
Abstract
Infectious laryngotracheitis virus (ILTV) is a cause of main respiratory disease of chickens controlled through live attenuated vaccines. To reduce the risk of adverse effects associated with live vaccines, a recombinant vaccine expressing PH-1 domain of viral glycoprotein B was constructed using the pET expression system under isopropylthiogalactoside (IPTG) induction. The potential immunogenicity of recombinant PH-1 (rPH-1) was evaluated in chickens. Eight-week-old specific-pathogen-free chickens were intramuscularly administered two doses of rPH-1, 25 and 50 μg, alone or with a combination of ISA70 adjuvant. The humoral immune responses were determined up to 3 months postvaccination at 2 weeks apart. The T cell proliferation response was determined on day 28 after primary immunization. The vaccinated birds with rPH-1/ISA70 developed higher and constant-specific anti-ILTV enzyme-linked immunosorbent assay (ELISA) antibodies than in those vaccinated with rPH-1 alone. Coinjection of rPH-1 and adjuvant significantly (p < 0.01) increased the T cell proliferation responses. There were no significant differences in eliciting the immune responses in chickens immunized with the higher dose of the antigen than that with the lower dose. The data indicate the immunogenic efficiency of rPH-1 against ILTV. Vaccination with recombinant proteins offers a preventing option to control the ILTV infection and could be a candidate to replace current live vaccines.
Introduction
Infectious laryngotracheitis virus (ILTV), an alpha herpesvirus, causes an acute and highly contagious upper respiratory disease in chickens. After the description of ILT by May and Tittsler in 1925, the first vaccine against poultry pathogens was established. To date, the disease is controlled by the administration of live attenuated vaccines prepared either by chicken embryo origin (CEO) or tissue culture origin (TCO) vaccine (19). The most limitation of these vaccines is the ability of ILTV strains to infect the nervous system and revert to virulence after bird-to-bird passages (14). To overcome the serious problem, viral vector vaccines based on turkey herpesvirus (HVT) and fowlpox virus (FPV) that genetically modified to express the virus glycoproteins have been developed that are used in the limited poultry-producing regions (11,13,26). The HVT-vectored ILT vaccine expresses the ILTV glycoproteins D and I are registered for subcutaneous administration in 1-day-old chicks and in ovo inoculation in 18-day-old embryos. The genes are the mediator for viral entry into the host and its cell-to-cell spread, respectively. Replication of the HVT vaccine virus in the chickens' tissues may explain low levels of induced protection in chickens compared with a conventional attenuated ILT vaccine (20). The FPV-vectored ILT vaccine expresses the ILTV glycoprotein B and UL-32 genes that play a role in viral entry and cleavage and encapsidation of the viral genome, respectively. Both rHVT-LT vaccines and the rFPV-LT vaccine are administrated via the wing-web puncture in 1-week-old birds, or in ovo in 18-day-old embryos, whereas the rFPV-gB vaccine is administered by wing web application (21). Despite the ability of the FPV-ILT vaccine in preventing mortality and reduction of clinical signs, the protective immunity provided by this vaccine is less than that provided by the modified live attenuated vaccines (9).
The CEO and TCO vaccines can exhibit varying levels of residual virulence of ILTV, which can increase after bird-to-bird passage. However, the TCO vaccine has a lower potential for reversion to virulence than the CEO vaccines. The development of new vaccines through genome-based technologies has been in attention in an effort to overcome the limitations of live vaccines. Glycoproteins have important roles in the entry, antigenicity, and pathogenesis of viruses and considered the main target for the production of recombinant vaccines. Glycoprotein B (gB) belongs to the Class III fusogens and is the most conserved component of the complex cell-entry machinery of herpesviruses (22,25). In addition to the fusogenic ability, gB has been identified as the primary target of IgG antibody responses in herpesviruses infections (18). The gB plays a significant role in the induction neutralizing antibodies. Hence, the glycoprotein may be the primary candidate for the development of recombinant vaccines. The multiple domains of ectodomain are critical for the gB functions including fusogenic activity, intracellular signaling, and cytokine expression during the early stage of virus infection (3,18). We, in a previous in silico study (12), showed that the PH-1 domain of ILTV gB has many epitopes that are capable of inducing humoral and cell-mediated immune responses. To evaluate the host responses, gB of an ILTV field isolate was expressed and formulated with adjuvant, and host immune responses after its administration were determined in this study.
Materials and Methods
Virus propagation
A field ILTV isolate (10) was propagated in embryonic chicken lung cells. Infected and uninfected cells were grown under the physiological culture conditions (37°C, 5% CO2, 65% humidity) in Dulbecco's modified Eagle's medium (DMEM) (INOCLON, Iran) supplemented with antibiotics and the appropriate amount of 10% fetal bovine serum (15). Cultures were examined daily for the presence of cytopathic effect.
Viral protein expression
Viral DNA was extracted from infected egg and PCR was performed for amplification of the gB PH-1 domain using the designed primers 5′-AACAGAACATTGCGCCGTAT-3′ and 5′-GGCACTTCAATCCATTTGCT-3′ corresponding to 643 bp. The restriction enzyme sites for the PH-1 DNA sequence screened in NEBcutter V.2 and the BamHI and XhoI restriction sites were introduced to the N and C terminals of the sequence, respectively. To improve the expression of the final construct in pET-28a (+) vector (Novagen), codon optimization in Escherichia coli strain K12 was performed using JCat server. The parameters including rho-independent transcription termination, ribosome binding site, and restriction enzymes cleavage sites were selected. The solubility on the basis of amino acid sequence was determined through protein–sol (
The PCR was carried out at 94°C for 1 min followed by 30 cycles of 94°C for 30 sec, 57°C for 45 sec, and 68°C for 60 sec, and a final extension of 68°C for 10 min. The PCR product was electrophoresed in 1% agarose gels and stained with RedSafe™ dye. The purified PCR product was digested with the restriction enzymes, and inserted into the XhoI-BamHI of pET-28a (+) vector using T4 DNA ligase. The resulting plasmid transformed into competent E. coli DH5α cells using the heat-shock method, which was grown aerobically at 37°C in Luria-Bertani (LB) broth. The integrity of the extracted plasmid was confirmed by DNA sequencing.
A recombinant PH-1 (rPH-1) clone was grown at 37°C in 500 mL of LB medium supplemented with 50 μg/mL kanamycin under agitation at 190 rpm. At an optical density (OD)600 = 0.6, the protein expression was induced by the addition of 0.4 mM isopropylthiogalactoside (IPTG) followed by incubation at 37°C with shaking at 190 rpm for 4 h. The bacterial cells were harvested at 1-h interval by centrifugation at 3220 × g and 4°C for 5 min and resuspended in Tris buffer (50 mM Tris-HCl, 150 mM NaCl, pH 7.0). The cells were sonicated for 5 min (30 sec sonication and 30 sec rest) and the lysate was centrifuged for 15 min. The recombinant proteins were purified by using Ni-NTA affinity chromatography by using Protino® Ni-TED (The Netherlands) following manufacturer's instructions. The purity and yield of recombinant protein were analyzed by 12% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and protein content was determined using the Bradford assay.
Vaccine candidate preparation and evaluation
In total, 100 8-week-old specific pathogen-free (SPF) chickens were equally divided into five groups. The purified rPH-1 protein at 25 and 50 μg was diluted in 0.5 mL of phosphate-buffered saline (PBS) and mixed with ISA70 adjuvant (Seppic, France) at a 30:70 (v/v) ratio. Group 1 (25 μg rPH-1 protein), group 2 (50 μg rPH-1 protein), group 3 (25 μg rPH-1/ISA70), group 4 (50 μg rPH-1/ISA70), and group 5 (distilled water) were injected with a total volume of 100 μL intramuscularly. Three weeks after prime, chickens in each group were boosted with the same vaccination regimen. The chickens were kept in a controlled environment, with free access to food and water for 3 months. The animal trial was performed strictly in accordance with the ethical guideline issued by Razi Vaccine and Serum Research Institute (
To determine the immune responses, blood samples were taken from all chickens at 2 weeks interval until the end of the experiment. The humoral immune response was assessed with both commercial kit (BioChek) and rPH-1 protein-based enzyme-linked immunosorbent assay (ELISA). In this case, an amount of 0.5 μg/mL rPH-1 protein was coated onto the flat-bottomed 96-well microplate (100 μL/well) overnight at 4°C. The plate was blocked by 5% skim milk for 1 h at room temperature, and subsequently, 100 μL of the diluted individual serum (1:500 in PBS containing 0.03% Tween-20) was added to the wells and incubated for 1 h. After three times washing, the wells were incubated with horseradish peroxidase (HRP)-conjugated goat antichicken IgG for another 1 h. The wells were washed and incubated with 50 μL of 3,3′,5,5′-tetramethylbenzidine (TMB) substrate for 15–30 min in the dark. The reaction was stopped by the addition of 100 μL of 2 N H2SO4 and then OD was measured at 450 nm using an automated microplate reader. The negative samples and ILTV positive samples confirmed through detection with a commercial ELISA kit were included as duplicates in the plate. Cutoff value was defined as the mean of OD450 plus threefold standard deviation (SD) of 20 negative control sera taken from unvaccinated SPF chickens at 95% confidence interval.
The T cell proliferation response to the PH-1 vaccine candidate was determined on day 28 after primary immunization. The peripheral blood mononuclear cells (PBMCs) were seeded in a 96-well plate at 2 × 105 cells/well in 5 mL of antibiotic-free DMEM supplemented with 10% fetal bovine serum. Cell cultures were stimulated either with rPH-1 or with phytohemagglutinin (PHA; 10 μg/mL). Cells were for 12 h at 37°C and 5% CO2 then 10 μL of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) (5 μg/mL final concentration) was added to each well. After a 4-h incubation, the absorbance was read at OD540 and proliferative activity for each group was expressed as a stimulation index (SI). The SI was calculated as the mean OD540 of stimulated cells–the mean OD540 of blank divided by the mean OD540 of unstimulated cells.
Statistical analysis
All experiments were performed three times and the data are presented as mean ± SD. Comparison among groups was done by using one-way analysis of variance (ANOVA) followed by t test. A p-value of <0.01 was considered to be statistically significant.
Results
The ILTV isolate was efficiently grown in chicken lung cells and produced rounded cells that fused with adjacent cells to form syncytia. The infected cells were detached at 72 h postinfection compared with uninfected cells (Fig. 1).
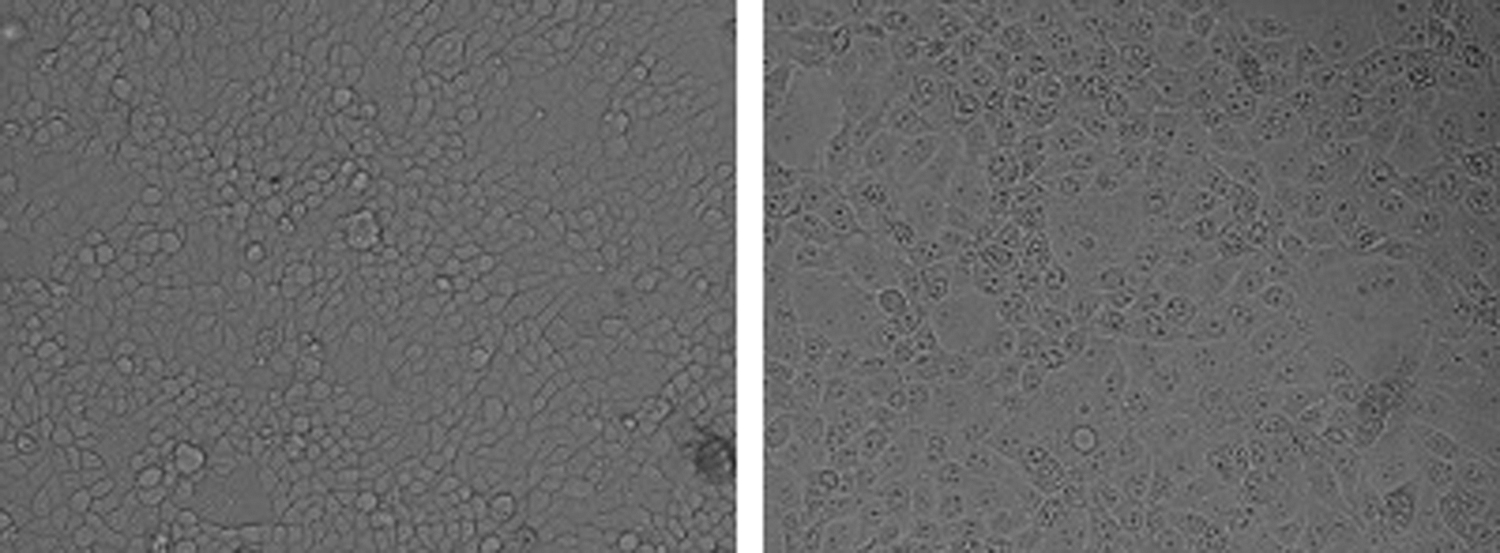
Isolation of ILTV on chicken lung cells. Syncytia forms at 72 h postinfection (right) compared with uninfected cells (left). ILTV, infectious laryngotracheitis virus.
The optimized length of the gB PH-1 domain sequence is 624 nucleotides. The GC content was calculated as 49.77% that lies in the optimum range (30–70%), and Codon Adaptation Index (CAI) was 1.00, which indicates high expression of gene in E. coli. The population average for the experimental data set (PopAvrSol) is 0.45, and, therefore, any scaled solubility value greater than the score is predicted to have a higher solubility. The scaled solubility value for PH-1 was estimated as 0.52 (higher than 0.45), which denotes the protein can express efficiently in E. coli host. The optimized codon was inserted between the BamHI and XhoI restriction sites at MCS of the pET28a (+) vector (Fig. 2).
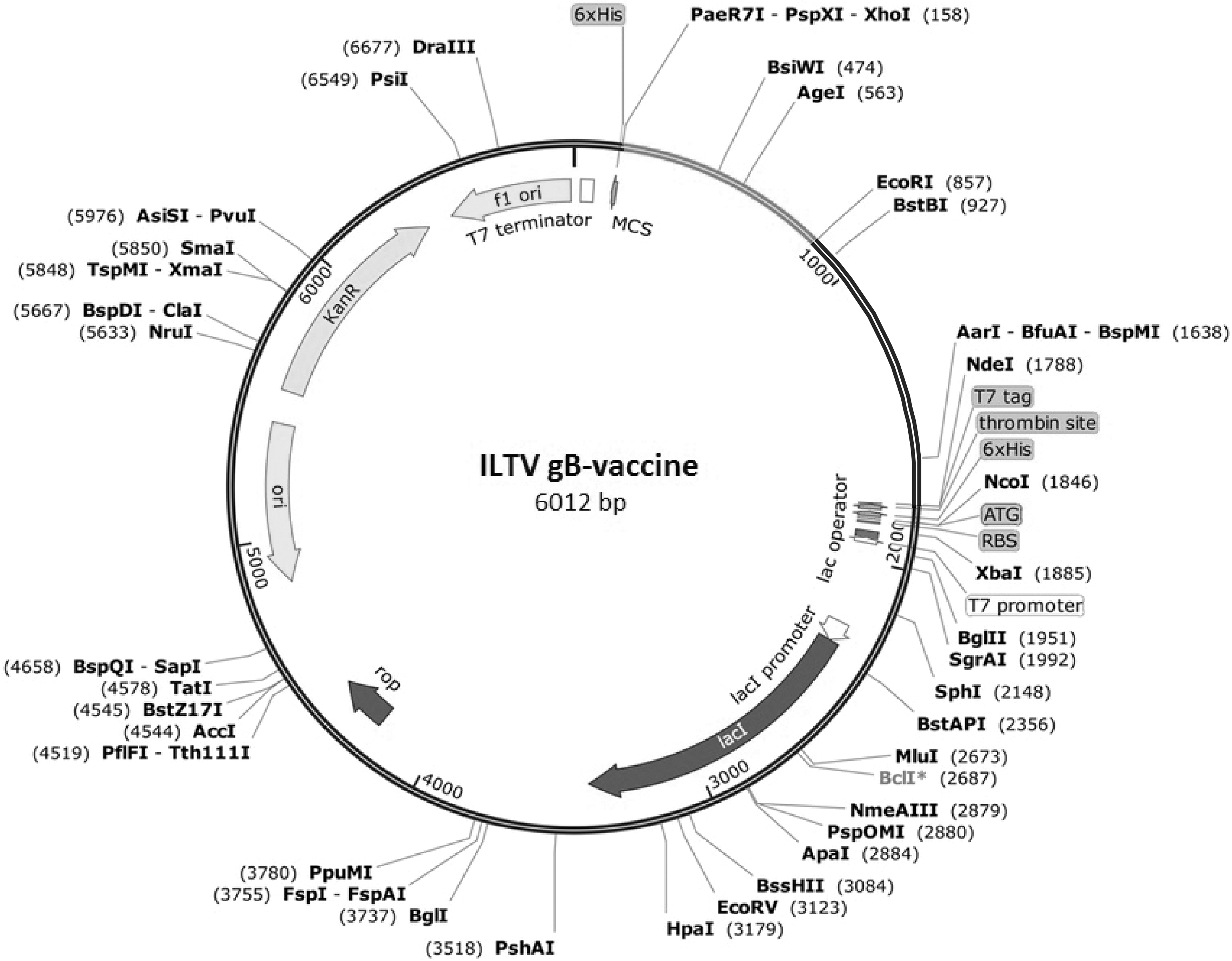
In silico cloning of the ILTV PH-1 gB into pET28a (+) vector. Light gray line represents the gene fragment insert between the XhoI-BamHI restriction sites of the vector.
The gene encoding PH-1 domain was amplified and cloned into the pET-28a (+) vector using the heat-shock procedure. The integrity of the recombinant vector was confirmed by double digestion using the enzymes digestion and sequencing. High-level expression of gB PH-1 protein was induced in the presence of IPTG after 4 h incubation. The rPH-1 yield was estimated ∼4.52 mg/mL of bacterial culture. The SDS-PAGE electrophoresis result showed an identifiable band at ∼24 kDa, which was similar to that predicted by ProtParam in our previous in silico study.
Induction of humoral immune responses to rPH-1 was assayed using ELISA. The cutoff level was 0.3, as obtained by calculating the mean OD450 of 0.162 and SD of 0.461 for the negative sera. Serum OD values detected by the rPH-1 ELISA were significantly higher in pullets vaccinated with the rPH-1/ISA70 compared with the rPH-1 alone (p < 0.01). However, the mean detected ODs of the two concentrations of rPH-1 were not different, meaning that increasing the amount of antigen from 25 to 50 μg has no effect on the induction of humoral response. The OD level increased at the second time point of blood sampling for all of the treatment groups and decreased at the end of the period in pullets that received rPH-1 only. The level was increased for the rPH-1/ISA70 treatment groups and remained constant at end of the experiment. No ILTV antibody titer was detected in control group (Fig. 3). Poor correlations were found between the results of rPH-1 ELISA and the commercial kit. In the determination of antibodies against rPH-1 by commercial ELISA, undetectable or low antibody levels were observed in the treatment groups, especially the 25 μg rPH-1 alone group. However, a significant rise in antibody level was observed in both rPH-1/ISA70 groups compared with the rPH-1 alone groups (p < 0.01). No ILTV antibody titer was detected in the control group (Fig. 4).

Detection of specific ILTV antibody using rPH-1-based ELISA. Values are expressed as mean counts ± SE. *Significant difference (p < 0.01) compared with dH2O; **significant difference compared with rPH-1 alone. ELISA, enzyme-linked immunosorbent assay; rPH-1, recombinant PH-1; SE, standard error.
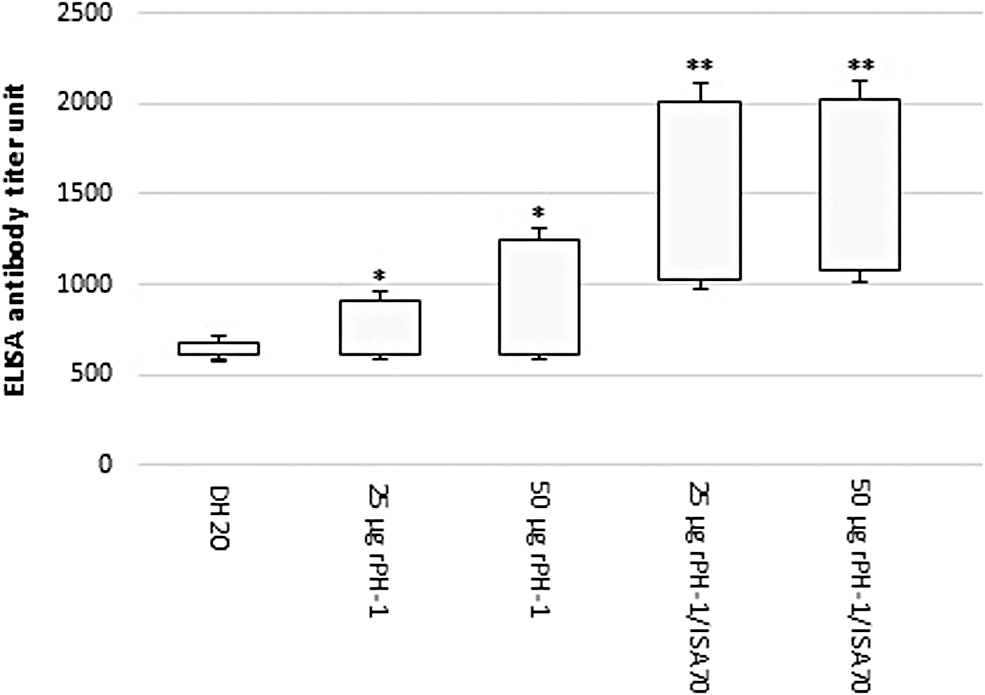
Mean of specific ILTV antibody response to rPH-1 protein using commercial ELISA kit. Values are expressed as mean counts ± SE. *Significant difference (p < 0.01) compared with DH2O; **significant difference compared with rPH-1 alone. A titer >1071 was considered as positive according to the kit instruction.
To determine the T cell proliferative response, PBMCs were prepared 1 week after the booster injection and assayed. A significant response of T cells was observed in all groups of vaccinated chickens as compared with the negative control group (p < 0.01) (Fig. 5). In stimulation with rPH-1, cells isolated from the chickens injected with both concentrations of rPH-1/ISA70 showed high SI. The proliferations with both concentrations of rPH-1 alone were relatively lower than those with the rPH-1/ISA70, which was not significant.

Peripheral blood T lymphocyte proliferation response to ILTV rPH-1 protein. The SI values are expressed as mean counts ± SE. *Significant differences from controls were signed with * (p < 0.01). SI, stimulation index.
Discussion
Since the first report of ILTV, vaccination has been considered an essential tool against this disease in endemic areas. The use of live attenuated vaccines is questionable due to the ability of the virus vaccine to revert to the virulent. Recombinant vaccines are an attractive alternative to traditional vaccines to overcome this disadvantage, however, they need adjuvants to trigger the desired immune responses (5,6,17). Based on the results of in silico study (12), we used the immunogenic part of gB protein for construction of a subunit vaccine candidate to enhance immune responses against ILTV. The rPH-1 protein adjuvanted with ISA70 could provide specific ILTV immune responses in SPF chickens.
gB is the most highly conserved herpesvirus envelope protein that mediate the fusion of viral and cellular membranes. This interaction leading to virus entry into the host cell. The protein binds to the host cellular lipid rafts through TLR2, activates nuclear factor kappa B (NF-κB) through the MyD88/TRAF6-dependent signal transduction pathway, and contributes to cytokine expression during the early stage of virus infection (4). The ILTV gB initially synthesizes a 110 kDa monomeric precursor protein in the endoplasmic reticulum and rapidly converts to homodimers. Maturation of the protein is done by proteolytical cleavage of the dimers in the Golgi apparatus (23). In this activated form, gB acts as an immunogen and is targeted for the induction of immune responses. Researches on immune potentials of ILTV proteins suggested that gB has a high antigenic feature and can elicit neutralizing antibodies and induce protective immune responses (2,7,29,30). To reduce the adverse effects of a full-length sequence antigen in vaccine formulations, one domain of gB that contributes to induce immunity was used. The ectodomain of gB is structurally composed of five functional domains and PH domains are involved in intracellular signaling, leading to T cell activation and cytokine expression (11).
In immunization with subunit vaccines, efficient immune response is highly dependent on adjuvant that enhanced the immunogenicity of a recombinant protein. Water-in-oil emulsions triggered an inflammatory response at the injection site and form a depot to protect the antigen from rapid release and degradation (1). Therefore, they may guarantee a long-term protective immune response. Humoral immune response was enhanced and prolonged by coinjection of rPH-1 and ISA70 compared with the injection of rPH-1 alone. This may explain depot formation by the adjuvant at the injection site to facilitate the slow release of antigen. The delivery platform promotes immune responses to the protein by prolonged exposure to the immune system and enhancing antigen uptake. Examination of the sera with the rPH-1-based ELISA showed that the recombinant protein could induce antigenic properties that significantly improve in the presence of adjuvant. The antibody levels were low and in some cases undetectable in vaccination with rPH-1 alone, although in vaccination with rPH-1/ISA70, they rose and thereafter remained stable. Estimation of low antibody levels in vaccinated chickens may be due to the use of commercial ELISA kits that utilize the whole virus antigen. Despite the sensitivity and specificity, it seems that this ELISA is inefficient for detecting ILTV antibodies in sera from vaccinated chickens with recombinant vaccines (16). The use of the specific glycoprotein-based ELISA may improve the evaluation of humoral responses after vaccination with the same protein. Unfortunately, the ELISA is not being commercially available.
Immune responses to ILTV are differentiated into immediate innate inflammatory responses and prolonged adaptive immune responses. Innate immune responses are induced by the production of inflammatory cytokines and infiltration of leukocytes into infected tissues at the early step of viral infection (8). It has been suggested that innate and cell-mediated, rather than humoral immune, responses play important roles in protecting chickens against ILTV infection (16,28). Therefore, we evaluated the proliferation response of lymphocytes after vaccination with rPH-1 in the presence or absence of adjuvant. Based on SI, the response in all groups immunized with rPH-1 was enhanced, indicating that the protein is immunogenic. Except for the quality of an antigen, recombinant proteins require a combination with adjuvants to enhance and direct the adaptive immune response against ILTV (5,6,17,27). ISA70 is widely used in poultry commercial inactivated vaccine formulations. But the adjuvant is not able to preferential stimulation of Th1-type cell-mediated immunity. We showed that there were no significant differences between the proliferation rate in the rPH-1/ISA70 groups compared with that of recombinant antigen alone. This may explain the potential of rPH-1 in induction of cellular immune responses.
For the efficient induction of immune responses, the determination of an optimum antigen dose and its quality are critical factors. In this study, the potential role of two concentrations of rPH-1 in the efficacy of the ILTV vaccine was evaluated. The same immune responses were induced in chicks that received the rPH-1 either at 25 μg or at 50 μg compared with control chicks. The low concentration of rPH-1 has the potential to induce immunity that is not toxic for the embryo; however, to induce an immune response in chickens, it needs adjuvant. In general, subunit antigens are insufficiently immunogenic and require suitable adjuvant for augmenting their immunogenicity. Success in nanotechnology has provided alternative strategies for the efficient delivery of the unstable subunit antigens. Encapsulation of the vaccine antigen with adjuvant in nanoparticles may result in increased stability and cellular uptake of antigen (24). Although this vaccination is an attractive delivery system, more experiments need to be done for confirmation of the final formulation of the vaccine candidate. The basic understanding of factors influencing the nanoparticle delivery systems, as well as the quality of the antigen–adjuvant cooperation, provides the design of a more effective vaccine formula.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by Razi Vaccine and Serum Research Institute under grant no. 2-18-18-94103.