
Abstract
Hepatitis B is a viral infection that can cause serious liver disease. Chronic hepatitis B (CHB) infection places individuals at higher risk of developing cirrhosis of the liver and hepatocellular cancer. Immune dysfunction, including altered distribution and functional status of T cell immunity, is a contributor to hepatitis B virus (HBV) pathogenesis. In this study, we examined the distribution of circulating γδ T cell subpopulations and levels of cell surface expression of suppressive markers on γδ T cells in individuals with CHB infection and clinical liver disease. A significantly higher proportion of terminally differentiated (TEMRA) (CD27−CD45RA+) γδ T cells along with significantly lower percentages of central memory (CD27+CD45RA−) and effector memory (CD27−CD45RA−) γδ T cells were observed in peripheral blood of these individuals. The expression of exhaustion markers-Tim-3 and Lag-3 was elevated in γδ T cells from CHB-infected individuals compared with healthy controls (HC) and blockade of these exhaustion markers resulted in restoration of interferon gamma (IFN-γ) secretion by γδ T cells. In addition, γδ T cells from CHB patients expressed increased levels of CD69, another important regulator of immune responses. Together, these results suggest that CHB patients with clinical sign of liver disease have TEMRA γδ T cells with a potentially exhausted phenotype that may in turn impair their immunoregulatory role and facilitate pathogenesis of CHB disease.
Introduction
Hepatitis B virus (HBV) infection is a major global health concern with an estimated 2 billion people exposed to the virus. Also, 80% cases of primary hepatic cancer are due to chronic hepatitis B virus (CHB) (19,24). HBV-induced hepatic cancer arises almost exclusively due to the chronically inflamed liver. The liver is enriched with immune cells that can mediate an effective immune response against pathogens. Under basal condition, the microenvironment of the liver is immunosuppressive, but the pathogenic assault accompanying HBV infection causes activation of intrahepatic as well as peripheral immune cells. When protective, efficient innate and adaptive immune responses are sufficient to protect the host in self-limited acute HBV infection.
However, cellular immunity mediated by CD4+ T cells, CD8+ T cells, and natural killer cells along with inflammatory cytokines, while clearing the virus, contribute to liver damage (11,28,29). The potential liver damage caused by these antiviral response results in leakage of liver enzymes, such as alanine aminotransferase (ALT) and aspartate aminotransferase (AST) into the blood stream. Under chronic HBV settings, prolonged stimulation of immune cells causes immune-mediated hepatocyte damage that in turn contributes to disturbance in immune-hepatocyte homeostasis predisposing patients to the development of hepatocellular carcinoma.
Although immunosuppressive cells, such as regulatory T cells (Tregs) and myeloid-derived suppressor cells (MDSCs) play important roles in dousing the inflammatory responses in CHB patients, they may also contribute to T cell dysfunction (7). A characteristic feature of dysfunctional T cell is the expression of exhaustion markers (e.g., Tim-3, Lag-3, CTLA-4, and PD-1) that limit effector function. The expression of exhaustion markers or inhibitory receptors on T cells is a gradual process, which is accompanied by loss of cytokine production, inefficient cytolytic potential, diminished proliferative response, and thereby, viral persistence. Therefore, an appropriate homeostatic balance between host immune cells and hepatocytes is essential to check HBV without eliciting a nonresolving inflammation.
The frequency of γδ T cells have been reported to be enriched in the liver during both infectious or noninfectious inflammatory settings (13,15,25). It has also been reported that γδ T cells play a regulatory role in maintaining a tolerizing environment in the liver by inhibiting the secretion of TH17 cytokines and mobilizing infiltration of MDSC to the liver (16,27). Notably, earlier studies have demonstrated that the functional capacity of γδ T cells is significantly decreased in CHB patients with strong inflammatory response in the liver (6,27). We, therefore, postulate that γδ T cells could become dysfunctional in CHB patients with clinical symptoms of abnormal liver function, which in turn would contribute to disrupting immune-hepatocyte homeostasis and consequently, liver injury.
In this study, we examined the proportion of γδ T cells and their subpopulations after CHB infection. Next, the expression of exhaustion and activation markers was also measured to evaluate the possible involvement of γδ T cells in CHB pathology.
Materials and Methods
Subjects
Blood samples were collected from 35 chronic HBV (CHB) patients with symptoms of liver disease and 30 healthy volunteer controls (HC) on written informed consent. CHB patients were diagnosed based on hepatitis B surface antigen (HBsAg) positivity for ≥6 months and with viral load of 5.4 × 103 to 8.1 × 107 copies/mL. Besides, the patients were having elevated alanine aminotransferase (ALT) and aspartate aminotransferase (AST) with some clinical symptoms. Peripheral blood mononuclear cells (PBMCs) from CHB and HC were isolated by Ficoll-Hypaque (Sigma-Aldrich, St. Louis, MO) density gradient centrifugation. Cells were suspended either in fluorescence activated cell sorting (FACS) buffer (0.01 M phosphate buffered saline (PBS) pH-7.4, 1% fetal calf serum (FCS), 0.01% sodium azide) or in Roswell Park Memorial Institute (RPMI) 1640 medium supplemented with 10% heat-inactivated human AB serum plus rIL-2 (100 U/mL), 2 mM glutamine, and antibiotics for subsequent use.
Ethics statement
The study was approved by the Institutional Ethics Committee of the Regional Medical Research Centre, Northeast Region, Dibrugarh.
Virological assessment
The HBV viral load was quantified using the High Pure extraction system and HBV test kit (Roche Diagnostics Corporation, Mannheim, Germany) in Cobas TaqMan 48 Analyzer (Rotkreuz, Switzerland). The assay has a limit of detection 6–10,000,000 IU/mL where 1 IU/mL correspond to 5.8 copies/mL. HBsAg and hepatitis B virus e antigen (HbeAg) detection in clinical specimens were done by using enzyme-linked immunosorbent assay (ELISA) kit, as per the manufacturer's instruction (BioRad, Hercules, CA; DRG International, Inc., Springfield, NJ).
Flow cytometry
Isolated PBMCs were stained for surface markers with the following monoclonal antibodies: anti-CD3-PE (Clone: UCHT1, 0.5 μg/mL), anti-γδ TCR-FITC (Clone: B1, 2.5 μg/mL), anti-CD27-PE (Clone: M-T271, 0.5 μg/mL), anti-CD45RA-PECy5 (Clone: HI100, 1 μg/mL), anti-γδ TCR-PE (Clone: B1, 2.5 μg/mL), anti-CD69-FITC (Clone: FN50, 2.5 μg/mL), anti-CD25-FITC (Clone: M-A251, 2.5 μg/mL) (BD Bioscience, San Diego, CA), anti-Tim-3-PE (Clone: AC145, 5 μg/mL), and anti-Lag-3-PE (Clone: REA351, 5 μg/mL) antibodies (Miltenyi Biotec, Bergish Gladbach, Germany). The required antibody or combination of antibodies were diluted in FACS buffer (0.01 M PBS pH-7.4, 1% FCS, 0.01% sodium azide) with an end volume of 50 μL per test and samples were stained with required antibodies for 45 min at 4°C. Subsequently, cells were washed with FACS buffer and intensity of fluorescence was measured by using flow cytometer (single laser, 5-colour FC500 with CXP software; Beckton Dickinson). The cells were gated on the basis of their forward and side scatter characteristics and the fluorescence intensity was assessed. Data obtained were analyzed using FlowJo vX.0.7 (Tree Star) software.
Functional assay
PBMCs were stimulated with PMA (phorbol 12-myristate 13-acetate, 50 ng/mL) and ionomycin (1 μg/mL) for 4 h in the presence of Brefeldin A (5 μg/mL; all from Sigma Aldrich). For blocking assay, cells were pretreated for 30 min at 37°C with or without either IgG-Fc control protein or Tim-3-Fc protein and Lag-3-Fc protein (R&D Systems, Minneapolis, MN). Cells were washed with PBS, followed by fixation with 1% cold paraformaldehyde for 15 min at 4°C, and permeabilized for 5 min with 0.1% saponin (Sigma Aldrich). Next, cells were incubated with antibodies for anti-γδ TCR-PE (Clone: B1, 2.5 μg/mL) and anti-interferon gamma (IFN-γ)-FITC (Clone: 4S.B3, 5 μg/mL) (BD Bioscience) for 30 min at room temperature, and then washed with saponin buffer. Thereafter, cells were acquired using flow cytometer (FC500; Beckton Dickinson) for analyzing. Analysis was carried out using FlowJo vX.0.7 (TreeStar) software.
Statistical analysis
Statistical analyses have been performed using GraphPad Prism version 6.0. The nonparametric Mann–Whitney test was used to compare the groups and p-values <0.05 were considered significant. Spearman rank correlation was used for correlation analysis.
Results
Changes in circulating γδ T cell subpopulations during CHB infection
Peripheral blood from CHB patients and HC (Table 1) was subjected to flow cytometric analysis to determine the percentage of γδ T cells and their subpopulations. The representative data showing percentage of γδ T cells in healthy and CHB subjects is depicted in Figure 1A. There was no significant difference between the percentages of circulating γδ T cells in the healthy individuals and CHB patients (Fig. 1B). The frequencies of γδ T cells showed no correlation with HBV viral load (r = −0.36; Supplementary Fig. S1). Based on the relative surface expression of CD27 and CD45RA molecules, γδ T cells can be subdivided into four subpopulations: naive (N), central memory (CM), effector memory (EM), and terminally differentiated (TEMRA) cells (10). Accordingly, γδ T cells were further gated and the expression of CD27 and CD45RA surface markers was determined. The percentages of various γδ T cell subpopulations were altered among HC and CHB patients (Fig. 1C); fluorescence minus one controls are shown in Supplementary Figure S2A. A significant decrease in the percentage of CM (CD27+CD45RA−; mean percentage ± standard deviation [SD] was 15.9 ± 13.9) and EM (CD27−CD45RA−; mean percentage ± SD was 21.0 ± 16.1) γδ T cells were observed in patients with CHB compared with HC (p < 0.005); the mean percentages ± SD for CM and EM were 30.1 ± 15.0 and 34.3 ± 14.8, respectively (Fig. 1D). Moreover, the percentage of TEMRA (CD27−CD45RA+; mean percentage ± SD was 45.7 ± 20.8) within the γδ T cell fraction was significantly increased in CHB patients as compared with that in HC (24.3 ± 19.9, p < 0.005) (Fig. 1D). The data suggest that although the percentage of total γδ T cells was unaltered in CHB patients, the proportion of effector γδ T cells (CM as well as EM) was decreased with a corresponding increase in frequency of TEMRA observed.
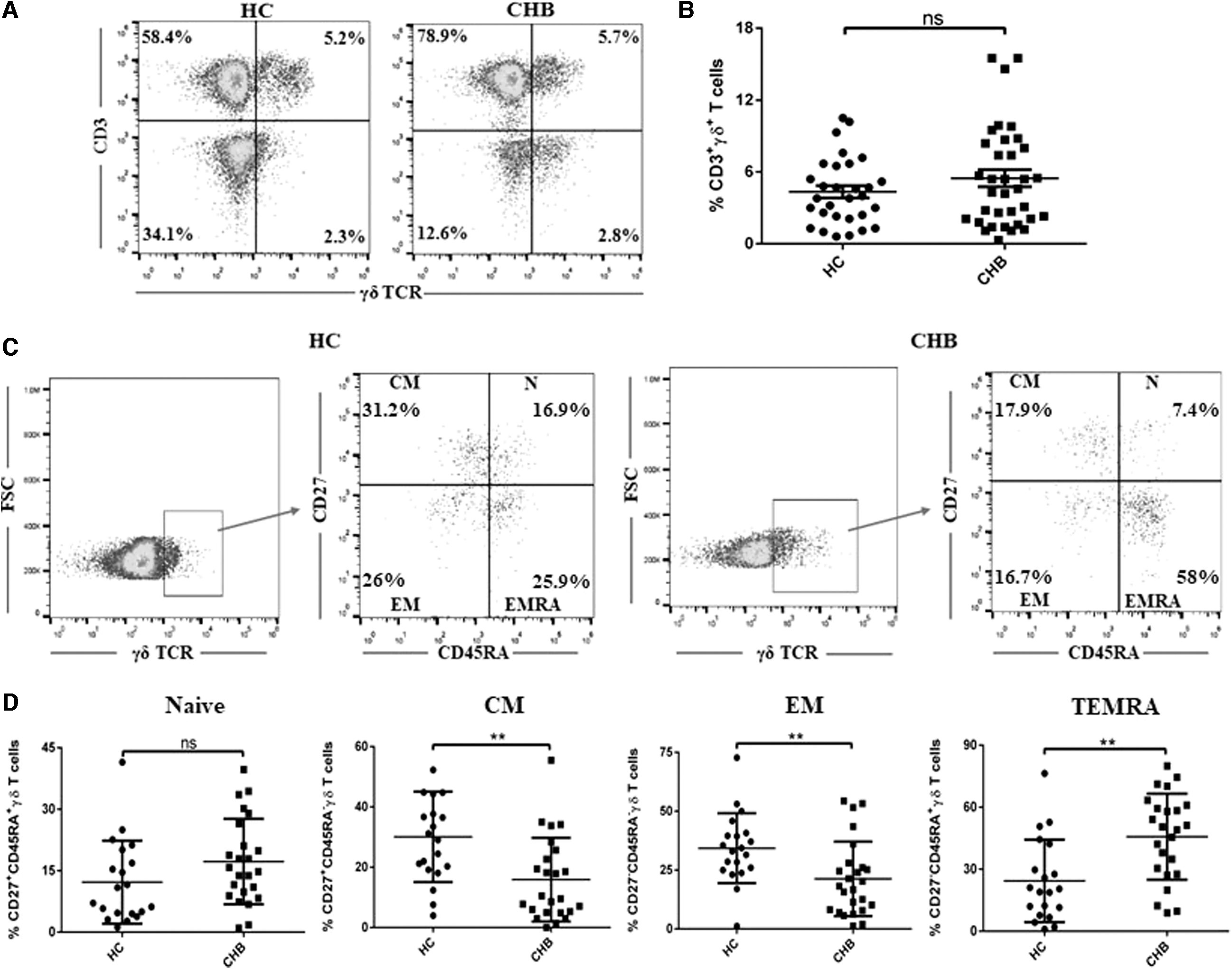
Determination of the frequencies of γδ TCR and γδ T cell subpopulations in CHB patients and HC.
Clinical Characteristics of the Study Groups
ALT, alanine aminotransferase; AST, aspartate aminotransferase; CHB, chronic hepatitis B; HBeAg, hepatitis B virus e antigen; HBV, hepatitis B virus; HC, healthy control; NA, not applicable.
Increased expression of Tim-3 and Lag-3 on γδ T cells from patients with CHB
The expression of Tim-3 and Lag-3 on γδ T cells was assessed in HC and CHB patients (Supplementary Fig. S2B). Figure 2A and Supplementary Figure S3 illustrate the expression of exhaustion markers-Tim-3 and Lag-3 within the γδ T cells (γδ+ TCR) gated population in both HC and CHB patients. Higher expression of Tim-3 and Lag-3 (mean percentage ± SD for Tim-3 and Lag-3 were 13.6 ± 11.44 and 4.8 ± 5.4, respectively) on γδ T cells was observed during CHB infection compared with that observed for HC (mean percentage ± SD for Tim-3 and Lag-3 were 7.5 ± 7.7, *p < 0.05 and 2.2 ± 2.5, **p < 0.005, respectively) (Fig. 2B). To assess whether expression of Tim-3 and Lag-3 inhibits IFN-γ secretion by γδ T cells, we examined the effects of blocking the Tim-3 and Lag-3 receptor on peripheral blood from patients with CHB. PBMCs isolated from CHB patients and HC were stimulated with PMA/ionomycin in the presence or absence of Tim-3-Fc protein and Lag-3-Fc protein. Stimulation of PBMCs with PMA/ionomycin led to expression of IFN-γ in γδ T cells, evaluated by intracellular cytokine staining, while the median fluorescence intensity of IFN-γ + γδ T cells was downregulated in CHB patients compared with HC (Supplementary Fig. S4A). The amount of IFN-γ in γδ T cells increased when PBMCs from patients with CHB were treated with Tim-3-Fc protein and Lag-3-Fc protein (Fig. 2C and Supplementary Fig. S4B). These findings demonstrate the existence of Tim-3 and Lag-3 mediated exhaustion of γδ T cells during CHB infection.
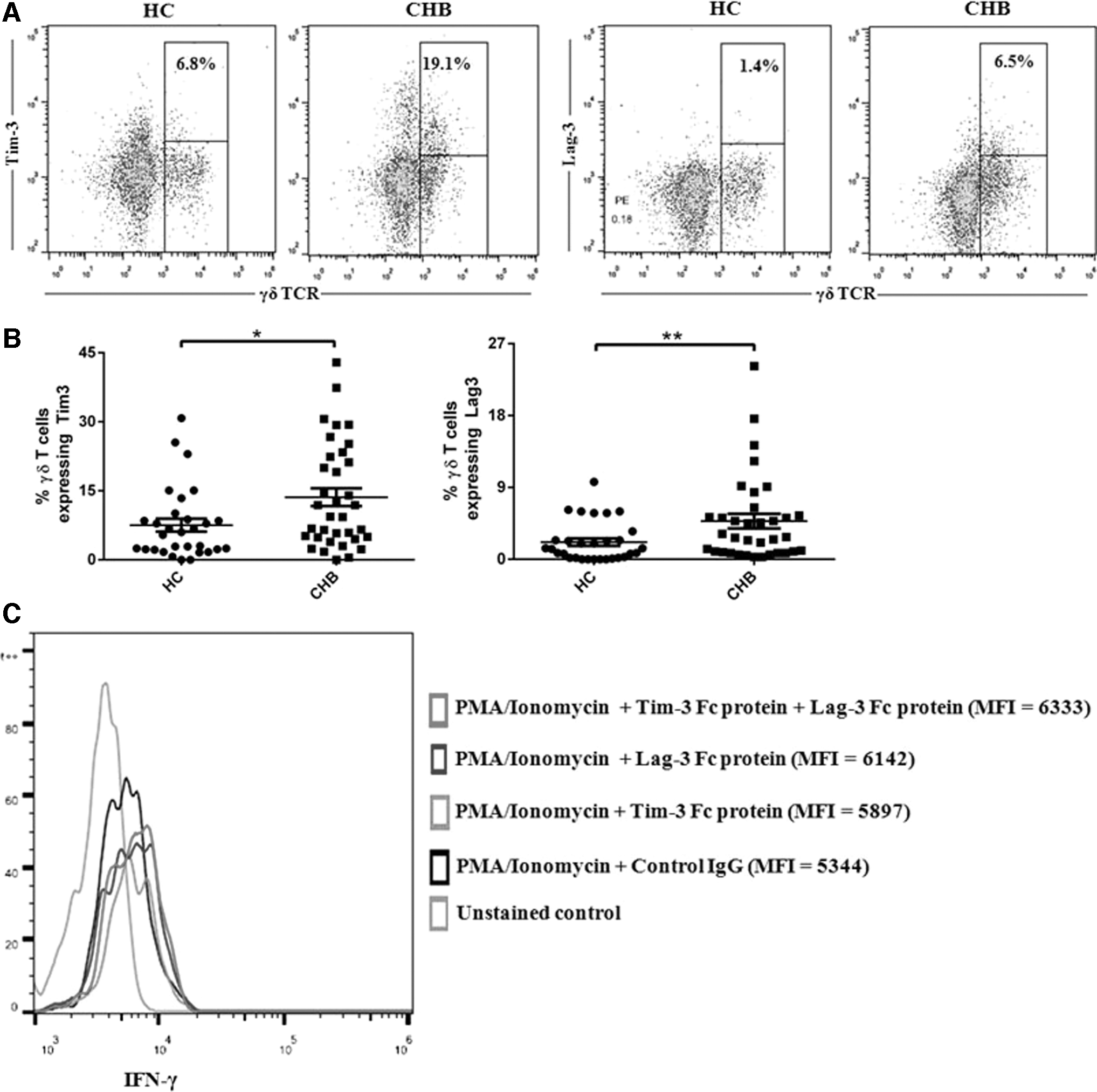
Upregulated expression of Tim-3 and Lag-3 on γδ T cells during CHB infection.
Upregulation of CD69 expression on γδ T cells isolated from CHB patients
CD69 is an important regulator of immune responses and has widely been used as an early activation marker, but recent evidence suggests that CD69 also induces exhaustion of tumor infiltrating lymphocytes in a murine model (20). The expression of both early and late activation markers, CD69 and CD25, respectively, was examined on γδ T cells from patients with CHB. Figure 3A contains representative plots showing the proportion of CD69 and CD25, respectively, within the γδ T cells-gated population in HC and CHB individuals. A marked increase in the percentage of CD69 on γδ T cells (mean percentage ± SD for CD69 was 9.3 ± 6.5) compared with that of HC (mean percentage ± SD for CD69 was 4.1 ± 3.9, **p < 0.005) was observed (Fig. 3B). However, there was no significant difference in the expression of CD25 on γδ T cells in CHB patients compared with HC (Fig. 3B). These data indicate that chronic infection with HBV results in the upregulation of CD69 on γδ T cells.
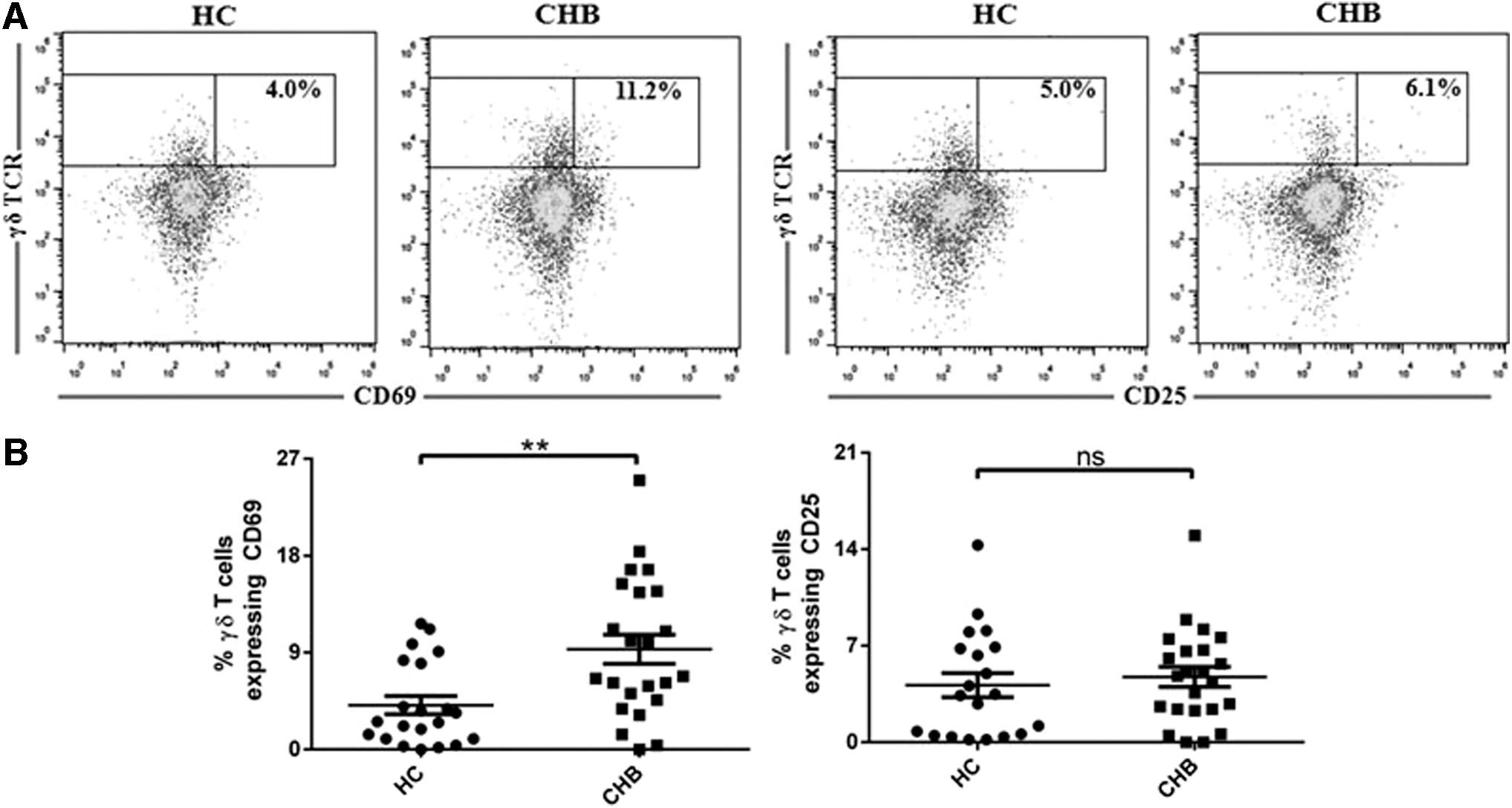
Increased expression of CD69 on γδ T cells from CHB patients.
Discussion
Hepatitis B is a serious health problem with an estimated 350 million people worldwide suffering from CHB (17). CHB is characterized by impaired CD4+ and CD8+ T cell response, including clonal depletion and T cell exhaustion (1,7). However, it is currently poorly understood how γδ T cells are affected during CHB infection with clinical sign of liver disease. Our data demonstrate that chronic exposure to HBV did not change the proportion of circulating γδ T cells compared with HC. This result is in line with a recent study with a North American cohort showing that the frequency of γδ T cells did not differ between CHB and healthy subjects (5).
Staining for CD27 and CD45RA cell surface markers permits identification of four subpopulations of γδ T cell: N (CD27+CD45RA+), CM (CD27+CD45RA−), EM (CD27−CD45RA−), and terminal differentiated (CD27−CD45RA+) (10). Peripheral blood CD27+CD45RA− and CD27−CD45RA− γδ T cell subpopulations have been reported to be involved in IFN-γ and cytolytic effector functions of γδ T cells, whereas CD27−CD45RA+ represent TEMRA γδ T cells (4,23). We, therefore, analyzed the percentage of different subpopulations of γδ T cells in CHB patients to evaluate functional profiles of these cells. A significant decrease in circulating frequency of CM (CD27+CD45RA−) and EM (CD27−CD45RA−) γδ T cells was found in CHB patients compared with HC. We also observed an increase in the percentage of TEMRA (CD27−CD45RA+) γδ T cells in CHB individuals compared with HC. These results demonstrated that γδ T cells during CHB infection might have impaired effector functions and undergo expansion to the terminal stage of differentiation.
The expression of exhaustion markers during CHB has received attention for their role in reducing effector functions of T cells. During CHB infection, αβ T cells that include CD4+ and CD8+ T cells have been shown to have an unregulated expression of exhaustion markers, such as Tim-3, Lag-3, and PD-1, resulting in the impaired effector function of these cells (2,12,21,22). The antiviral αβ T cell response of CHB patients has been suggested to be impaired through Tim-3/galectin-9 pathway (21). Furthermore, the presence of Lag-3 and PD-1 on CD4+ T cells during CHB infection has been shown to inhibit production of Th1 cytokines (12). Similarly, high levels of PD-1 on CD8+ T cells in patients with CHB are reported to correlate with repressed IFN-γ and IL2 production (3,26).
Although, cytotoxic T cell effector machinery is required to clear HBV-infected hepatocytes, this is achieved, however, at the cost of destroying hepatocytes (14,26). Both the control of HBV infection and liver injury are dependent on the interplay of the host immune system and hepatocyte damage. It thus seems reasonable to believe that induction of tolerance by the process of immune exhaustion is necessary for maintaining immune homeostasis and averting pathology in the liver. Indeed, a study using HBV-carrier mouse model showed that hepatic γδ T cells play a role in promoting cell surface expression of Tim-3, Lag-3, and PD-1 expression on CD8+ T cells by mobilizing MDSC infiltration, leading to CD8+ T cell dysfunction and subsequent infection-induced liver tolerance (16).
In our study, we observed higher levels of exhaustion markers-Tim-3 and Lag-3 on peripheral γδ T cells during symptomatic CHB infection. Our preliminary results showed no changes in the expression of other exhaustion markers-CTLA-4 and PD-1 on γδ T cells from these patients (data not shown). We also found that blockade of Tim-3 and Lag-3 enhances effector function of γδ T cells. Future studies are needed to determine whether exhaustion of γδ T cells during CHB infection underlie removal of MDSC-mediated suppression of αβ T cell functions in CHB.
Another key finding in our study is the significantly higher level of CD69 expression on γδ T cells in CHB patients compared with uninfected control subjects. CD69 is widely used as an early activation marker of T cells because of its rapid induction on the surface of T cells on activation. However, CD69 has been shown to be involved in suppressing inflammatory responses by inhibiting the function of Th17 cells and augmenting suppressive potential of Treg cells (8,9,18). Importantly, an absence of CD69 expression by T cells has been reported to prevent exhaustion of the T cells and rescues antitumor effector functions of CD8+ T cells in a murine breast cancer model (20). It is, therefore, not surprising that we found only increased expression of CD69 without any significant changes in the frequency of late activation marker CD25 on γδ T cells in patients with CHB compared with HC.
In summary, our study shows that the frequencies of γδ T cell subpopulations CM (CD27+CD45RA−) and EM (CD27−CD45RA−) are decreased, whereas TEMRA (CD27−CD45RA+) γδ T cells are increased in CHB patients. This reflects the inefficiency of γδ T cells to perform effector functions during CHB infection. Our results also demonstrated higher number of γδ T cells with cell surface expression of exhaustion markers: Tim-3 and Lag-3 from CHB-infected patients. Moreover, the upregulated CD69 expression on γδ T cells during CHB infection may further contribute in downregulating γδ T cell function. Further research to understand the possible association between γδ T cells and chronic CHB in regulating immune homeostasis and averting hepatocyte pathology will provide better insights in the management and treatment of CHB infection and disease progression.
Footnotes
Acknowledgment
We thank Joy P. Gogoi for his technical assistance.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by DST INSPIRE Faculty Award, Ministry of Science & Technology, India grant DST/INSPIRE/04/2014/001692 to D.G.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.