
Abstract
Porcine circovirus type 2 (PCV2) is the causative agent of PCV-associated disease, which harms the swine industry worldwide. Open reading frame 2 of PCV2 encodes the principal immunogenic capsid (Cap) protein, which induces neutralizing antibodies and protective immunity. Cap has been developed as a subunit vaccine against PCV2 infection, although its use is hindered by low immunogenicity. Here, we hypothesized that the truncated diphtheria toxin DT390 might enhance the immunogenicity of Cap. To verify this hypothesis, we fused Cap with DT390, which was expressed using the unique diphtheria toxin-resistant Pichia pastoris expression system. We assessed the immunogenicity of DT390-Cap using BALB/c mice. DT390-Cap induced significantly higher Cap-specific and neutralizing antibodies than Cap alone with or without the ISA201 adjuvant. DT390-Cap with ISA201 adjuvant induced production of more Cap-specific antibodies and neutralizing antibodies than Ingelvac CircoFLEX (positive control). DT390-Cap induced slightly higher Th2-associated interleukin-4 production than Cap alone but did not affect Th1-associated interferon-γ production. The protection study demonstrated that DT390-Cap induced more effective protective immunity than Cap alone, when challenged with PCV2. The viral loads in the lungs, liver, and thymus in mice immunized using DT390-Cap were significantly lower than in those immunized with the corresponding Cap with or without the ISA201 adjuvant. Taken together, the engineered DT390 effectively enhanced the immunogenicity and protective immunity of Cap in mice. Thus, DT390-Cap is a promising novel vaccine candidate against PCV2 infection.
Introduction
Porcine circovirus (PCV) belongs to the family of Circoviridae and contains a single-stranded circular DNA genome (7,24). There are four recognized types of PCVs: porcine circovirus 1 (PCV1), porcine circovirus 2 (PCV2), porcine circovirus 3 (PCV3), and porcine circovirus 4 (PCV4) (16). PCV2 has been extensively studied in the past few decades and is considered to be the main causative agent for PCV-associated disease, which is now recognized as a serious threat to the swine industry worldwide (17,31). PCV2 infection is widespread in almost all pig-producing countries (1,8).
The PCV2 genome contains two major open reading frames (ORFs), namely, ORF1 and ORF2. ORF1 encodes two replication-related proteins (Rep and Rep′) (11). ORF2 encodes the main immunogenic capsid (Cap) protein, which is responsible for producing the PCV2-specific neutralizing antibodies (13,15). PCV2 Cap subunit vaccines are one of the most successful tools for protecting pigs against PCV2 infection (1,3). However, low immunogenicity is still the main limitation of the Cap subunit vaccine (18). Previously, our laboratory has expressed Cap using the yeast Pichia pastoris and demonstrated limited immunogenicity and protective immunity in mice (Tao et al., unpublished data). The expressed Cap protein was used as a control in this study.
CRM197 is a well-characterized diphtheria toxin mutant with a good safety profile as a carrier protein in vaccines (12,19,21). Truncated diphtheria toxin DT390 contains the catalytic domain and transmembrane domain of diphtheria toxin (29). Based on our long-term working experience with DT390 in immunotoxin development, we hypothesized that DT390 might enhance the immunogenicity of the subunit vaccines. In this study, we genetically engineered Cap to truncated diphtheria toxin DT390 and expressed the DT390-Cap fusion protein using the diphtheria toxin-resistant P. pastoris expression system. We assessed the immunogenicity and ability to provide protective immunity of DT390-Cap against PCV2 using BALB/c mice.
Materials and Methods
Virus, cell lines, and antibodies
The strain of PCV2 SH (GenBank: AY686763.1) was provided by Prof. Jiang Ping of the Nanjing Agricultural University. The original virus titer was 106.4 TCID50/mL (30). PCV2-free PK-15 cells were cultured for more than three passages in Dulbecco's modified Eagle's medium (DMEM; Gibco, Beijing, China) supplemented with 10% fetal bovine serum (FBS; Biological Industries, Beit Haemek, Israel) at 37°C in a 5% CO2 incubator. PCV2 was inoculated in a 50 − 60% confluent monolayer of PK-15 cells. After incubation at 37°C for 2 h, the culture medium was discarded and fresh DMEM containing 2% FBS was added, following which the cells were cultured at 37°C continuously for ∼24–48 h. After reaching confluence of >90%, the cells were passaged using 0.25% trypsin. Then, the cells were cultured until passage 20. The cells and medium were collected and freeze-thawed thrice, followed by centrifugation at 3,000 rpm and 4°C for 30 min; the virus-containing supernatant was collected and stored at −80°C until subsequent usage. PCV2 virus of passage number 20 was used for the challenge study. The concentration of the harvested PCV2 virus solution was 107.5 TCID50/mL, measured by indirect immunofluorescence. The directly harvested PCV2 virus solution was diluted 10 times as working PCV2 virus solution for the infection/challenge. The concentration of the working PCV2 virus solution was 106.5 TCID50/mL. The infection/challenge dose was 0.85 mL of 106.5 TCID50/mL per mouse.
Horse radish peroxidase (HRP)-conjugated anti-His tag mouse monoclonal antibodies (mAbs) (Cat No. CW0285), goat anti-mouse IgG-HRP (Cat No. CW0102) antibodies, and goat anti-rabbit IgG-HRP (Cat No. CW0103) antibodies were purchased from CWBIO (Beijing, China). Mouse mAbs [8A4] against diphtheria toxin A subunit (Cat No. ab8308) were obtained from Abcam (Cambridge, United Kingdom). PCV2 Cap antibodies (Cat No. orb312690) were obtained from Biorbyt (Britain, United Kingdom). PCV2 indirect immunofluorescence detection kit was purchased from the China Institute of Veterinary Drug Control Institute (containing specific mAbs for PCV2, FITC-labeled IgG antibody).
Protein expression and purification
To construct DT390-Cap, PCV2 Cap DNA (PCV2 Cap) was amplified (using PCV2 Cap in pwPICZalpha as template) using polymerase chain reaction (PCR) primers, pCap-Nco harboring XhoI and NcoI sites (5′CCG CTC GAG CCA TGG GGT GGT GGT GGT TCT ACT TAC CCA AGA AGA AGA TAC AGA3′) and pCap-Eco harboring the EcoRI site (5′CCG GAA TTC TTA GTG GTG GTG GTG GTG GTG TGG GTT CAA AGG TGG GTC CTT3′); the PCR product was then cloned in the pwPICZalpha vector between XhoI and EcoRI sites for sequencing. The pwPICZalpha vector was derived from Invitrogen's yeast P. pastoris expression vector pPICZalpha with minor modification (29). The insert was subsequently excised using NcoI and EcoRI and cloned in DT390-containing P. pastoris expression vector pwPICZalpha-DT390 between NcoI and EcoRI sites, yielding the final construct DT390-Cap in pwPICZalpha. The linearized DT390-Cap DNA construct was transformed in diphtheria toxin-resistant P. pastoris strain (10), and the transformants were selected on YPD agar plates (1% yeast extract, 2% peptone, 1.5% agar, and 2% dextrose) containing 100 μg/mL zeocin. Six colonies were randomly selected and cultivated to express protein on a small scale. The culture supernatants were analyzed using sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and western blotting using anti-His tag mAbs under nonreducing conditions [Invitrogen™ NuPAGE™ LDS sample buffer (4 × ), Cat No. NP0007; Thermo Fisher Scientific, Waltham, MA]. After analysis, one clone was selected and cultured to express DT390-Cap on a large scale, as described previously (6). The secreted DT390-Cap in the supernatant was captured using two-step purification. A Ni-Sepharose fast flow resin was used for the first step of purification. The bound proteins were eluted with elution buffer containing 500 mM imidazole. A strong anionic exchange resin, Poros 50HQ, was used for the second step of the purification. The bound protein was eluted with elution buffer containing 200 mM sodium borate and 100 mM NaCl. The details of the purification procedures have been described previously (6). Cap alone was also expressed and purified using the P. pastoris expression system.
Western blotting
Protein samples were mixed with nonreducing loading buffer or treated with PNGase F reagent (New England Biolabs, Beijing, China), separated using SDS-PAGE, stained with Coomassie Blue Fast, and electrotransferred onto a polyvinylidene fluoride membrane (Millipore, MA) at 200 mA for 1 h. The membrane was blocked using 5% nonfat dry milk in 1 × TBS-0.02% Tween 20 for 2 h with shaking and then incubated with HRP-conjugated anti-His mouse mAbs (1:10,000), anti-PCV2 Cap antibodies (1:1,000), and mouse anti-diphtheria toxin A subunit mAbs [8A4] (1:5,000) at room temperature for 2 h with shaking. After three washes with Tris-buffered saline–Tween 20, the membrane was incubated with goat anti-mouse IgG-HRP (1:20,000) and goat anti-rabbit IgG-HRP (1:20,000) at 37°C for 1 h with shaking. 3,3′,5,5′-tetramethylbenzidine (TMB) one-component membrane peroxidase substrate (Seracare, MA) was used for the detection of proteins per the manufacturer's instructions.
Assessment of immunogenicity in mice
The detailed experimental design is given in Figure 1 and Table 1. Thirty of 6-week-old specific pathogen-free grade female BALB/c mice were purchased from Charles River (Beijing, China). The mice were randomly assigned to six groups of five animals per group and were humanely managed according to the established guidelines of the animal ethical committees of Shanxi Agriculture University. The groups were as follows: (i) phosphate-buffered saline (PBS)+ISA201 as the negative control group (n = 5), in which the animals were intramuscularly immunized with PBS mixed with Montanide ISA201 (Seppic, Shanghai, China) in 50 μL in a ratio of 25:25 (v/v) according to the manufacturer's instruction; (ii) Ingelvac CircoFLEX® vaccine as the positive control group (n = 5), in which the animals were intramuscularly immunized with 1.8 μL/g of Ingelvac CircoFLEX (commercial PCV2 subunit vaccine, Lot No. 309-570; Boehringer Ingelheim Vetmedica, China) in 50 μL. The 1.8 μL/g murine dose was converted from porcine dose based on the following formula: murine dose = the porcine dose × R (where R = 9). The dose is 1 mL for a 5 kg pig. Murine dose = 0.2 mL/kg × 9 = 1.8 mL/kg (1.8 μL/g) (5); (iii) Cap alone group (n = 5), in which the animals were intramuscularly vaccinated with Cap protein (165 μg/kg in 50 μL); (iv) DT390-Cap group (n = 5), in which the animals were intramuscularly vaccinated with DT390-Cap protein (400 μg/kg in 50 μL); (v) Cap+ISA201 group (n = 5), in which the animals were intramuscularly vaccinated with Cap (165 μg/kg in 25 μL) mixed with Montanide ISA201 (v/v, 25:25); (vi) DT390-cap+ISA201 group (n = 5), in which the animals were intramuscularly vaccinated with DT390-Cap (400 μg/kg in 25 μL) mixed with Montanide ISA201 (v/v, 25:25). One hundred sixty-five micrograms cap protein and 400 μg DT390-Cap protein both contain equal molecular mass of 5.79 × 10−3 μmol. The serum samples were collected from the orbital vein for measurement of antibodies against Cap and PCV2-neutralizing antibodies −3, 14, 21, 28, and 42 days after the primary immunization (dpi). At 28 dpi, all groups were challenged with 0.85 mL of 106.5 TCID50/mL PCV2 per mouse through three administration routes, including intraperitoneal injection (0.5 mL), subcutaneous injection (0.3 mL), and nasal drops (0.05 mL). All mice were killed at 42 dpi and samples from the lungs, liver, and thymus were collected for detection of PCV2 viral loads.
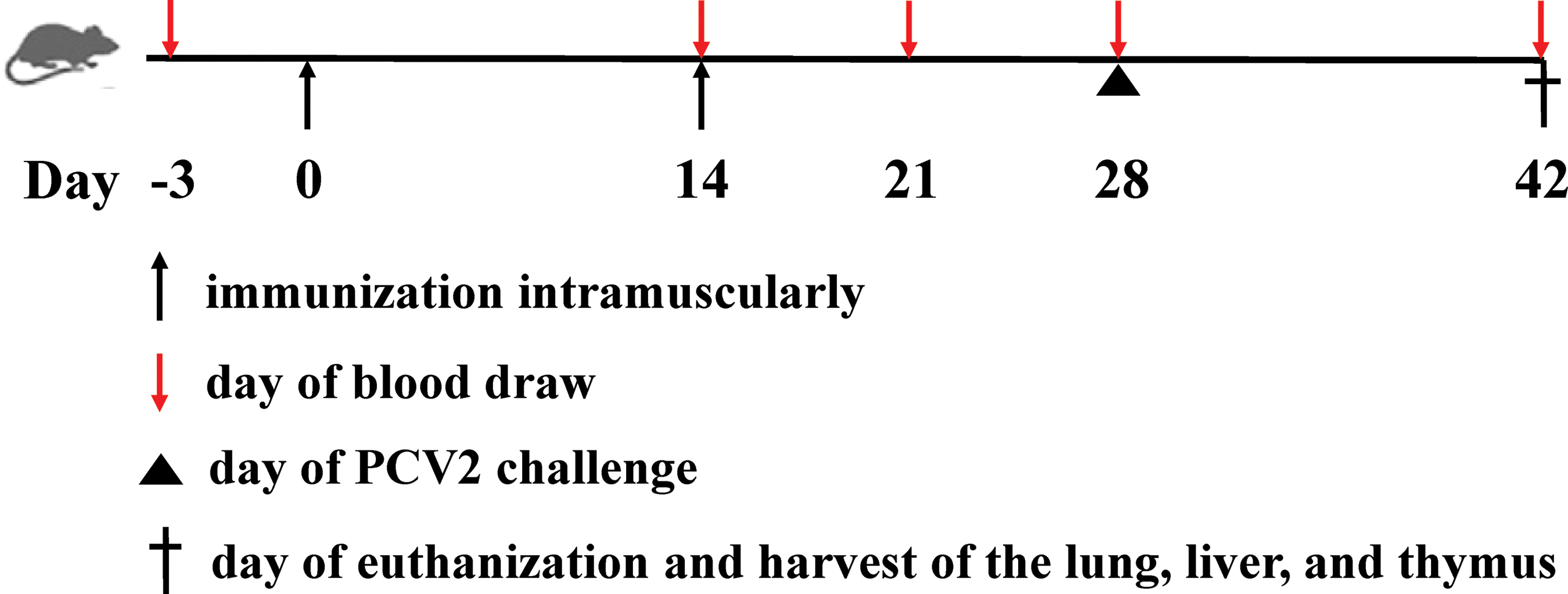
Schematic showing the in vivo immunogenicity and protection study of DT390-Cap in BALB/c mice. Cap, capsid. Color images are available online.
Summary of the In Vivo Immunogenicity Study of DT390-Cap
Two immunizations: day 0 as primary immunization and day 14 as booster immunization.
Cap, capsid; IM, intramuscular; PBS, phosphate-buffered saline; W/O/W, water-in-oil-in-water.
Emulsion of the antigen protein with Montanide ISA201 was prepared as follows. Twenty-five microliters each of the antigen protein and Montanide ISA201 were prewarmed separately in a water bath (31°C) for 5 min, following which the prewarmed antigen protein was dropped into the prewarmed ISA201 slowly to a final ratio of 1:1 (v/v), with gentle shaking on the vortex shaker for 5 min. The emulsion was cooled in a cold bath at 20°C for 1 h and then injected into mice.
Indirect enzyme-linked immunosorbent assay
The level of IgG specific for Cap in the immunized murine serum was measured using enzyme-linked immunosorbent assay (ELISA). In brief, ELISA plates (Costar) were coated with 100 μL of purified recombinant Cap (expressed and purified in our laboratory using the P. pastoris expression system), which was diluted to a final concentration of 2 μg/mL in coating buffer (pH = 9.6), incubated overnight at 4°C, and then washed thrice with PBS containing 0.05% Tween 20 (PBST; pH = 7.4). Nonspecific binding of the antibodies was blocked with 100 μL of PBST containing 1% bovine serum albumin (BSA) for 2 h at 37°C. The plate was washed thrice with PBST. The serum sample was diluted twofold in 1% BSA-PBST from 1:25 to 1:800 dilutions. Fifty microliters of 1:800 diluted serum sample was added to each well and incubated for 1 h at 37°C. After washing thrice with PBST, the bound antibodies were incubated with 100 μL goat anti-mouse IgG (1:5,000) diluted in 1% BSA-PBST. After incubation for 30 min at 37°C and three PBST washes, 100 μL TMB (Solarbio, Beijing, China) was added to each well and the mixture was incubated for 15 min at 37°C. The reaction was stopped by adding 50 μL of 1 N sulfuric acid to the mixture, and the optical density at 450 nm was measured (Berthold microplate reader; Berthold Technologies, Bad Wildbad, Germany).
Detection of neutralizing antibody
PCV2-neutralizing activity was measured using the endpoint dilution reduction assay. The pooled serum samples (20 μL per mouse) from each group were heat inactivated at 56°C for 30 min. One hundred microliters of twofold serial dilutions of serum were incubated with 50 TCID50 virus in 100 μL for 2 h at 37°C. The serum-virus mixtures were inoculated in 70 − 80% confluent PK-15 cells cultured in 96-well plates and incubated at 37°C for 2 h. The inoculum was removed, and the cell cultures were washed thrice with DMEM. The maintenance medium was then added for incubation. After 72 h, the culture plate was fixed with 80% cold acetone solution at 4°C for 30 min and blocked with 3% BSA in PBS at 37°C for 1 h. The cells were incubated with specific mAbs for PCV2, followed by FITC-labeled IgG antibody as the secondary antibody. The cells were observed using a fluorescence microscope (Olympus, Tokyo, Japan). The neutralization antibody titer was determined as the reciprocal of the highest serum dilution that was able to reduce ≥80% fluorescent focus in PK15 cells as described previously (9).
Cytokine measurement
The serum samples were collected from the experimental mice 21, 28, and 42 dpi. The serum interferon (IFN)-γ and interleukin (IL)-4 levels were measured using a mouse cytokine ELISA kit (BlueGene Biotech, Shanghai, China) according to the manufacturer's instructions.
Measurement of PCV2 viral load
PCV2 DNA from the lungs, liver, and thymus of all experimental groups at 14 days postchallenge (dpc) was quantified using real-time fluorescent PCR. Gene copy number, representing viral load, was calculated according to the standard curve by plotting cycle threshold values against the copy number of different dilutions of a standard recombinant plasmid vector containing PCV2 Cap gene fragment (23).
Statistical analysis
Statistical analysis was performed using the GraphPad Prism 7.04 software (San Diego, CA). All data were expressed as the mean ± standard error of mean. Differences among multiple groups were assessed using one-way analysis of variance (ANOVA), followed by Tukey's multiple comparison test. Significant statistical differences were expressed as follows: *p < 0.05, **p < 0.01, ***p < 0.001, and ****p < 0.0001. p < 0.05 was considered the threshold for statistical significance.
Results
Expression and purification of DT390-Cap
As given in Figure 2A, DT390-Cap was composed of two domains: truncated diphtheria toxin DT390 and Cap. Codon-optimized PCV2 Cap DNA was cloned in the DT390-containing P. pastoris expression vector pwPICZalpha-DT390 (27). To facilitate purification, we added a six histidine (6 × His) tag to the C-terminus of the construct. DT390-Cap was expressed in diphtheria toxin-resistant P. pastoris using shake flasks. The secreted DT390-Cap protein in the supernatant was directly captured using a Ni-Sepharose fast flow resin. A strong anionic exchange resin, Poros 50HQ, was used for the second step of purification. SDS-PAGE and western blot analysis confirmed the expression and purification of the DT390-Cap fusion protein (Figs. 2B and 3A–C). Anti-His mAb, anti-PCV2 Cap polyclonal antibody, and mouse anti-diphtheria toxin A subunit mAb were used for the western blot analysis. The final purification yield was ∼3 mg/L of the original harvested supernatant.
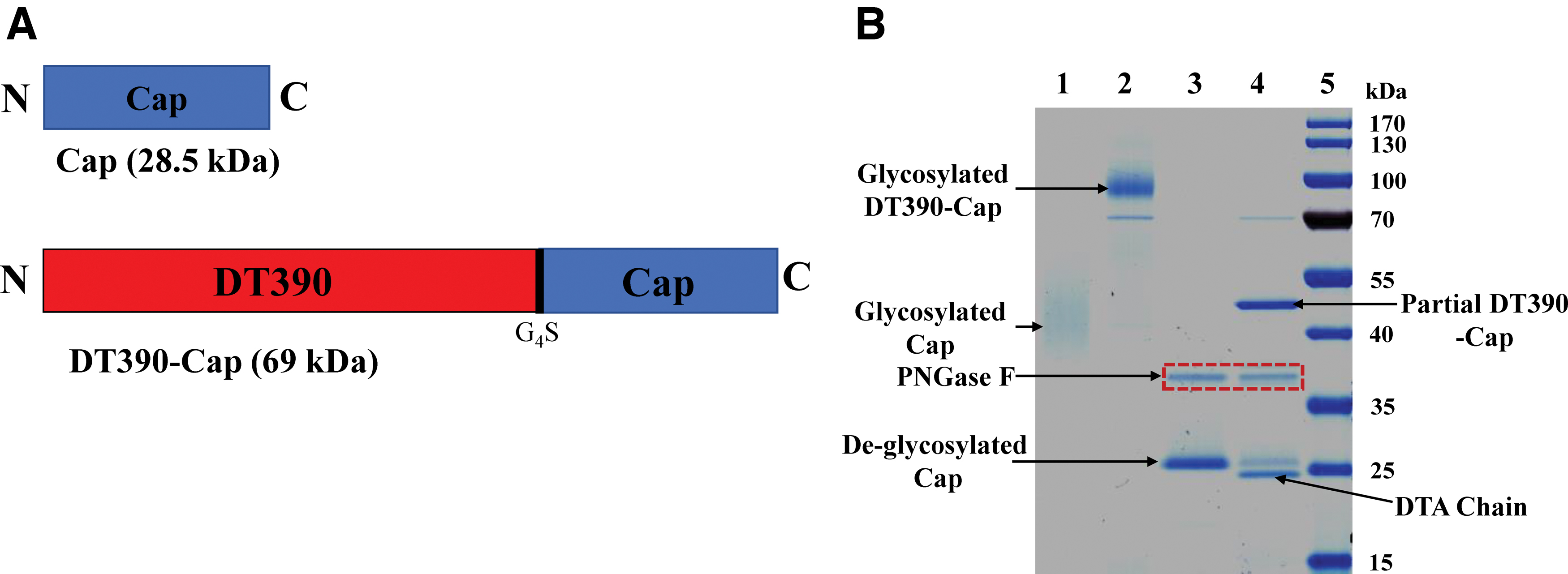
Schematic diagrams and SDS-PAGE analysis of DT390-Cap.
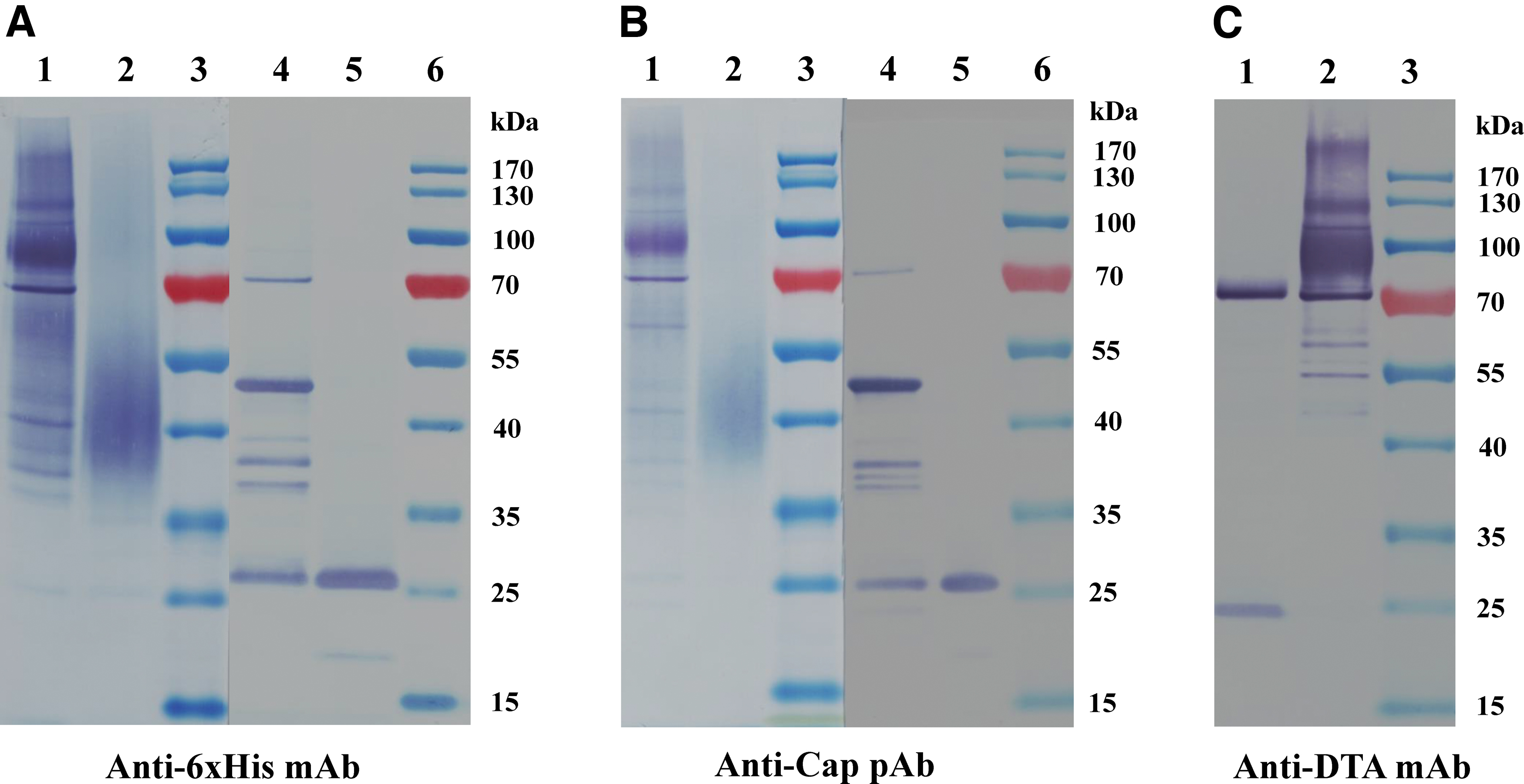
Western blot analysis of DT390-Cap.
DT390-Cap elicited higher humoral immune response
BALB/C mice were used to assess the in vivo immunogenicity of DT390-Cap. Considering the potential nonspecific toxicity of the truncated diphtheria toxin DT390, we first determined the maximum tolerated safe dose of DT390-Cap in BALB/C mice to be 400 μg/kg (data not shown). The following six groups were included in this study: (i) PBS+ISA201, as the negative control group; (ii) Ingelvac CircoFLEX vaccine, as the positive control group; (iii) Cap alone group; (iv) DT390-Cap group; (v) Cap+ISA201 group; (vi) DT390-cap+ISA201 group. Intramuscular immunization at the back of the thigh was performed on days 0 and 14 (Fig. 1 and Table 1). Levels of Cap-specific antibodies were monitored using indirect ELISA. Recombinant Cap was used to coat the ELISA plate. All immunization groups were negative at −3 and 14 dpi. At 21 dpi, Cap-specific IgG was detected in some immunization groups, including the Ingelvac CircoFLEX positive control group, DT390-cap group, Cap+ISA201 group, and DT390-Cap+ISA201 group, whereas negative results were obtained at this time point with the negative control and Cap alone group. Cap-specific IgG levels of DT390-Cap+ISA201 were remarkably higher than those of the positive control Ingelvac CircoFLEX, DT390-Cap, and Cap+ISA201 groups. At 28 and 42 dpi, the Cap alone group was still negative for Cap-specific IgG. The OD450 value of DT390-Cap+ISA201 group was ∼2- to 3-fold higher than that of the Cap+ISA201 group (Fig. 4).

Cap-specific antibody response detected using indirect ELISA. Collected serum samples were analyzed for the presence of IgG against Cap using indirect ELISA. The purified recombinant Cap protein was used to coat the ELISA plate. Optical densities were read at 450 nm. Data are shown as mean ± SEM. Statistical significance of the differences between each of the groups was analyzed using one-way ANOVA. Statistical significance has been indicated as follows: *p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001. ANOVA, analysis of variance; ELISA, enzyme-linked immunosorbent assay; SEM, standard error of mean. Color images are available online.
Neutralizing antibodies against PCV2 play an important role in protecting pigs from PCV2-associated diseases. Therefore, the titers of neutralizing antibodies (serum dilution) were assessed in immunized mice using this vaccine candidate (22). As given in Figure 5, at 21 dpi, low neutralizing antibody titers were detected in the DT390-Cap+ISA201 group (1:3) and Ingelvac CircoFLEX positive control group (1:1). At 28 dpi, the neutralizing antibody titers were detected in all immunization groups (Ingelvac CircoFLEX group, 1:6; DT390-Cap group, 1:3; Cap+ISA201 group, 1:3; DT390-Cap+ISA201 group, 1:6) except for those in the PBS+ISA201 and Cap alone groups, which were consistent with the indirect ELISA titers. At 42 dpi, neutralizing antibody titers were detected in all immunization groups (PBS+ISA201 group, 1:3; Ingelvac CircoFLEX group, 1:12; Cap group, 1:3; DT390-Cap group, 1:6; Cap+ISA201 group, 1:12; DT390-Cap+ISA201 group, 1:24). The neutralization antibody titer of the DT390-Cap+ISA201 group was the highest among all groups at 42 dpi and was significantly higher (twofold) than that of the positive control Ingelvac CircoFLEX vaccine group. Therefore, our results demonstrated that the DT390-cap with or without ISA201 adjuvant effectively induced specific and neutralizing antibodies in mice.

Measurement of PCV2-neutralizing antibodies in serum of immunized mice. Serum samples collected were examined for the PCV2-neutralization antibody titers using an endpoint dilution reduction assay. The neutralizing antibody titer was determined as the reciprocal of the highest serum dilution that was able to reduce ≥80% fluorescent focus in PK15 cells. PCV2, porcine circovirus type 2. Color images are available online.
IFN-γ and IL-4 levels after the DT390-Cap immunization
Although the Cap subunit vaccine mainly elicited humoral response (32), we monitored the levels of the Th1-associated representative cytokine IFN-γ and Th2-associated representative cytokine IL-4 using ELISA. As given in Figure 6A, B, the IFN-γ and IL-4 levels increased gradually and nonspecifically after the immunization in all groups. No difference was observed for the IFN-γ level between the different experimental groups. In contrast, the IL-4 levels in the DT390-Cap groups were slightly higher than that in the corresponding Cap alone groups with or without ISA201 at 21, 28, and 42 dpi.
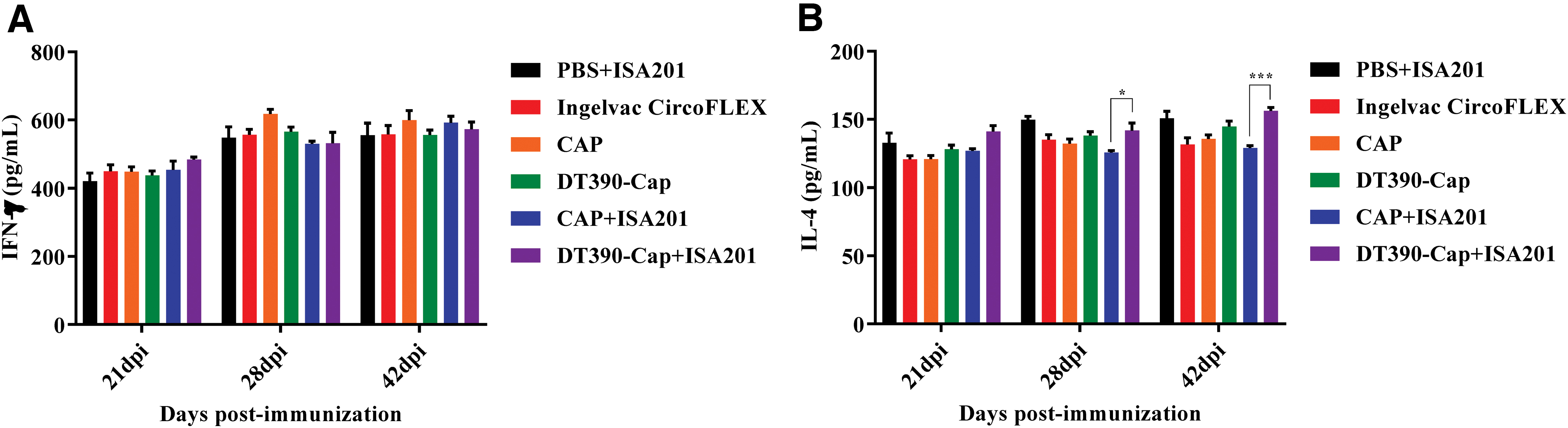
Determination of cytokine levels using mouse cytokine ELISA kit. The serum samples were collected from the immunized mice at 21, 28, and 42 dpi. The murine IFN-γ
DT390-Cap provided improved protection against PCV2 challenge
All immunized mice were challenged using 0.85 mL of 106.5 TCID50/mL PCV2 SH strain using intraperitoneal injection (0.5 mL), subcutaneous injection (0.3 mL), and nasal drops (0.05 mL) at 28 dpi. The viral loads were examined at 42 dpi (14 dpc). Based on previously published literature (4,26), the viral load was well detected at 7, 14, and 21 dpc. Therefore, we selected 14 dpc to detect the viral load in this study. The lungs, liver, and thymus were collected, and the genomes were extracted to assess the expression of the PCV2 Cap gene using real-time PCR. As given in Figure 7A, the viral loads of all immunized groups were significantly lower than that of the PBS+ISA201 negative control group, in the lungs. The viral loads in the liver and thymus of DT390-Cap groups with or without ISA201 were lower than those of the corresponding Cap alone groups (Fig. 7B, C), although the result was not statistically significant because of wide variation. The protection data demonstrated that the engineered DT390 improved the protective immunity of Cap subunit antigen, making it more effective against the PCV2 challenge.
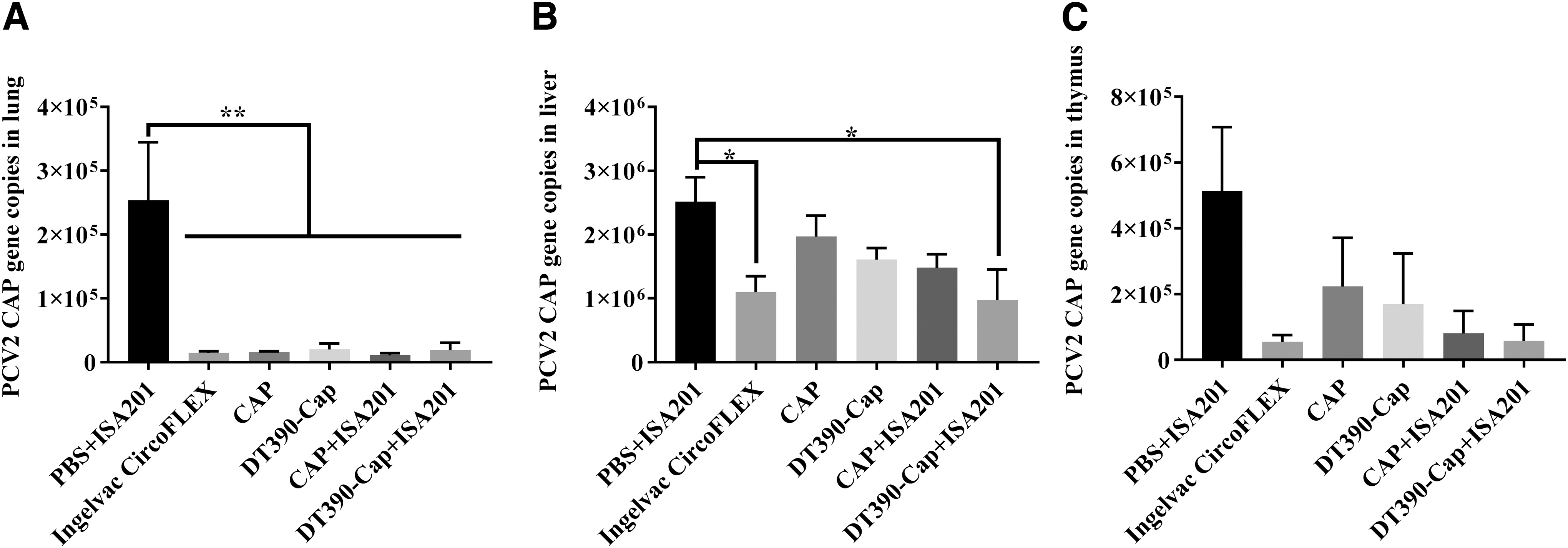
Evaluation of protection in immunized mice, against challenge using strain SH of PCV2. All immunized mice were challenged using 0.85 mL of 106.5 TCID50/mL PCV2 SH strain by intraperitoneal injection (0.5 mL), subcutaneous injection (0.3 mL), and nasal drops (0.05 mL) at 28 dpi. The viral loads were detected at 42 dpi (14 dpc). Samples were collected from the lungs
Discussion
The diphtheria toxin mutant CRM197 has been widely used as a highly effective and approved carrier protein for commercial conjugate vaccines against infectious diseases (14). However, the antigen and carrier protein require separate production, followed by chemical conjugation. This increases the manufacturing cost and leads to variations between batches. In this study, we aimed to circumvent these problems by generating a vaccine candidate in which PCV2-Cap was fused to truncated diphtheria toxin DT390, and the fusion protein was expressed using the diphtheria toxin-resistant P. pastoris expression system. This is a novel approach for using the engineered DT390 as a carrier to improve the immunogenicity and protective immunity of the subunit vaccines.
P. pastoris has been widely used for expressing recombinant proteins, including diphtheria toxin-based immunotoxins/fusion toxins (2,20,25,28). The P. pastoris expression system has the advantages of being inexpensive, high-yield, and scalable for industrial production (29). DT390-Cap was expressed using the unique diphtheria toxin-resistant P. pastoris expression system. The production level was high and purification quality was excellent. These advantages will facilitate the clinical development and commercial success of this vaccine candidate. In addition, the expressed DT390-Cap was N-linked glycosylated. It is known that glycosylation affects the antigenicity and immunogenicity of the vaccine. We speculated that this N-linked glycosylation might also contribute to the improved immunogenicity of DT390-Cap.
The commercial Cap subunit vaccine used in our control group was produced by expressing Cap protein with recombinant baculoviruses in insect cells. The sonicated lysate was directly used as the vaccine. In contrast, highly pure DT390-Cap was obtained by purification with Ni-Sepharose fast flow resin and the anionic exchange resin Poros 50HQ. We speculate that our two-step purified DT390-Cap will have better safety profile as a subunit vaccine candidate.
In this study, 50 TCID50 of PCV2 was used to determine the neutralizing antibody titer. We selected 50 TCID50 instead of 100 TCID50 based on a pilot experiment performed before the formal assays. We observed that 100 TCID50 of PCV2 was so high that the twofold dilution of the serum of the DT390+ISA201 group at 21 and 28 dpi could not neutralize it and the neutralizing antibody titer could not be obtained. In contrast, 50 TCID50 worked well. Therefore, we eventually selected 50 TCID50.
DT390-Cap mainly elicited a humoral response, as reported for other Cap-based subunit vaccines (32). An important index of the cellular immune response is cytokine secretion by Th1 and Th2 lymphocytes. Therefore, we monitored the levels of Th1-associated IFN-γ and Th2-associated IL-4 in the serum of vaccinated mice. The results showed that DT390-Cap did not induce more IFN-γ production than Cap alone. However, DT390-Cap induced slightly higher IL-4 production than Cap alone at 28 and 42 dpi. This indicated that DT390 might enhance Th2-based cellular immune response of Cap to a certain degree.
As given in Figure 4, we did not subtract the nonspecific background from the Cap-specific antibody titer detected using ELISA. The nonspecific background was observed in all groups even at 3 days before the immunization. This is the reason for the apparent inconsistency between the antibody titers measured using ELISA and neutralizing antibody assay. Moreover, the titers were consistent when the background values were subtracted from the results of the specific antibody assay. The ELISA had to be titrated more thoroughly to remove the nonspecific background values. Alternatively, we needed to subtract the background values before reporting the ELISA results.
We have noticed the discrepancy between the antibody titers (given in Figs. 4 and 5) and the gene copy numbers in lung samples (given in Fig. 7). No difference was observed for the gene copy numbers in the lung samples among the experimental groups except for that in the negative control group, the underlying reason for which remains unknown (Fig. 7). In contrast, in the liver and thymus, the protection (gene copy numbers in Fig. 7) exhibits an inverse correlation with the antibody titers (given in Figs. 4 and 5). As given in Figure 7, the highest viral load was detected in mouse liver, as previously reported by our laboratory (23). Therefore, the liver might be the preferred target for PCV2 replication and the indicator for best protection.
Our immunogenicity study using mice has two limitations. First, we directly used the maximal tolerated dose of DT390-Cap without optimizing the DT390-Cap dose in mice. It is necessary to optimize the DT390-Cap dose. Second, we did not include the DT390 control group.
Two pilot studies for immunogenicity without challenge were performed and comparable humoral immune response was obtained in mice (data not shown). Owing to restrictions in the animal facility, in vivo immunogenicity and protective immunity were assessed using mice, and not pigs. In the future, we will assess the in vivo immunogenicity and protective immunity of DT390-Cap using pigs as the target animals. We also plan to develop a new version of DT390-based Cap fusion protein, DT390G53E-Cap, after removing the possible nonspecific toxicity of the DT390 domain. Furthermore, we intend to fuse two or more copies of the Cap gene to DT390, to further enhance the immunogenicity and protective immunity of the Cap subunit vaccine.
Conclusions
In this study, we genetically engineered PCV2-Cap fused to the truncated diphtheria toxin DT390 and expressed the DT390-Cap fusion protein using the diphtheria toxin-resistant P. pastoris expression system. DT390-Cap was significantly more effective than Cap alone in eliciting Cap-specific antibodies, neutralizing antibodies, and protective immunity against PCV2 infection in mice.
Footnotes
Acknowledgments
The authors thank Angimmune LLC for kindly providing the diphtheria toxin-resistant P. pastoris strain and the codon optimized DT390 DNA. The authors also acknowledge Prof. Jiang Ping, Nanjing Agricultural University for providing us the PCV2 SH strain.
Ethical Approval
This study was approved by the animal ethical committees of Shanxi Agriculture University, Shanxi, China.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the Key Research and Development Program of Shanxi Province, China (Grant No. 201903D421037).