremic individuals (5,23). The insufficient mCD100 shedding might contribute to the exhaustion of T cells, B cells, and NK cells in chronic HIV and HCV infections (5 –7). Importantly, CHB patients had higher mCD100 on T cells and lower sCD100 in serum (27). Both therapeutic sCD100 treatment and matrix metalloproteases-mediated sCD100 release revealed an important role in the regulation of intrahepatic anti-HBV CD8+ T cells responses (27). The current results showed no significant difference of sCD100/mCD100 between controls and ASC. This indicated that HBV infection itself might not be the main reason for the imbalance between sCD100 and mCD100 due to the high level of HBV DNA in ASC. Further, our current data also revealed a significant increase in peripheral sCD100 level and a decrease in mCD100 level on peripheral CD14+ monocytes in HBV-ACLF patients. sCD100 level in HBV-ACLF patients was even lower compared with CHB patients. In contrast, mCD100 level on CD14+ monocytes was comparable between CHB and HBV-ACLF patients. This indicated that sCD100 might derive mainly from other immune cells, including T cells, B cells, and NK cells, in CHB patients. Moreover, sCD100 downregulation in HBV-ACLF patients might be due to the insufficient shedding from immune cells, and elevated mCD100 on CD14+ monocytes might also take part in the regulation of innate immune responses in HBV-associated ACLF. Interestingly, the HBV-ACLF group was, indeed, older than other three groups, revealing that age might also relate to the reduced immune responses. Taken together, further experiments for mCD100 staining combined with immune check point molecule intracellular staining should be performed to explain whether monocytes with higher mCD100 expression showed an exhausted phenotype, whereas the role of immunoregulatory activity of sCD100 in HBV-related ACLF is still needed for further elucidation.
CD14+ monocytes not only play a crucial cytotoxic role in innate immune response, but they also serve as important antigen-presenting cells for T cell activation. CD14+ monocytes reveal direct cytotoxic function to pathogen-infected cells and tumor via two different pathways (26). On the one hand, CD14+ monocytes produced proinflammatory cytokines (mainly IL-1β, IL-6, IFN-γ, and TNF-α) and cytotoxic molecules (mainly granzymes), leading to the necrosis and apoptosis of target cells (14,29). On the other hand, surface expression of FasL and TRAIL on CD14+ monocytes also mediated the elimination of compromised cells (11,21). We found that CD14+ monocytes from HBV-ACLF patients revealed dysfunctional and exhausted phenotype due to reduced cytotoxicity to target cells in vitro. Importantly, IFN-γ, TNF-α, and granzyme B production was decreased in both direct and indirect contact systems; however, IL-1β/IL-6 secretion by CD14+ monocytes, or FasL/TRAIL expression on CD14+ monocytes was comparable between HBV-ACLF patients and healthy individuals. This indicated that certain proinflammatory cytokines and cytotoxic molecules, but not FasL or TRAIL-associated signaling pathways, might be impaired in HBV-related ACLF. Further, there were no remarkable differences of secretion of IFN-γ, TNF-α, or granzyme B between direct and indirect contact systems in both HBV-ACLF patients and controls, suggesting that CD14+ monocyte-induced target cell death was independent of direct cell-to-cell contact. Taken together, impaired proinflammatory cytokines and cytotoxic molecules might contribute to CD14+ monocyte exhaustion in HBV-ACLF patients. CD14+ monocyte restoration might be one of the novel therapeutic strategies for HBV-ACLF treatment.
sCD100 enhanced NK cells, dendritic cells, CD4+ T cells, and CD8+ T cells activation in allergy (30), autoimmune disorders (9,13), cancers (24), and chronic viral infections (7,12,27), However, a few studies focused on the regulatory function of sCD100 to CD14+ monocytes. The present data showed that recombinant sCD100 stimulation not only enhanced CD14+ monocytes cytotoxicity to target cells, but it also promoted antigen-presenting activity of CD14+ monocytes for T cell activation in HBV-ACLF patients. sCD100 stimulation only elevated the secretion of granzyme B by CD14+ monocytes in both direct and indirect contact systems. This was partly consistent with the recent published results (26), and it revealed the pivotal role of granzyme B in the cytotoxicity of CD14+ monocytes. Moreover, sCD100 stimulation effectively promoted the CD14+ monocytes-mediated activation and differentiation of IFN-γ-producing Th1 and IL-17-producing Th17 cells, as well as IFN-γ-producing CD8+ T and TNF-α-producing CD8+ T cells under direct contact condition in HBV-associated ACLF. Importantly, induction of inflammatory cytokines in T cells cocultured with sCD100-stimulated monocytes was antigen nonspecific. Thus, there still remained questions regarding which signal is important for this T cell activation and which responses are independent of mCD100 of T cells themselves. Further experiments are also needed for MHC-class II, CD80, and CD86 staining. Previous reports have been demonstrated that various cytokines (14,26) and Notch signaling (29) directly modulated the antigen-presenting activity of CD14+ monocytes. sCD100 also enhanced CD14+ monocyte-induced T cell activation in HBV-ACLF, suggesting that low plasma sCD100 level might be insufficient for either CD14+ monocyte cytotoxicity or adaptive immune response in HBV-ACLF.
In summary, decreased peripheral sCD100 might contribute to CD14+ monocyte inactivation in HBV-ACLF patients. sCD100 showed important immunomodulatory activity to CD14+ monocytes, including granzyme B-induced cytotoxicity and direct cell-to-cell contact-mediated T cell activation during HBV-ACLF. The current data suggested that severe inflammation induced sCD100/mCD100 imbalance to inactivate CD14+ monocyte response, which might be beneficial for the survival of HBV-ACLF patients.
Footnotes
Acknowledgment
The authors thank Dr. Hao Zuo from The Second Hospital of Jilin University for the helpful discussion and critical revision of the article.
Author Disclosure Statement
No competing financial interests exist.