
Abstract
Pneumonia is a pulmonary disease among children. Evodiamine, a traditional Chinese medicine, is known for anti-inflammatory effect. This study aimed to investigate the impact of evodiamine on severe pneumonia-like cells and the underlying mechanism involved. H5N1 and pneumoniae D39 was used to induce severe pneumonia-like conditions in BEAS-2B cells. The cell viability in BEAS-2B cells after treatments with 0, 20, 40, 60, 80, and 100 μM evodiamine was examined using MTT assays. The protein concentrations of inflammatory cytokines tumor necrosis factor (TNF)-α, interleukin (IL)-6 and IL-1β, and Toll-like receptors (TLRs) were measured by enzyme-linked immunosorbent assay methods and the protein and mRNA changes in C/EBPβ/CREB were measured using Real Time-quantitative polymerase chain reaction and Western blot methods. Our results revealed that Evodiamine significantly decreased TNF-α, IL-6, and IL-1β in BEAS-2B cells. Moreover, evodiamine markedly reduced TLR2,3,4 protein expression and the phosphorylated protein of C/EBPβ and CREB. Besides, evodiamine combined with clindamycin exerted more significant effects than clindamycin alone. Taken together, our results demonstrated that evodiamine enhanced the anti-inflammation effect of clindamycin in the BEAS-2B cells infected with H5N1 and pneumoniae D39 through CREB-C/EBPβ signaling pathway.
Introduction
Pneumonia is a severe lung inflammation in the children, which is caused by bacteria and viral infection and has high mortality worldwide (23). It has been previously observed that the occurrence rate of pneumonia is higher in 3- to 5-year-old children than in old people and infants (7). Previous studies showed that avian influenza, first occurring in birds or poultry, was also an epidemic in human being and caused severe pneumonia (1). To date, influenza virus H1N1, H5N1, and H7N9 could infect humans and cause influenza, further possibly induce primary viral or secondary bacterial pneumonia, resulting in respiratory distress syndrome, posing threats on lives of the infected (19,29). It is widely recognized that influenza virus triggered the inflammation cytokines, including tumor necrosis factor (TNF)-α, interleukin (IL)-6 and IL-1β, and activated the host immune activities (12,16,21,32). Clinically, neuraminidase inhibitors such as oseltamivir and zanamivir have been widely used in prevention and treatments of influenza worldwide yet the underlying side effects were worrying as well, which include nausea and vomiting. (11). Some clinical trial reported that the combined use of oseltamivir, naproxen, and clarithromycin reduced the mortality in the patients with severe influenza compared with the cases that took the medication of oseltamivir alone (13). Recently, combined use of traditional herbal medication and antiviral drugs was explored in animals, and it was found that the introduction of polygalasaponin F could not only enhance the survival rate of influenza virus-induced mice but also alleviate the side effects, including weight loss (38).
Evodiamine is an alkaloid extraction derived from fruit of Evodia rutaecarpa (Juss) Benth and was reported for its functions of anti-inflammation (24), anti-influenza A virus (4), anticardiovascular disease (8), and antiobesity (36). Evodiamine, as an inhibitor of topoisomerase I, displayed its antibacterial effect against Klebsiella pneumoniae (37). An earlier study also demonstrated the antiviral function against influenza A H1N1 virus through regulation of chemokines and cell migration of lung epithelial cells (3). However, the underlying regulatory mechanisms of evodiamine in the regulation of severe pneumonia in vitro still remains to be explored. Clindamycin is an antibiotic and has been reported to function against pneumococci and beta-hemolytic streptococci (25). Previous trials in mice with postviral bacterial pneumonia showed that the combined treatment of clindamycin and beta-lactam resulted in better outcomes than either drug alone (18). Therefore, we hypothesized that the collective use of evodiamine and clindamycin might inhibit the inflammatory cytokines overproduction more effectively than the single use of evodiamine in the severe pneumonia-like cell model. We established the cellular model by infecting the bronchial epithelial cells, BEAS-2B cells with pneumoniae D39 and H5N1 virus. Furthermore, we examined the impact of drugs on the cytokines and the potential signaling pathways involved.
Methods
Ethical statement
The cellular assays were performed in accordance with the regulations of Guangzhou Medical University.
Cell culture
BEAS-2B and MDCK cell lines were purchased from American Type Culture Collection (ATCC, VA) and cultured in an RPMI-1640 medium (Sigma Aldrich, CA) containing 10% fetal bovine serum, 100 U/mL penicillin, and 100 U/mL streptomycin. BEAS-2B cells are human bronchial epithelial cells. MDCK is short for Madine Darby canine kidney cells, which was used for adaptive incubation for H5N1 virus. The cells were incubated in a humidified incubator with 5% CO2 at 37°C. The medium was replaced every 2–3 days.
Bacteria and virus preparation and coinfection
The serotype 2 Streptococcus pneumoniae D39 was used in this study (ATCC, CA) and petri plates were used to culture the bacteria adding trypticase soy agar II (modified; Becton-Dickinson) and 5% defibrinated sheep blood (TSAII-BA), incubating 5% CO2 at 37°C, according to the previous report (20). The avian influenza virus Jilin/9/2004(H5N1) was bought from Chinese Center for Disease Control and Prevention (Beijing, China). The virus propagation was placed in the cavities of 10-day-old chicken eggs and the virus adapted to grow in MDCK cells.
Cells were incubated with D39 first as following the method previously reported (9). Cells were then washed with phosphate-buffered saline (PBS) for three times followed by the incubation using RPMI-1640 medium (1 mL) containing H5N1 with 5% CO2 at 37°C. The suspension was removed after 3 h of incubation and cells were washed with PBS followed by the re-incubation with evodiamine, clindamycin, or both for 24 h.
Drug treatment
Evodiamine was purchased from Macklin (Shanghai, China) and was placed in 100% dimethyl sulfoxide. Clindamycin was bought from Beyotime. Different concentrations of evodiamine (0, 20, 40, 60, 80, and 100 μM) was added to treat the cells for 24 h in an incubator with 5% CO2 at 37°C. BEAS-2B cells coinfected with D39 and H5N1 were seeded on 96-well plates (5 × 104 cell/well) and the RPMI-1640 medium was used to culture the cells. Evodiamine (40 μM), 4 μM clindamycin, or both was added to treat the infected cells for 24 h in an incubator with 5% CO2 at 37°C. Cells were further collected for the functional assays.
MTT assay
Ten microliters of MTT solution (Sigma Aldrich, MO) was added to each well and the cells were incubated for 4 h at 37°C. The supernatant was removed and dimethyl sulfoxide (100 μL; Sigma Aldrich) was added to dissolve formazan followed by the incubation. The absorbance was measured by the laboratory microplate reader at a wavelength of 490 nm.
Real Time-quantitative polymerase chain reaction
Trizol reagent was used to extract total RNA from BEAS-2B cells according to the instructions given by manufacturer (Beyotime, Shanghai, China). PrimeScript RT Master Mix kit was used for the reverse transcription of RNA into cDNA (Takara, Beijing, China). SYBR Premix Ex Tap II was used to perform Real Time-quantitative polymerase chain reaction (RT-qPCR) on cDNA samples (Takara). ABI PRSIM 7500 Real-time PCR system (Applied Biosystem, Shanghai, China) was used to analyze the PCR data and a 2−ΔΔCt method was used to determine the relative expressions. The primers used are given in Table 1.
Primers Used
Enzyme-linked immunosorbent assay method
The cell supernatants from all groups were collected for enzyme-linked immunosorbent assay (ELISA) experiments and the specific ELISA kits for human Toll-like receptor (TLR)2,3,4, TNF-α, IL-6, and IL-1β (Abcam, Shanghai, China) were used to measure the concentrations of the cytokines by strictly following the protocols. The OD values were read at the wavelength of 450 nm.
Western blot analysis
The total proteins were extracted from cells in each group, respectively, using RIPA buffer (Solarbio, Beijing, China). Then the proteins were separated on sodium dodecyl sulfate-polyacrylamide gel electrophoresis and further transferred onto polyvinylidene difluoride (PVDF) membranes. Five percent skim milk and Tris Buffered Saline Tween (TBST) were added, and the membranes were incubated for 2 h at room temperature. Later, the primary antibodies were added, including anti-C/EBPβ, anti-CREB, antiphospho-CREB and anti-β-actin from Cell Signaling (Shanghai, China). The membranes were incubated for a night at 4°C. On the second day, the membranes were incubated further with the secondary antibody antirabbit immunoglobulin G, horseradish peroxidase (1:1,000) for 2 h at room temperature. TBST was used to wash the PVDF membranes for five times and SignalFire™ Plus ECL kit was used to measure the protein expression, respectively. ImageJ (
Statistical analysis
Each experiment was performed at least three times. All the data were analyzed by using SPSS (IBM, IL) and expressed as the mean ± standard deviation. GraphPad Prism 7.0 software (GraphPad Software, CA) was used for figure generation based on the results. Student's t-test was used for the comparison between two groups, whereas one-way analysis of variance was used for the comparison among multiple groups followed by Tucky corrections. The general alpha is 0.05.
Results
Evodiamine inhibited inflammatory cytokines in BEAS-2B cells coinfected with D39 strain and H5N1 virus
The cell viability in BEAS-2B cells after evodiamine treatment (0, 20,40, 60, 80, and 100 μM) were measured by using MTT assays and results showed that no significant changes were noted in the groups of 0–40 μM treatment of evodiamine (Fig. 1A). Therefore, we selected this range of concentration in the following experiments. Also, protein concentrations of proinflammatory cytokines, TNF-α, IL-6, and IL-1β were measured using ELISA methods. It was found that TNF-α, IL-6, and IL-1β were overproduced after cells were infected with the D39 strain and H5N1 virus while evodiamine inhibited the inflammatory cytokines and the 40 μM was more significant in inhibitory effect on TNF-α, IL-6 and IL-1β cytokines (Fig. 1B).
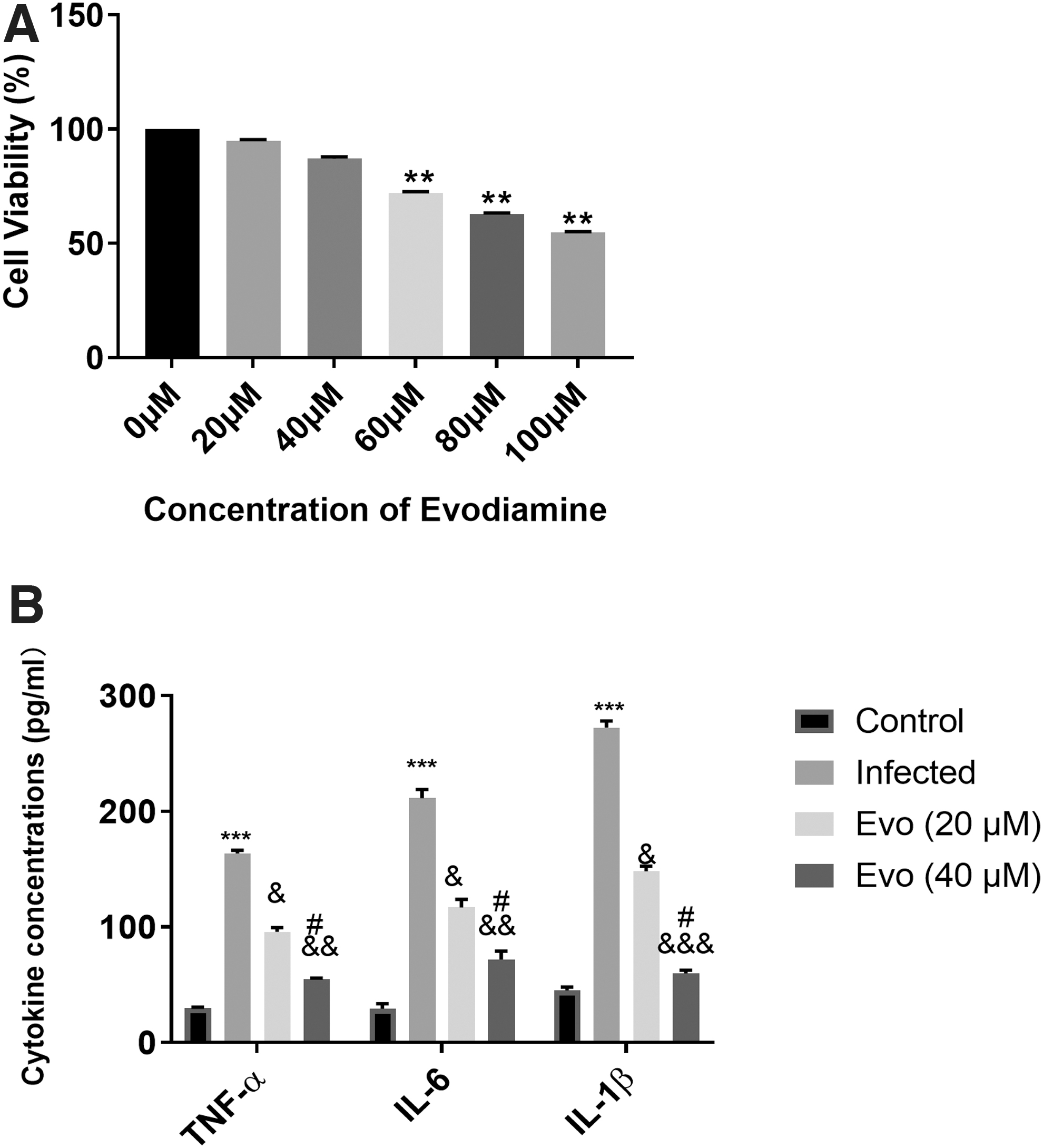
Evodiamine inhibited inflammatory cytokines in BEAS-2B cells coinfected with D39 strain and H5N1 virus.
Effect of evodiamine on TLRs and CREB-C/EBPβ signaling pathway in BEAS-2B cells coinfected with D39 strain and H5N1 virus
ELISA method also examined the TLR2,3,4 protein concentrations in cells after coinfection, and additional evodiamine treatment (20, 40 μM) and it found that the TLR2,3,4 were provoked by the coinfection and inhibited by the introduction of evodiamine (Fig. 2A). Forty micromolar evodiamine inhibited the TLR2,3,4 more significantly than the 20 μM evodiamine (Fig. 2A). RT-PCR tests showed that the mRNA expression levels of CREB and C/EBPβ were also enhanced in the coinfected cells and the addition of evodiamine decreased the mRNA expression and 40 μM group displayed the lower expression than the 20 μM group (Fig. 2B). Western blot results revealed that the phosphorylation rate of CREB and the C/EBPβ protein expression was promoted by the coinfection and inhibited by the treatment of evodiamine (Fig. 2C, D).

Effect of evodiamine on TLRs and CREB-C/EBPβ signaling pathway in BEAS-2B cells coinfected with D39 strain and H5N1 virus.
Evodiamine promoted the effect of clindamycin in inflammatory cytokines inhibition in coinfected BEAS-2B cells.
ELISA methods assessed the changes in cytokine concentrations of TNF-α, IL-6, and IL-1β and pointed out that the evodiamine consolidated the function of clindamycin in inhibiting TNF-α, IL-6 (Fig. 3).
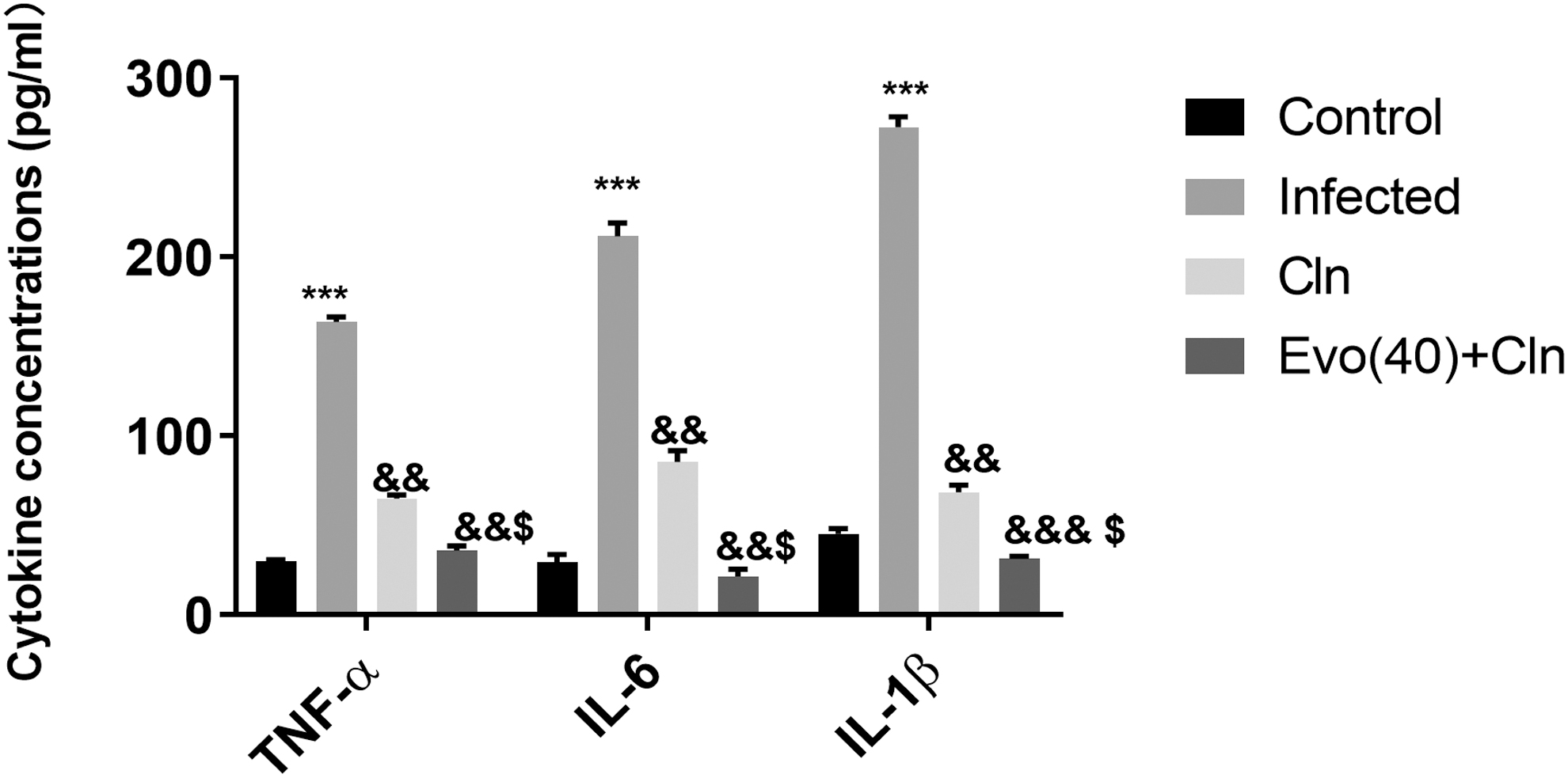
Evodiamine promoted the effect of clindamycin in inflammatory cytokines inhibition in coinfected BEAS-2B cells. The cytokine concentration of TNF-α, IL-6, and IL-1β were measured using ELISA method in infected BEAS-2B cells after treatment with clindamycin alone, or together with evodiamine (40 μM). ***p < 0.002, versus control. &&& p < 0.002, && p < 0.005, & p < 0.01, versus infected. $ P < 0.01 versus Cln. Cln: clindamycin; Evo(40): evodiamine 40 μM.
Evodiamine enhanced the clindamycin effect on inactivation of TLR/CREB-C/EBPβ signaling pathway in BEAS-2B cells
To further determine the anti-inflammatory effect of evodiamine in combination with clindamycin, the protein concentrations of TLR2,3,4 were examined using ELISA assays and results showed that Evodiamine further increased the inhibitory function of clindamycin in TLR2,3,4 (Fig. 4A). Moreover, C/EBPβ and CREB mRNA expression was also downregulated by clindamycin, and evodiamine involvement inhibited the expression further compared with clindamycin alone (Fig. 4B). Western blot analysis showed that CREB phosphorylation and C/EBPβ protein levels were reduced by clindamycin and the combined use of evodiamine further inactivated the CREB-C/EBPβ pathway (Fig. 4C, D).
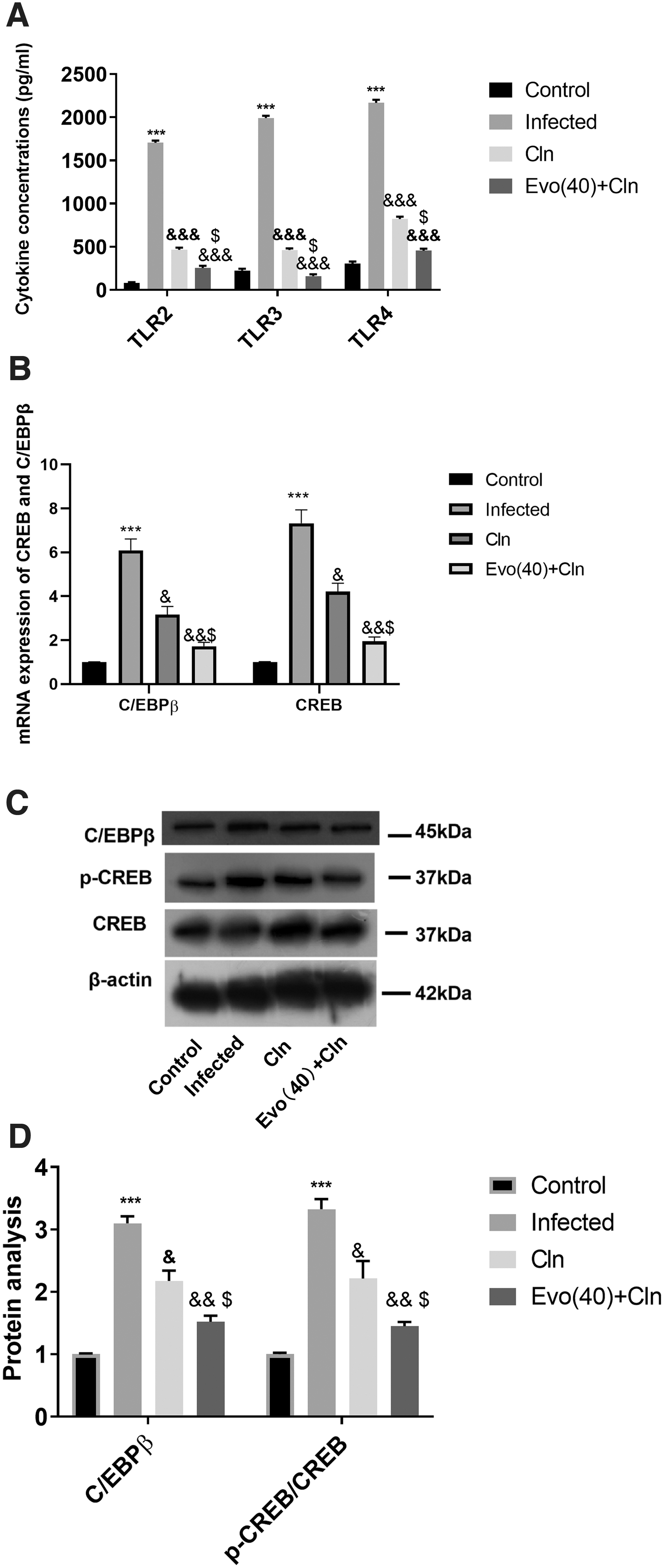
Evodiamine enhanced the clindamycin effect on inactivation of TLR/CREB-C/EBPβ signaling pathway in BEAS-2B cells. Coinfected BEAS-2B cells were further treated with clindamycin alone or with evodiamine.
Discussion
In this study, we established a cellular model of severe pneumonia by infecting the BEAS-2B cells with pneumoniae D39 strain and the influenza virus H5N1. A previous study introduced the same cellular model with ours, based on which, it was disclosed that Menispermum dauricum extraction, combined with clindamycin, exerted its antiviral and antibacterial function in vitro and also animal models through regulating a canonical inflammation-related pathway, nuclear factor-κB (NF-κB) (22). Clindamycin was used with primaquine for the treatment of acute pneumonia previously (39). In this research, we investigated a more popular herbal extraction, evodiamine in the severe pneumonia-like cell model. We discovered that evodiamine lowered the proinflammatory cytokines, IL-6, TNF-α, and IL-1β, which was triggered by the coinfection, which was similar to the previous finding in the dauricine study (22). Furthermore, we also found that the evodiamine also inhibited the TLR2,3,4 protein expression and inactivated the CREB-C/EBPβ pathway in the cells. Also, the clindamycin presented the similar role to evodiamine in this study and the combined use showed that evodiamine promoted the function of clindamycin.
The TLRs family are recognized as pattern recognition receptors and closely related to inflammation and innate immunity (2). TLR2,3,4 interact with bacteria and virus by detecting the lipoproteins, lipopolysaccharide (LPS)/MD2 and dsRNA, respectively, in cells (6,35). Previous study reported that TLR3/4 knockout might help to exempt mice from infection with H5N1 (33). And TLR3 was activated by influenza A virus, triggering serious inflammatory cytokines (14).TLR2/4 were involved in the responses S. pneumoniae (30). It was also reported that through TLR2 activation, the proinflammatory cytokines were provoked in the context of H5N1 infection (15,34). Therefore, we examined the TLR2,3,4 in our study and found that the changes in TLR2,3,4 were in accordance with those in the proinflammatory cytokines, IL-6, TNF-α, and IL-1β, suggesting that the antiviral and antibacterial function of evodiamine might exert through the TLR2,3,4 inhibition in this cell model.
CREB was involved in inflammation regulation (27) and triggering the signaling pathway could contribute to the encoding of inflammatory cytokines such as IL-10, IL-1β, and chemokines such as CXCL1 (17). CREB, cAMP responsive element binding protein, was revealed to mediate TLR4 in inflammation (5). TLR2/4 was reported to induce CREB in macrophages (26). Moreover, the DNA-binding activity of CREB was upregulated by the inflammation induction by LPS in rat lungs (28). C/EBPβ, CCAAT-Enhancer-binding Protein β, has shown its modulational role in adipocyte differentiation (10) and inactivation of reduced the inflammation and endoplasmic reticulum stress (40). C/EBPβ is a downstream target of CREB, and the inhibition of CREB-C/EBPβ signaling pathway was discovered to reduce the bacteria survival in infected cells (31). However, the performance of CREB-C/EBPβ pathway in the cells coinfected with pneumoniae D39 strain and the influenza virus H5N1 still remained unknown. We unraveled that the CREB-C/EBPβ pathway was provoked by the coinfection and was inactivated by evodiamine or clindamycin and further inhibited by the combined use of both drugs.
Taken together, our findings concluded that evodiamine further enhanced the anti-inflammation effect of clindamycin in the BEAS-2B cells infected with H5N1 and pneumoniae D39 strain through CREB-C/EBPβ signaling pathway.
Availability of Data and Materials
All data used in this study are available from the corresponding author on reasonable request.
Footnotes
Authors' Contributions
X.L. and X.P. conceptualized, planned, and carried out the experiments. X.L. contributed in the experiments and carried out the data analysis. X.L. carried out the data curation and wrote the draft of the article. X.P. was involved in data visualization and contributed in conducting the experiments. X.L. handled the software analysis and validation work. X.P. and Z.L. contributed to the experiments. All authors read the article, provided critical feedback, and approved the study for publication.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
There is no funding involved.