
Abstract
Toll-like receptors (TLRs) represent the immune link between the innate and the adaptive immune signals against various pathogens. This study aimed to evaluate the TLRs3 and 7 as immune-markers in differentiating between hepatitis C virus (HCV)-infected and -uninfected patients. Also, the use of the TLR3 and TLR7 as immune markers was compared with the prevalent bio and immune markers for autoimmune diseases in HCV-infected or -uninfected patients. The levels of GPT, GOT, B cell activated factors, tumor necrosis factor-alpha (TNF-α), and interleukin (IL)-10 were measured in plasma, while the levels of TLR3 and TLR7 were quantified in lysates of peripheral blood mononuclear cells from healthy donors, HCV-infected patients, nonalcoholic fatty liver (NAFL) patients without autoimmune diseases and with autoimmune diseases (HCV-infected patients with autoimmune diseases [HCV+auto], nonalcoholic fatty liver patients with autoimmune diseases [NAFL+auto]), rheumatoid arthritis (RA), and systemic lupus erythematosus (SLE) patients. The relative expression of TLR3, TLR7, TNF, and IL-10 in cell lysates was assessed against glyceraldehyde 3-phosphate dehydrogenase (GAPDH) by quantitative real time-polymerase chain reaction (qRT-PCR). Results showed that TLRs 3 and 7 levels were significantly higher in SLE, RA, HCV, HCV+auto, and the NAFL patients compared to the normal control. The cell lysates from SLE patients expressed TLR3 at relatively significantly higher mRNA levels compared to normal subjects or other patient groups. The NAFL+auto patients expressed TLR7 at relatively significantly high mRNA levels compared to normal subjects or other patients. The RA patients expressed TLR7 at relatively significantly higher mRNA levels when compared to HCV, HCV+auto, and NAFL+auto patients. Conclusions: At the protein level, TLR7 can differentiate between HCV and NAFL patients. In addition, both TLRs3 and 7 can serve as potent markers in differentiating between NAFL and NAFL+auto.
Introduction
Hepatitis C virus (HCV) is one of the major global causes of death and morbidity (2,12). Chronic HCV infection affect around 130–150 million of the world's population (2). A high prevalence of chronic HCV infection has been reported in African and Eastern Mediterranean countries, with a prevalence of 14.7% in the population of Egypt (3,20,31). The word hepatitis refers to liver inflammation. Most forms of hepatitis result from viral infection, although in some cases, it is caused by an autoimmune disorder, in which the body's immune system attacks liver cells because it will become unable to differentiate between harmful invaders and healthy liver tissue (21). The new HCV-specific antiviral drugs aim to inhibit the viral RNA replication and causing its eradication by targeting the enzymes involved in its replication (19,35). However, some liver inflammations of undetectable viral etiology occur. In addition, liver damage caused by alcohol, toxins, and certain drugs is mediated by inflammation (21). Moreover, some inherited diseases can cause inflammation and hepatitis, along with prolonged obstruction of bile flow (21). Some forms of liver inflammation produce mild symptoms, while others can be serious or life threatening (21). Thus, differentiation between liver inflammations due to various etiologies still represents big challenge. Therefore, we aim to develop new marker(s) that successfully distinguish between HCV-mediated and nonviral hepatitis.
Immune response to pathogens depends on toll-like receptors (TLRs) to recognize specific molecular products, which lead to acute immune responses to kill the pathogen. Also, TLRs control induction of adaptive immunity by stimulating dendritic cell maturation (41). The identification of TLR signaling pathways may unravel molecular mechanisms of self-tolerance and the development of autoimmunity (50).
In the liver, TLRs recognized molecular signature in autoimmune liver disease, alcoholic liver disease, nonalcoholic steatohepatitis, fibrogenesis, and chronic hepatitis B virus (HBV) and HCV infections (9). TLRs-based therapies may represent a promising approach in the treatment of viral hepatitis (9). Sghaier et al., (44) demonstrated that TLRs 3\4 polymorphisms are promising biomarkers of liver cirrhosis and cancer associated with HBV and HCV infection.
TLRs 3\7 recognizes viral RNA and subsequent increase in IFN-α transcription. Mohamed et al., (33) demonstrated the lower expression of TLRs 3\7 in HCV patients. TLR7 recognizes HCV genome (14). Immune modulation of TLRs is recommended as new therapeutic tool for HCV (45). Zhang et al., (54) demonstrated that novel anti-HCV vaccination strategies should target the induction of TLR7/8 in antigen presenting cells. The antiviral effects of TLR3 signaling on viral hepatitis are likely mediated via stimulating a variety of cells to produce type I IFN that inhibits HCV or HBV replication (23). TLR3 can directly sense HCV infection in human hepatocytes to suppress HCV replication (10). Interestingly, TLR3 signaling is suppressed during viral hepatitis, which in turn may contribute to the virus escape from the surveillance of the innate immunity (54).
Sugiura et al., (46) suggested the criteria of autoimmune hepatitis (AIH): positive anti-HCV antibody, high titer of antinuclear antibody (ANA) in addition to elevate serum alanine aminotransferase (ALT), and HCV-RNA expression. TLR3 and TLR7 play a role in the pathogenesis of a variety of autoimmune diseases (51). The expression of TLR3 was markedly increased in AIH (36). The incidence of nonalcoholic fatty liver disease (NAFLD) and AIH has some similar laboratory and histological findings (47).
Controversially, the downregulation of TLR3 was shown to ameliorate the steatosis in mice, which suggests its potential effect as immunotherapeutic target for the treatment of hepatic inflammation and steatosis (29). Similarly, the impact of TLR7 in preventing progression of NAFLD was previously proved both in vivo and in vitro (27).
The present research focused on evaluating the potential impact of TLRs 3\7 as immune markers to differentiate between HCV-infected and non-HCV-infected patients in presence or absence of autoimmune diseases.
Materials and Methods
All procedures performed in the study were in accordance with the ethical standards or practice of the institution, at which the study was conducted. Collection of human blood samples was approved by the Medical Ethics Committee of the National Research Centre in Egypt (Registration number of the ethical approval: 17159).
Subjects and samples collection
Blood samples were collected from October 2016 to May 2019 (The Medical Unit at the National Research Center). Eighty-one samples used throughout the study were divided into seven groups: (1) 9 healthy control volunteers with no history of HCV-infection, liver disturbance diseases, or autoimmune diseases (normal), (2) 11 HCV-infected patients (HCV), (3) 11 HCV-infected patients who are positive for autoimmune markers (HCV-infected patients with autoimmune diseases [HCV+auto]), (4) 4 HCV\HBV noninfected patients but had liver disturbance as nonalcoholic fatty liver (NAFL), (5) 6 HCV\HBV-noninfected patients, but had liver disturbance as NAFL and are positive of autoimmune marker (nonalcoholic fatty liver patients with autoimmune diseases [NAFL+auto]), (6) 19 patients suffering from rheumatoid arthritis (RA), and (7) 21 systemic lupus erythematosus (SLE) patients.
Serum was separated from 10 mL of individual whole blood samples and kept in aliquots at −80°C until use in the subsequent assays. Peripheral blood mononuclear cells (PBMCs) were separated from the anticoagulated blood using the lymphoprep™ reagent (LYS-3772; Fresenius KabiNorge, Oslo, Norway) according to Boyum (8).
The PBMCs were subjected for four sonication rounds to release their protein contents. The lysates were further frozen to −80°C and thawed to room temperature for three successive times each for 2 min. Then, the lysates were later subjected to centrifugation at 1,500 g for 10 min. The supernatants were harvested, aliquoted, and frozen till being assayed.
Detection of liver enzymes
The liver enzymes AST and ALT were assayed by an automated kinetic method at λ max 340 nm using commercial detection kits (Diamond Diagnostics, Cairo, Egypt) in sera of normal, HCV, HCV+auto, NAFL, NAFL+auto, SLE, and RA subjects according to the method of Reitman and Frankel (43).
Diagnosis of HCV and HBV
Antibodies to HCV and HBV antigens were detected in sera from normal, HCV, HCV+auto, NAFL+auto, SLE, and RA subjects by commercial rapid tests (Ref.Lo31-10331-Imported by Lab Dimension Co. LCA7656–01. A CON Laboratories, Inc., 10125 Mesa Rim Road, San Diego, CA. Presence of HCV antibodies was confirmed by commercial ELISA kit (Prechek Bio, Inc., California).
Indirect immune-fluorescent assay
Indirect immune-fluorescent assay (IFA) was carried out on slides of mouse tissue (kidney, liver, stomach) to evaluate antismooth muscle antibodies (ASMA), antimitochondrial antibodies (AMA) detected in the sera from healthy, HCV-infected and fatty liver noninfected-HCV-patients (IFA System, NOVA Lite ANA KSL, INOVA Diagnostics). ANAs were detected in the sera from healthy, HCV and NAFL patients on human cell line (HEP-2) according to the NOVA Lite RHEP-2 ANA Kits/Substrate Slides. Positive reactions of ANA, ASMA, and AMA were recorded according to established criteria.
Detection of BAFF by ELISA
Human BAFF/BLyS in plasma from normal, HCV, HCV+auto, NAFL, NAFL+auto, SLE, and RA subjects was determined using the Quantikine BAFF/BLyS/TNF134B ELISA kit (
Quantification of interleukin-10, tumor necrosis factor-alpha, TLR3, and TLR7 using ELISA
The levels of interleukin (IL)-10 and tumor necrosis factor-alpha (TNF-α) in plasma, TLR 3, and TLR 7 in cell lysates from PBMCs were detected by ELISA as described by Engvall and Perlmann (16) with some modification. Microplate was coated by 50 μL\well of 1:50 diluted plasma or cell lysates prepared from 105 cells/mL from normal, HCV, HCV-auto, NAFL, NAFL+auto, SLE, and RA patients and incubated overnight at room temperature. Diluted anti-TNF-α antibody (rabbit polyclonal to TNF alpha; Abcam, Cambridge, UK), anti-TLR3 antibodies, or anti-TLR7 antibodies (Abcam) were added. Peroxidase conjugated goat anti-rabbit IgG H&L (Abcam) was used at 1:1,000 dilute (50 μL\well). Levels of human IL-10 were detected by ELISA kits from (KOMA BIOTECH INC). The concentrations of IL-10 were calculated based on experimental standard curves. The changes in OD were recorded at λ max 492 nm using a multi-well plate reader (Tecan; Sunrise GmbH).
Expression of TNF-α, IL-10, TLR3, and TLR7 using quantitative real time-polymerase chain reaction
Total RNA was extracted from 200 μL each individual cell lysate of normal, HCV, HCV+auto, NAFL, NAFL+auto, SLE, and RA patients using the PureLink® RNA Mini Kit (Invitrogen, Carlsbad, CA) according to the manufacture's protocol and was quantified using a nano-drop photometer (Implen, Munich, Germany). Equal concentration (100 ng) of total RNA from each cell lysate was mixed with oligo dT primer to reverse transcribe it into cDNA (Omniscript RT kit; Qiagen, Hilden, Germany) in presence of reverse transcriptase enzyme mixture containing buffer, dithiotheritol, and deoxynucleotide triphosphate. To amplify individual genes, specific primers for TLR3, TLR7, TNF-α, and IL-10 were used and the glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was detected as a house keeping gene (Table 1). Quantitative real time-polymerase chain reaction (qRT-PCR) was performed using 250 ng sample cDNA and QuantiTect SYBR Green (Qiagen) in a Rotor Gene Q Real-Time PCR machine (Qiagen). The amplification program was as follows: 95°C for 15 min; 40 cycles: 94°C for 15 sec, 50–58°C for 30 sec, and 72°C for 30 sec. Levels of mRNA expression of the studied genes were normalized to mRNA expression of GAPDH using the 2−ΔCt cycle threshold calculation.
List of Designed Primers Used for Quantitative Real Time-Polymerase Chain Reaction
Statistical analysis
Data were analyzed using GraphPad Prism Software version 8.0.2 (La Jolla, CA). Statistical significance was assessed using student's t-test. Individual data points were also presented as scattered plots to demonstrate the range of recorded individual variations, which may influence standard division and standard error.
Results
Immunological investigations
Level of anti-HCV antibodies
HCV-Ab titers in sera from HCV, HCV+auto, NAFL, NAFL+auto, SLE, RA, and healthy individuals were (0.656 ± 0.449), (0.644 ± 0.556) and negative result for the other groups, respectively.
The titer was significantly higher (p < 0.05) in both HCV and HCV+auto patients compared to the rest of the studied human groups.
Level of autoimmune markers
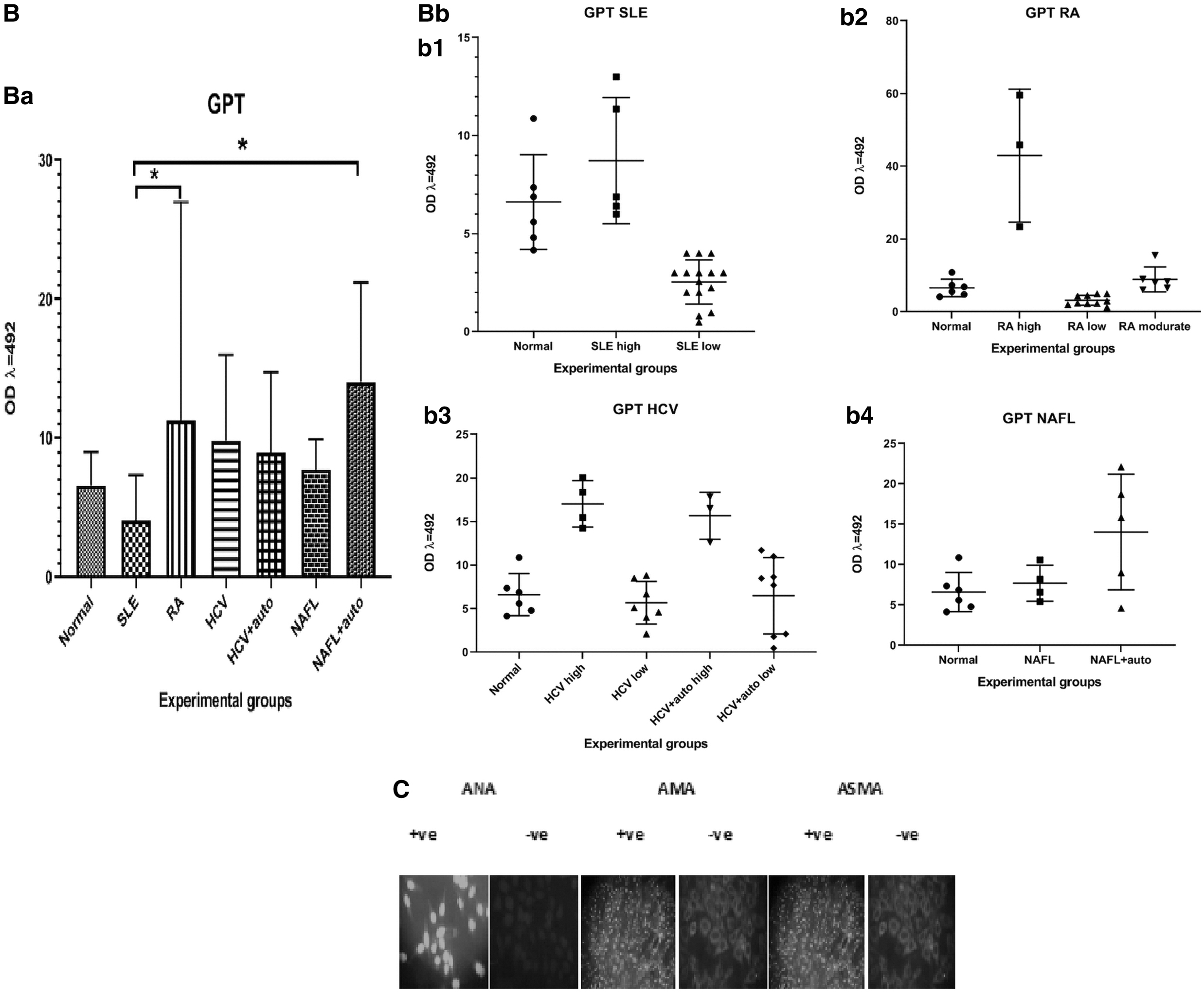
Among the HCV patients, the mean percentage of positive reactions of the ANA, AMA, and ASMA were 40%, 0%, and 32%, respectively. NAFL patients showed percentage positive reactions for ANA, AMA, and ASMA equal to 50, 0, and 30, respectively (Fig. 1C).

Level of BAFF
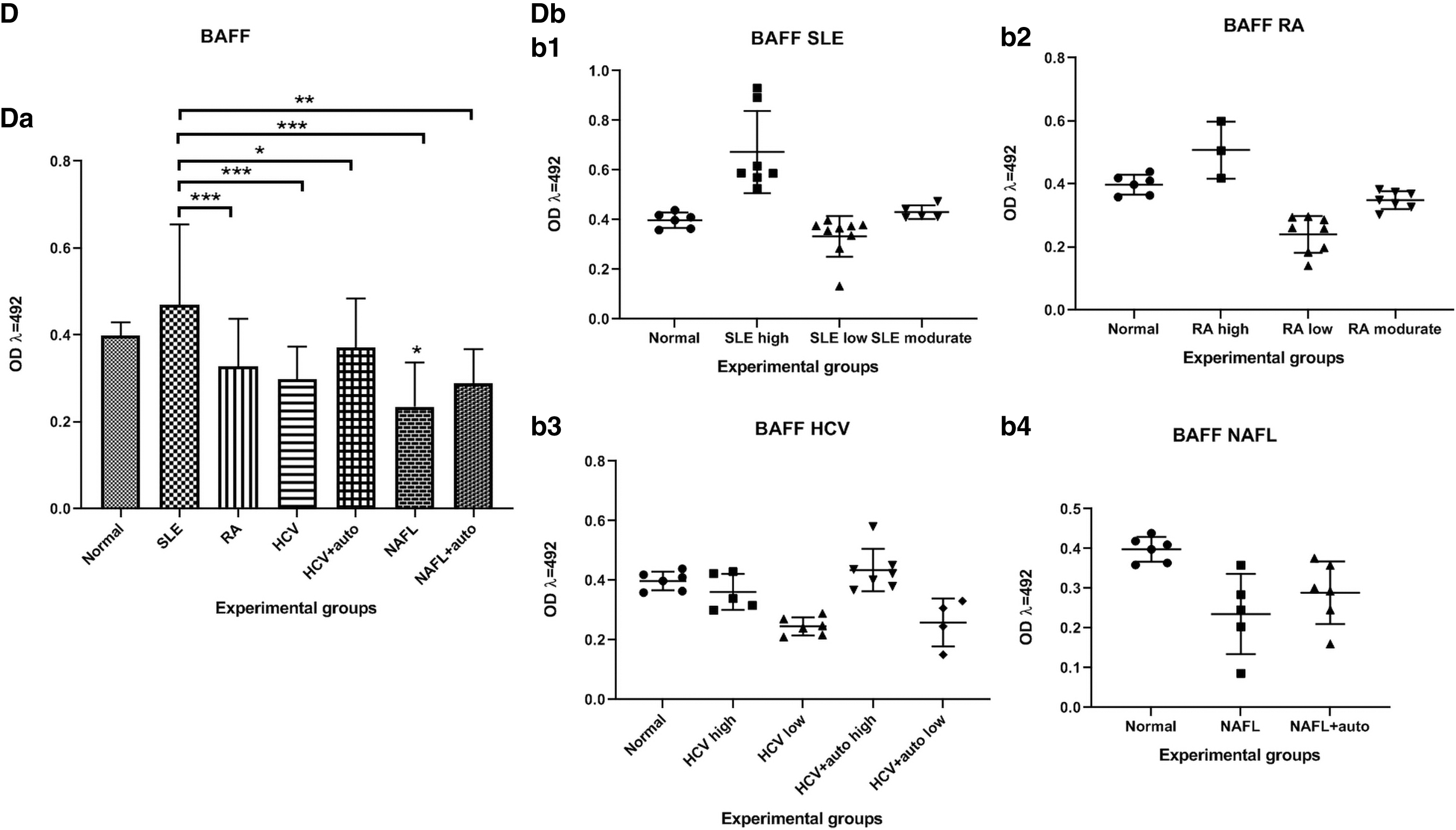
The BAFF level was significantly lower (p < 0.05) among the NAFL patients compared to normal controls. It was also significantly lower (p < 0.001, 0.001, 0.05, 0.001, 0.01) among the RA, HCV, HCV+auto, NAFL, and NAFL+auto patients, respectively, compared to the SLE patients (Fig. 1D).
Levels of IL-10 and TNF-α proteins
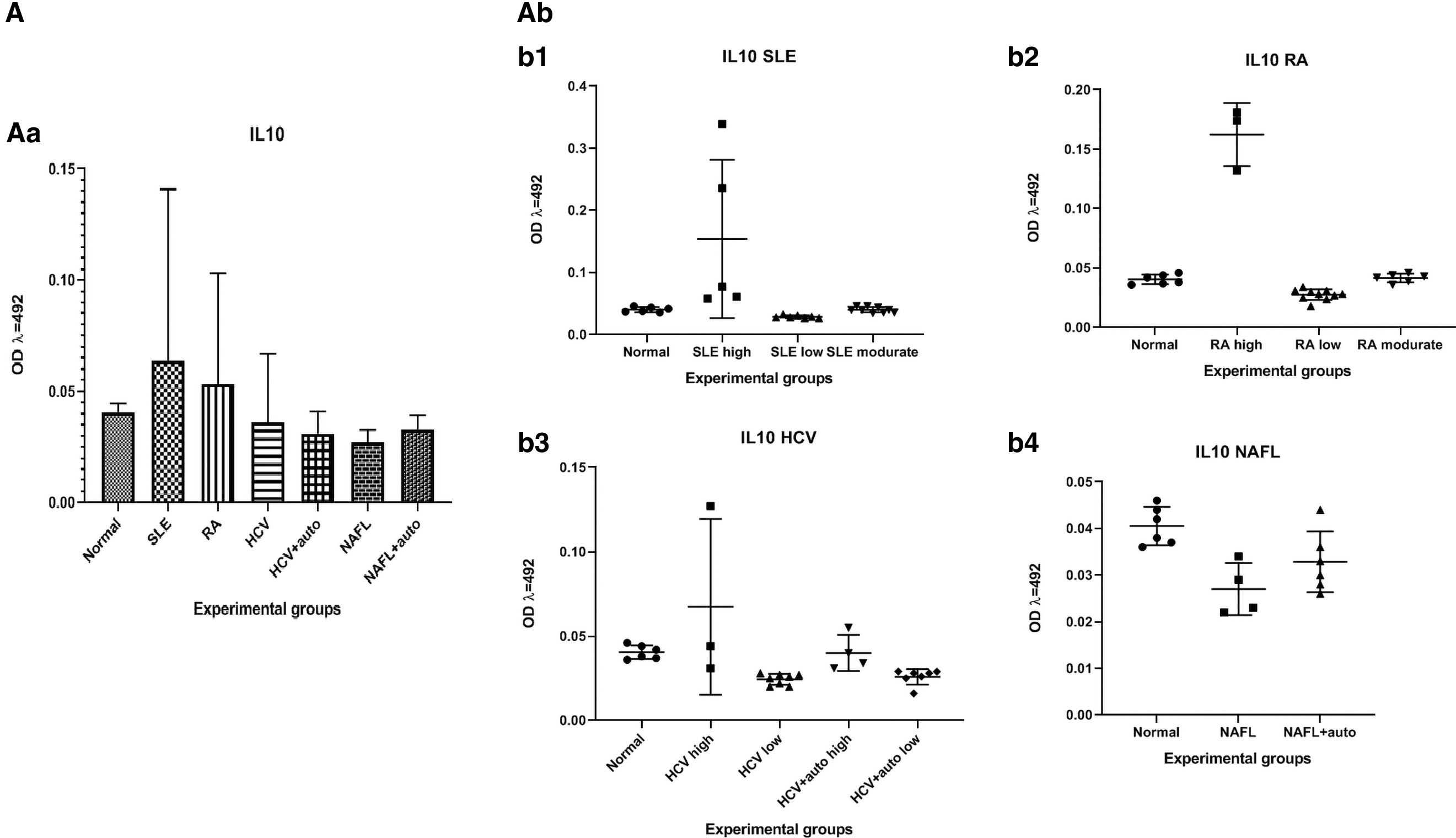
The IL-10 level in plasma showed no significant differences between various patients' groups or between each of the patients' groups and the normal controls (Fig. 2A).
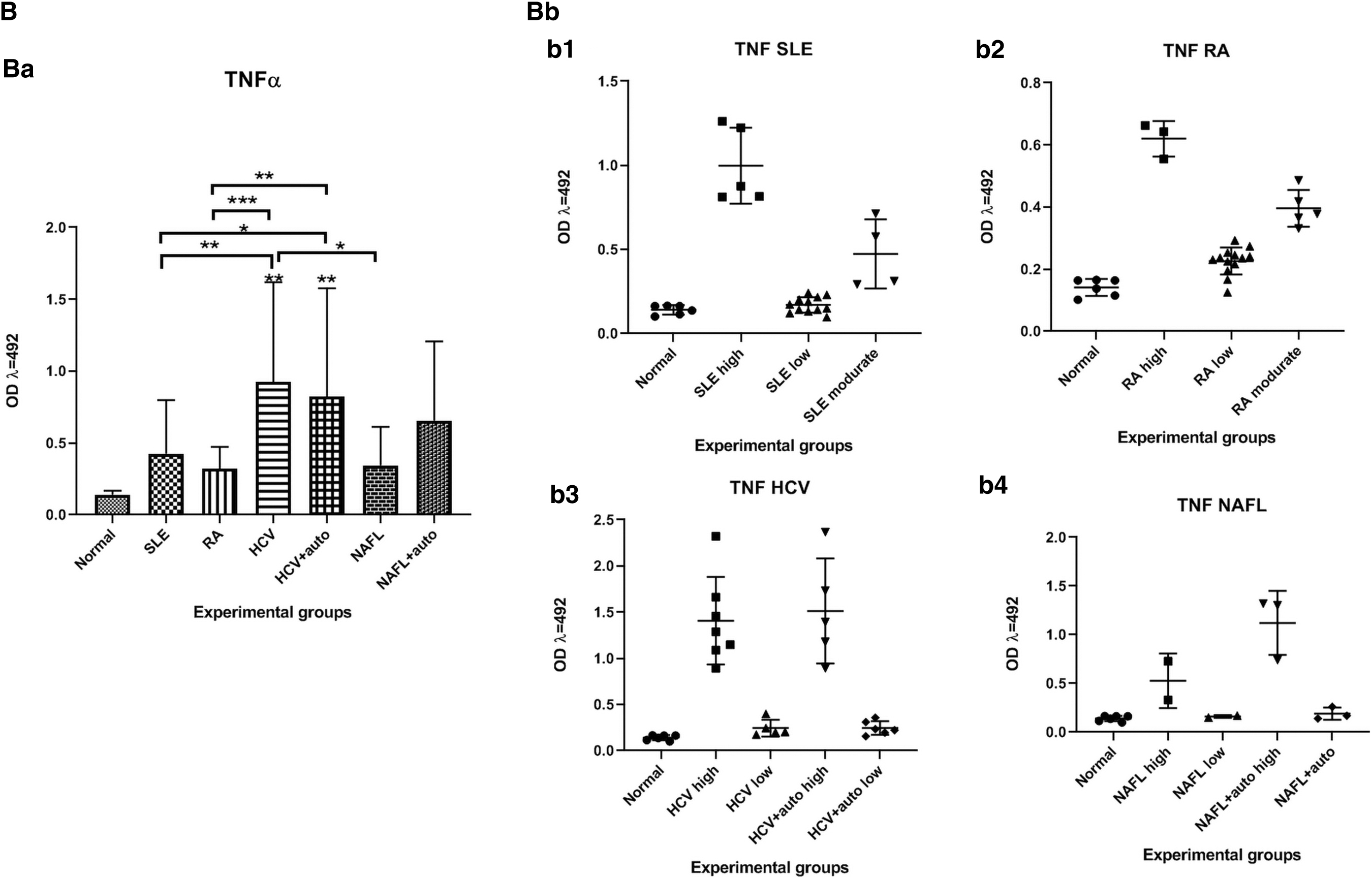

The TNF-α level was significantly higher (p < 0.01, 0.01) among both the HCV and the HCV+auto patients compared to normal control. Also, the levels were significantly higher (p < 0.01, 0.05) among the HCV and HCV+auto patients compared to the SLE ones. The TNF-α level was significantly higher (p < 0.001, 0.01) in both the HCV and HCV+auto patients compared to the RA patients. The TNF-α level was significantly lower (p < 0.05) between HCV and NAFL (Fig. 2B).
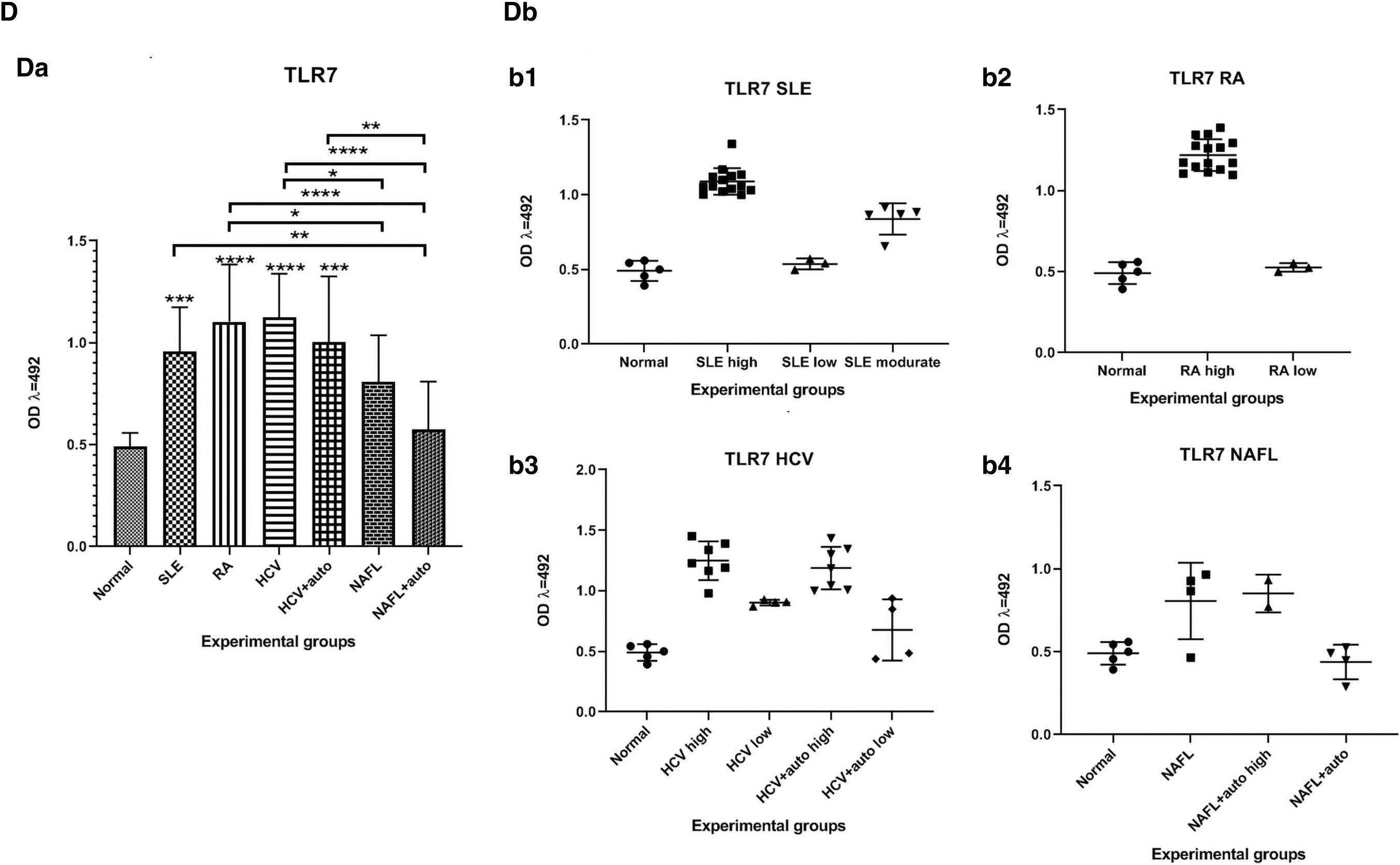
Levels of TLR3 and TLR7 proteins
The TLR3 levels were significantly higher (p < 0.05, 0.001, 0.001, 0.01, 0.05) among the cell lysates from SLE, RA, HCV, HCV+auto, and the NAFL patients, respectively, in comparison to the normal. The lysate from NAFL+auto patients showed no significant changes in the levels of TLR3 compared to the normal level. Also, TLR3 levels were significantly higher (p < 0.05) in lysates from the RA, HCV, and NAFL+auto patients compared to the level in lysates from SLE patients. The TLR3 levels were significantly higher (p < 0.001, 0.001, 0.01, 0.05) in cell lysates from the RA, HCV, HCV+auto, and NAFL patients, respectively, compared to the level from NAFL+auto patients (Fig. 2C).
The TLR7 levels were significantly higher (p < 0.001, 0.0001, 0.0001, 0.001) in cell lysates from the SLE, RA, HCV patients, and the HCV+auto patients, respectively, in comparison to normal group. The level of TLR7 in cell lysates from NAFL+auto and NAFL patients showed no significant changes compared to normal. Also, its levels were significantly higher (p < 0.01) among the NAFL+auto patients compared to the SLE patients. The TLR7 levels were significantly higher (p < 0.05, 0.0001) among the cell lysates from both the NAFL and NAFL+auto patients compared to RA patients. The TLR7 levels were significantly higher (p < 0.05, 0.0001) among lysates from both the NAFL and the NAFL+auto patients compared to the HCV patients. Furthermore, the TLR7 levels were significantly higher (p < 0.01) from the HCV+auto subjects compared to NAFL+auto patients (Fig. 2D).
Biochemical assessment
Level of liver enzymes
The GPT level in plasma from normal, HCV, HCV+auto, NAFL, NAFL+auto, SLE, and RA patients showed no significant differences between various patients' groups or between each of the patients' groups and the normal (Fig. 1B). GOT levels were significantly higher (p < 0.01) in the RA patients compared to normal (Fig. 1A).
Molecular analysis
Levels of TNF-α and IL-10 mRNA expression
The cell lysates from SLE patients expressed IL-10 at relatively significantly higher (p < 0.05) mRNA levels compared to normal subjects, whereas the cell lysates from HCV, HCV+auto, NAFL, NAFL+auto, and RA patients did not show difference in the expression profile of the TNF-α mRNA compared to normal ones (Fig. 3A). The cell lysates from NAFL+auto patients expressed TNF-α mRNA at significantly higher (p < 0.0001, 0.0001, 0.0001, 0.0001, 0.01) levels compared to normal group, HCV+auto, NAFL, SLE, RA, and HCV patients, respectively (Fig. 3B).
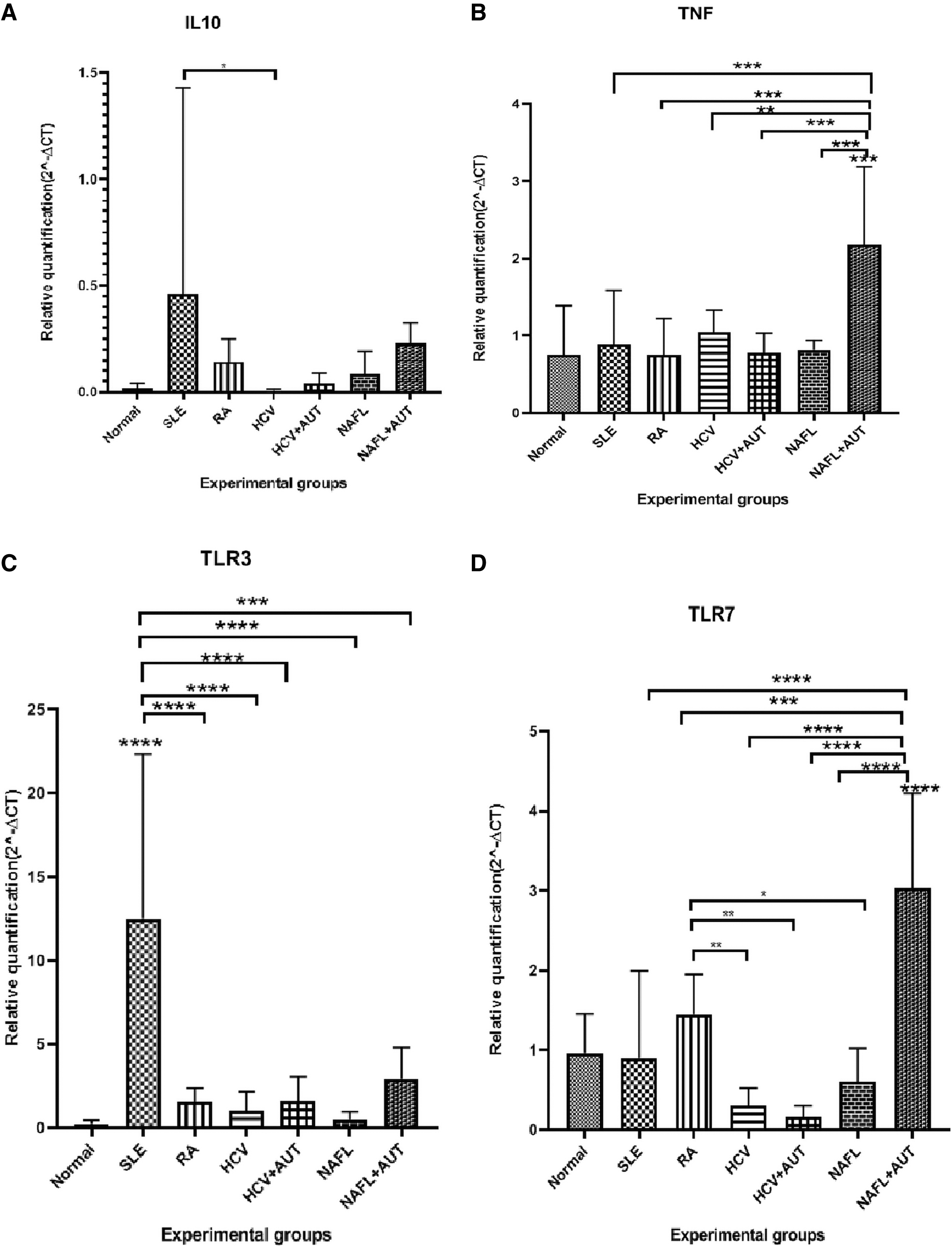
The expression of IL-10
Levels of TLR3 and TLR7 mRNA expression
The cell lysates from SLE patients expressed TLR3 mRNA at significantly higher (p < 0.0001) mRNA levels compared to normal control, HCV, HCV+auto, NAFL, NAFL+auto, SLE, and RA patients (Fig. 3C).
The cell lysates from NAFL+auto patients expressed TLR7 mRNA at significantly higher (p < 0.0001) mRNA compared to normal, HCV, HCV+auto, NAFL, SLE, and RA patients (Fig. 3D). The cell lysates from RA patients expressed TLR7 mRNA at significantly higher (p < 0.001) levels compared to HCV, HCV+auto, and NAFL patients (Fig. 3D).
Discussion
In the present study, three issues are illustrated. First, the ability of TLR3 or TLR7 to differentiate between HCV and non-HCV patients. Second, the affectivity of TLRs 3\7 to be used as new predictive immunodiagnostic markers for the conversion of liver diseases into autoimmune disorders. Finally, whether the currently available and known bio- or immune-markers are efficient enough to solve these issues, consider the simple known autoimmune markers: ANA, AMA, and ASMA. It is well known that among the HCV patients, the mean percentage of positivity rates of ANA, AMA, and ASMA were 40%, 0%, and 32%, respectively. Meanwhile, patients with NAFL exhibited positivity rates of ANA, AMA, and ASMA equal to 50%, 0%, and 30%, respectively. Himoto et al., (22) and Keppeke et al., (25) had the same results as ours; 10–40% of NASH and HCV patients have ANAs. Our study involved liver enzymes as simple biomarkers. Interestingly, the GOT levels was significantly elevated only in RA patients but not in other patient groups compared to normal. Similar findings were also obtained in patients with RA and non-RA as explained by Dimov and Popov (15). Noteworthy, elevation of liver enzymes during the treatment of patients with RA is an important factor which must be considered. Consistently, Curtis et al. (13) demonstrated that 14–35% of patients with RA had abnormal liver enzyme levels during therapy.
Regarding the anti-HCV drugs, they have been shown to improve the levels of liver enzymes (18,46). This interprets and illustrates the significantly decreased liver enzyme levels recorded among the HCV, NAFL patients in our study. Furthermore; significant increase in the anti-HCV-Ab titer was detected in HCV and HCV+auto patients compared with normal subjects, NAFL, NAFL+auto, SLE, and RA patients. Actually; anti HCV-Ab titer can well differentiate between HCV and non-HCV-infected patients, but it cannot predict the conversion of HCV infection to AIH, whereas a liver damage in HCV patients was more than in AIH with similar histopathological observations (18,46).
The current results showed that the BAFF level was significantly lower in NAFL patients compared to normal ones. Controversially, Puri and Sanyal (42) disagree with this result and demonstrated that patients with NAFLD had higher serum BAFF levels. BAFF level was significantly lower in NAFL>HCV>RA>NAFL+auto>HCV+auto patients compared to SLE patients. Also, a significantly higher BAFF level was recorded in the HCV+auto patients than in the NAFL subjects. Unfortunately, HCV infection is characterized by multiple B cell disorders; leading to a decrease in the count of B cell with concomitant increase in the BAFF levels (28). High serum levels of BLyS/BAFF with acute HCV can predict its evolution to chronic infection (48) and associated with autoimmunity (49). The treatment may be the reason of decreasing the BAFF level among the RA, HCV, HCV+auto, NAFL, and NAFL+auto patients compared to SLE.
Accordingly, the immunological assessment revealed no significant change in the IL-10 level at the protein level in the NAFLD patients, compared to the normal ones. In addition, the mRNA levels of IL-10 levels were significantly upregulated in the cell lysates from SLE patients, compared to HCV patients. However, Borai et al. (7) reported that severe NAFLD patients showed significant decrease in IL-10 levels. Moreover, AbdElazeem, et al., (1) demonstrated that the circulating IL-10 concentrations were significantly elevated in SLE patients.
The current results showed that the TNF-α level was significantly higher among the HCV>HCV+auto>NAFL+auto patients compared to normal subjects. Similar findings were also obtained in patients with HCV infection as explained by Nelson et al., (37). The TNF-α level was significantly higher among the HCV or HCV+auto patients compared to NAFL patients. So, TNF-α level showed no specific diagnostic signature among tested groups. On the contrary, the cell lysates from the NAFL+auto patients expressed TNF-α mRNA at significantly higher levels compared to normal, HCV+auto patients, NAFL, SLE, RA, and HCV patients. TNF-α is mainly involved in inflammatory immune responses (38), and its overexpression occurred in case of autoimmunity (26). Both TNF-α and IL-1β regulate the level of IL-10, which influence type I IFN secretion. Therefore, IL-10 appears to play a central role in regulating production of cytokines induced by HCV core protein (40). TNF-α released mainly by the mononuclear cells affects all liver cell types and orchestrates production of many other mediators relevant in chronic liver diseases (39). TNF-α is thought to be a mediator in the development of viral hepatitis (17). Moradi et al., (34) demonstrated that the levels of IFN-I, IL-10, and TNF-α were overexpressed in HIV, HCV, and HIV/HCV coinfected patient. Bocsan et al., (5) demonstrated that TNF-α levels were significantly higher in NAFL patients compared to control with negative correlation with IL-10. We observed the same results in NAFL patients who were positive in autoimmune markers both at the protein and mRNA levels. There was a prominent imbalance between TNF-α and IL-10 in patients with NAFLD (5). The positive reactivity of the NAFL patients for the autoimmune markers observed in our results represents a great addition to the previously obtained results recorded in other studies.
TLRs give the innate immune response a considerable specificity of pathogens (6,32,53). HCV core protein interacts with TLRs on monocytes, inducing them to produce cytokines (40). Negative regulation of TLRs results in diminished proinflammatory cytokine production and limits excessive pathology (4). Our data postulated no significant change in the TLR3 transcription in both the HCV and HCV+auto patients compared to normal controls; but unlikely, its levels in both groups were significantly downregulated, compared with SLE. Also, there was no significant change in the TLR7 transcription in both the HCV and the HCV+auto patients compared to normal; but significantly decreased when compared with NAFL+auto patients. Moradi et al., (34) demonstrated that TLR3, TNF-α, and IFN-I were correlated negatively with HCV load, and the opposite was seen for both TLR7 and IL-10. Mohammed et al., (33) reported that HCV may induce downregulation of TLR3\TLR7 expression on innate immune cells with subsequent decrease in INF-alpha production as one of the immune evading mechanisms that participates in viral persistence and increasing the expression level of TLRs may help in treatment of HCV infection. TLR3 was significantly downregulated in chronic HCV compared to healthy controls compared to low viral load. The significant reduction in the expression of TLR3 was observed in nonresponders compared to responders after treatment (6). The downregulation of TLR3 expressions in chronic HCV correlates with the disease severity and the outcome of HCV infection (24). Furthermore, TLR3 could be a novel therapeutic target for the treatment of hepatic inflammation and steatosis (29,30,52). The increase in TNF-α during NAFLD promotes the activation of the NLRC4 inflammasome, which increases production of IL-18 and IL-1β and triggers pyroptosis. These exacerbate inflammation and promote disease development (11).
In conclusion, TLR3 was shown to be unable to differentiate between HCV and NAFL patients at both the protein and mRNA levels. Beneficially, TLR3 can differentiate between NAFL and NAFL+auto at the protein level. Importantly, at the protein level; TLR7 can differentiate between HCV and NAFL patients. At the transcription level, the TLR7 can also differentiate between the NAFL and the NAFL+auto patients, since a significantly higher expression of TLR7 in the NAFL+auto patients compared to healthy individuals, HCV, HCV+auto, NAFL, SLE, or RA patients was observed. Last and not least, TLRs 3 and 7 can differentiate between the NAFL and NAFL+auto patients.
Footnotes
Acknowledgments
The authors acknowledge the National Research Center of Egypt for providing all needed facilities and logistics for the work.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The study was supported by the grant number 11010101awarded from the National Research Center of Egypt.