
Abstract
Failure of current therapies to cure chronic hepatitis B has led to renewed interest in therapies that stimulate the host immune system. APOBEC3 (A3) family enzymes have been shown to induce mutations in hepatitis B virus (HBV) covalently closed circular DNA (cccDNA) leading to inhibition of HBV transcription and replication. Pattern recognition receptor (PRR) agonists have been reported to suppress HBV, but it is unclear whether these agonists induce A3 gene expression in hepatocytes. We, therefore, evaluated whether PRR signaling activates the expression of A3 genes and other innate immunity genes and restricts HBV infection. HepG2-sodium taurocholate cotransporting polypeptide (NTCP) cells were infected with HBV and treated with various PRR agonists. The level of HBV infection was subsequently assessed by measurement of HBV biomarkers, including HBV DNA, cccDNA, HBs, and HBe antigens in infected hepatocytes. Among all tested PRR ligands, only Poly(I:C)-HMW/LyoVec and Poly(I:C)-HMW significantly inhibited hepatitis B surface antigen (HBsAg), hepatitis B e antigen (HBeAg), HBV DNA, and cccDNA, whereas R848 and lipopolysaccharide (LPS) only showed significant inhibition on HBsAg and HBeAg, but not virus DNA. CpG and Pam3CSK4, on the other hand, had no significant inhibitory effect on any of the HBV infection parameters. Moreover, Poly(I:C)-HMW/LyoVec and Poly(I:C)-HMW were the only ligands that significantly increased IL-8 secretion. Interestingly, HBV infection reduced IL-8 secretion induced by Poly(I:C)-HMW and to a lesser extent Poly(I:C)-HMW/LyoVec. Poly(I:C)-HMW/LyoVec had a significant effect on increasing the expression level of A3F, A3G, A3H, TLR3, RIG-1, and MDA5 genes. Our data suggest that PRR agonists may control HBV infection through different mechanisms. The RIG-1 and MDA5 agonist, Poly(I:C)-HMW/LyoVec, seems to downregulate HBV infection through induction of A3 genes.
Introduction
Despite the availability of an effective vaccine, hepatitis B virus (HBV) is still a worldwide health problem with >250 million chronically infected patients. The virus is a leading cause of liver diseases, such as cirrhosis and hepatocellular carcinoma (29).
HBV is a small enveloped DNA virus; after infection, the capsid is transported to the nucleus, where relaxed circular DNA (rcDNA) is converted into a covalently closed circular DNA (cccDNA). cccDNA serves as a template for HBV RNA synthesis and its eradication is the main target for the treatment of chronic hepatitis B infection (CHB) (4,6). HBV leads to acute or chronic infection, and the majority of patients with acute hepatitis can eliminate the virus with no requirement for the treatment. However, chronic HBV infection is the result of a dynamic interaction between HBV, host hepatocytes, and immune system. CHB patients in comparison with patients with acute infection have either impaired or distorted or diminished immunity (11); therefore, immunotherapy could be an alternative approach to control or suppress viral replication by potentiating antiviral immunity in CHB patients. Pegylated interferon (IFN)-α and oral nucleotide/nucleoside analogs are current treatment modalities for chronic patients, but unsuccessful results like not being able to eradicate cccDNA by these treatments has led to renew interest in immunotherapy to improve the host immune system (5).
Activation of innate immunity is the first-line response against pathogens. Pattern recognition receptors (PRRs), as crucial part of innate immunity, sense pathogens and mediate initiation of adaptive immune response and release of inflammatory cytokines (18). PRRs are classified as toll-like receptors (TLR), retinoic acid-inducible gene-1 (RIG-1)-like receptors (RLR), NOD-like receptors (NLR), and C-type lectin receptors (CLR). PRRs detect pathogen-associated molecular patterns (PAMP) derived from different pathogens, such as viruses and bacteria and activate downstream signaling pathways leading to specific gene expression and secretion of anti-viral cytokines and other antimicrobial peptides.
Antiviral cytokines, such as type I and type III IFN have been shown to induce anti-HBV effect in hepatocytes (2,8). Many viruses can activate PRR pathways; however, it has been shown that HBV is a “stealth” virus and innate immune system cannot detect HBV in early stage of infection (37). Moreover, HBV proteins like polymerase, HBx, and hepatitis B surface antigen (HBsAg) inhibit PRR signaling pathways (8,19,35). Therefore, activation of innate immunity and using PRR agonists could be a promising candidate for immunotherapy to improve the immune response in CHB patients.
To replicate efficiently, viruses must overcome the innate immunity. Human apolipoprotein B mRNA-editing catalytic polypeptide (APOBEC) family comprises cytosine deaminase enzymes, which are a powerful intracellular barrier and play an important role in innate immune response against viruses through deamination of their DNA and/or RNA sequence (12). Recent studies have shown that APOBEC3 (A3) subfamily could inhibit HBV infection. It has been shown that one of the mechanisms by which various cytokines, such as IFNλ (28), IFNγ, and tumor necrosis factor (TNF-α) (39) can inhibit HBV infection is mediated through increasing A3 gene expression.
Recently developed HepG2-NTCP cell line can support a complete HBV life cycle, including the early steps of infection. The details on the innate immune responses in this in vitro model are not fully characterized. In other words, the innate immune responses in hepatocytes after HBV infection and also after immunotherapy with immunomodulators need further investigation. To provide a better understanding of the potential role of TLR agonists in induction of an efficient and long-lasting antiviral effect in established HBV infection, the anti-HBV effect and functional activity of a wide spectrum of TLR agonists in HBV-infected HepG2-NTCP cells were evaluated. Since there is no report regarding the effect of PRR agonists on induction of A3 gene expression in hepatocytes, in the present study, we addressed this issue and investigated the inhibitory effect of PRR agonists on innate immunity genes and HBV infection in HepG2-NTCP cells.
Materials and Methods
Cell culture
HepG2-NTCP cells (generously provided by Professor Ulrike Protzer from Institute of Virology, Technische Universität München/Helmholtz Zentrum, München) were cultured in Dulbecco's modified Eagle's medium (DMEM-low glucose) (Gibco, United Kingdom) supplemented with 10% fetal bovine serum (FBS) (Gibco, USA), 100 U/mL penicillin and 100 μg/mL streptomycin (Sigma), nonessential amino acids (Gibco, United Kingdom), and 30 μg/mL Blasticidin (Gibco, USA) at 37°C in 5% CO2 humidified incubator.
HepAD38 cells (kindly provided by Professor Christoph Seeger from the Institute for Cancer Research, Fox Chase Cancer Center, Philadelphia) were cultured in DMEM/F12 medium (Gibco, United Kingdom) supplemented with 10% FBS, 100 U/mL penicillin and 100 μg/mL streptomycin, and nonessential amino acids.
Preparation of HBV inoculum
HepAD38 cells, which stably produce ayw subtype HBV (genotype D) (20), were cultured on collagen-coated flasks in the absence of tetracycline to induce HBV replication. Virus particles were precipitated and concentrated 120-fold in the presence of 8% PEG 8000 (Sigma). The concentrated virus stock was aliquoted and stored at −70°C, and the HBV viral load was determined with a TaqMan-based quantitative real-time PCR (qPCR) method (Gene Proof, Czech Republic).
HBV infection and cell stimulation
HepG2-NTCP cells were cultured in 24-well collagen-coated plate with density of 5 × 104 cells/well for 3 days. To differentiate the HepG2-NTCP cells, which is necessary for successful HBV infection, the cells were cultured in DMEM (low glucose) differentiation medium containing 1.74% DMSO, 1% FBS, 100 U/mL penicillin and 100 μg/mL streptomycin, and nonessential amino acids. After 2 days of differentiation, cells were infected with the concentrated HBV (obtained from supernatant of HepAD38 cells) at a multiplicity of infection (MOI) 400 genome equivalent/mL (GEq/cell). HBV infection of differentiated HepG2-NTCP cells was performed in DMEM differentiation medium supplemented with a final concentration of 2.5% PEG8000 for 16 h. Subsequently, the HBV inoculum was removed and cells were washed three times with PBS and fresh differentiated medium was added to each well.
At 5 and 9 days postinfection (dpi), the supernatant of cells was changed and then first and second treatments with PRR agonists were performed, respectively. The third treatment with PRR agonists was performed at 13 dpi without changing the medium. After 24 h (14 dpi), the supernatant of cells was collected and DNA and RNA of cells were extracted (Fig. 1). Establishment of HBV infection in HepG2-NTCP cells was determined by measuring cccDNA and HBV DNA by qPCR and also quantification of HBsAg and HBeAg in the supernatant of infected cells by enzyme-linked immunosorbent assay (ELISA). In all experiments, Myrcludex-B (Myr), (kindly provided by Professor Stephen Urban from Department of Infectious Diseases, Molecular Virology, University Hospital Heidelberg, Heidelberg), as inhibitor of HBV infection that inhibits the virus entry, was used (33). Seven hundred nanomolar of Myr was incubated with cells for 30 min before infection and then HBV inoculum was added to treated cells. Fresh medium lacking Myr was replaced with the culture medium after 16 h of incubation.

Schematic of the experimental design. HepG2-NTCP cells were cultured in a 24-well plate for 3 days, and then cells were maintained in differentiation medium. After 2 days of differentiation, cells were infected with the concentrated HBV at MOI 400 GEq/cell. Following removal of the HBV inoculum, cells were washed three times with PBS and fresh differentiation medium was added to each well. At 5 and 9 dpi, the supernatant of cells was changed and then first and second treatments with PRR agonists were performed, respectively. The third treatment with PRR agonists was performed at 13 dpi without changing the medium. At 14 dpi, the supernatant of cells was collected and DNA and RNA of cells were extracted. Establishment of HBV infection was determined by measuring cccDNA and HBV DNA by qPCR and also quantification of HBsAg and HBeAg in the supernatant of infected cells by ELISA. cccDNA, covalently closed circular DNA; dpi, days postinfection; ELISA, enzyme-linked immunosorbent assay; HBV, hepatitis B virus; HBeAg, hepatitis B e antigen; HBsAg, hepatitis B surface antigen; HepG2-NTCP, HepG2-sodium taurocholate cotransporting polypeptide; MOI, multiplicity of infection; PRR, pattern recognition receptor; qPCR, quantitative real-time PCR.
TLR and RLR agonists
All PRR agonists were purchased from InvivoGen, except lipopolysaccharide (LPS), which was purchased from Sigma. TLR1/2, TLR3, TLR4, TLR7/8, TLR9, and RIG-1/MDA5 were, respectively, stimulated/engaged with Pam3CSK4 (500 ng/mL), Poly(I:C)-HMW (5 μg/mL), LPS (500 ng/mL), Resiquimod or R848 (5 μg/mL), CPG ODN 2006 (5 μM or 38.5 μg/mL), and Poly(I:C)-HMW/LyoVec (100 ng/mL).
Detection of interleukin-8 and HBV infection seromarkers
The supernatant of infected and/or stimulated cells were harvested at 14 dpi and examined for the presence of IL-8, HBsAg, and HBeAg by ELISA. Measurement of IL-8 was carried out by the Sanquin IL-8 ELISA kit (Sanquin, Amsterdam, Netherlands) according to the manufacturer's instructions. HBeAg was measured by the commercial ELISA Kit (Dia Pro, Italy) according to the manufacturer's protocol.
For HBsAg detection by ELISA, 10 μg/mL of two affinity-purified anti-HBs monoclonal antibodies (4G4 and 6E3 mAbs), which have been previously generated and characterized in our laboratory (7,9,10), were coated on ELISA plate. After overnight incubation and blocking with 1% bovine serum albumin, supernatant of infected cells was added and incubated for 1h at 37°C. After washing with PBS containing 0.05% Tween 20 (Sigma), biotin-conjugated anti-HBs mAb (Enzygnost HBsAg 6.0; SIEMENS, Germany) was added and incubated for 1 h at 37°C. Following washing, streptavidin/HRP conjugate (Enzygnost HBsAg 6.0) was added and incubated for 1 h at 37°C. After washing, 3,3′,5,5′-tetramethylbenzidine (TMB) substrate was added and the reaction stopped with H2SO4 and optical density measured at 450 nm by an ELISA reader (BioTek PowerWave XS).
Nucleic acid extraction and qPCR/quantitative reverse transcription PCR
At 14 dpi, infected and/or stimulated cells were collected for RNA and DNA extraction. Total DNA was extracted with phenol/chloroform/isoamyl alcohol based on the method described by Sun et al. (30), and used for quantification of cccDNA and HBV DNA. RNA was isolated with the NucleoSpin RNA II Kit (Macherey-Nagel, Germany) and cDNA synthesis was performed using M-MuLV reverse transcriptase (Thermo Fisher), and the cDNA was used for quantification of TLRs, RLRs, and APOBECs gene expression. qPCR was performed using the Hot Firepol Evagreen qPCR Mix Plus (Solis Biodyne, Estonia) and StepOnePlus Real-Time PCR System (Applied BioSystems). The genes were normalized to housekeeping genes PRNP (PRioN Protein) and RPLP0 (ribosomal protein lateral stalk subunit P0) as internal controls for DNA and cDNA quantification, respectively, and data were analyzed with 2−ΔΔCT method.
During cccDNA-selective PCR, due to the presence of HBV DNA molecules, a false-positive cccDNA detection may happen. To avoid false-positive detection of cccDNA, extracted DNA was treated with T5 exonuclease (New England Biolabs). Briefly, 8.5 μL of extracted intracellular total DNA was incubated with 5 U of T5 exonuclease for 2 h at 37°C and was subsequently inactivated by incubation at 95°C for 2 min. After T5 exonuclease treatment, 30 μL of distilled water was added to each sample to achieve fourfold dilution and samples were quantified by qPCR. HBV DNA, cccDNA, and housekeeping gene were evaluated in untreated samples simultaneously. The primer sequences used in this study are listed in Table 1.
The Primers Sequences Employed in This Study
Statistical analysis
Statistical analysis was performed using GraphPad Prism 6 and Student's t-test. For all tests, p-values <0.05 were considered as statistically significant. [p < 0.05 (*), p < 0.01 (**), p < 0.001 (***), and p < 0.0001 (****)].
Results
Expression pattern of PRR and APOBEC genes in HepG2-NTCP cells
To determine the expression pattern of innate immunity genes in HepG2-NTCP cells, differentiated cells were lysed and their RNA were extracted and cDNA was synthesized, as described in the Materials and Methods section. PRR and APOBEC mRNA expression levels were quantified in differentiated HepG2-NTCP cells with quantitative reverse transcription PCR (RT-qPCR). PRRs that we evaluated in this study were TLR1, 2, 3, 4, 6, 7, 9, 10, RIG-1, and MDA5. Our results showed that RIG-1 and MDA5 were highly expressed, and TLR1, 3, 6, and 9 were moderately expressed, whereas TLR2, 4, 7, and 10 were expressed at low levels in HepG2-NTCP cells (Fig. 2A). Among the APOBEC3 gene family A3C, A3F, and to a lesser extent A3B genes were highly expressed. Whereas A3A, A3DE, A3G, and A3H genes were expressed at a low level or almost undetectable (Fig. 2B).

mRNA expression level of TLR, RLR, and APOBEC in differentiated uninfected HepG2-NTCP cells. Differentiated HepG2-NTCP cells were lysed and their RNA was isolated and cDNA synthesis was performed as described in the Materials and Methods section. mRNA expression level of TLR, RLR
The antiviral effect of TLR/RLR ligands on established HBV infection in HepG2-NTCP cells
To evaluate the antiviral effects of TLR/RLR ligands on HBV infection, HepG2-NTCP cells were infected with HBV at MOI 400 GEq/cell and treated with Pam3CSK4, Poly(I:C), LPS, R848, CpG ODN 2006, and Poly(I:C)/LyoVec ligands. Based on HBeAg and HBsAg levels, all TLR/RLR ligands, except Pam3CSK4 and CpG were found to significantly inhibit HBV infection in HepG2-NTCP cells. However, Poly(I:C)/LyoVec and Poly(I:C) were the only ligands that showed inhibitory effect on HBV DNA and cccDNA. The most antiviral effect was observed by Poly(I:C)/LyoVec, as evidenced by significant reduction of HBsAg (64%), HBeAg (58%), cccDNA (82%), and HBV DNA (60%). In addition to Poly(I:C)/LyoVec, the other ligand that could significantly reduce HBV DNA was Poly(I:C), which led to 58% and 43% reduction of HBV DNA and cccDNA, respectively. Besides suppressing the HBV DNA and cccDNA, Poly(I:C) significantly suppressed HBsAg (18%) and HBeAg (28%) (Fig. 3A–D)
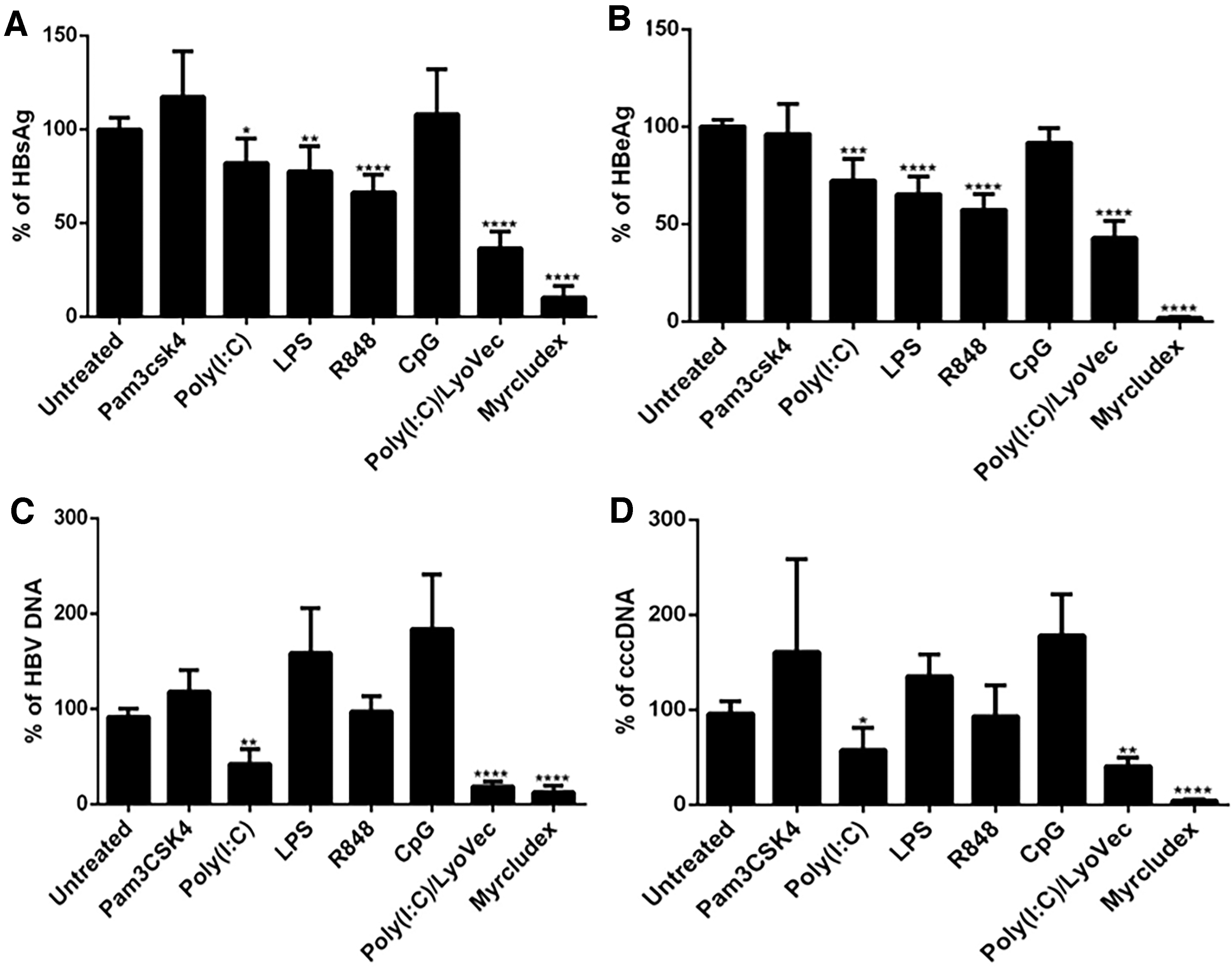
Antiviral effect of TLR/RLR ligands on established HBV infection in HepG2-NTCP cells. HepG2-NTCP cells were infected by HBV at MOI 400 GEq/cell, then treated three times with the indicated ligands (see Materials and Methods section for concentrations) at 5, 9, and 13 dpi. Twenty-four hours after the last treatment (14 dpi), cell supernatants were collected and HBsAg
Although R848 significantly inhibited HBsAg (34%) and HBeAg (43%) production in HBV-infected cells (Fig. 3A, B), this ligand did not show significant inhibitory effect on HBV DNA or cccDNA (Fig. 3C, D).
Based on HBsAg and HBeAg levels, a modest antiviral effect was observed following LPS treatment leading to a 23% and 35% reduction in HBsAg and HBeAg levels, respectively (Fig. 3A, B). Myr, as an inhibitor of HBV attachment and entry, efficiently inhibited HBV infection, as evidenced by a significant reduction in all HBV infection biomarkers (Fig. 3A–D).
Since Poly(I:C) and Poly(I:C)/LyoVec were the only ligands with inhibitory effect on all four HBV infection markers, including HBsAg, HBeAg, rcDNA, and cccDNA, we continued the rest of our study with only these two ligands.
The effect of Poly(I:C) and Poly(I:C)/LyoVec on secretion of IL-8
IL-8 is a chemokine, which is secreted after activation of TLR signaling pathways (16). We assessed the induction of IL-8 secretion after TLR ligand stimulation to make sure that the TLR ligands have a stimulatory effect on HepG2-NTCP cells and also to evaluate the general functionality of TLR3 and RIG-1/MDA5 in our system. In addition, production of IL-8 was compared between HBV-infected and also HBV-uninfected HepG2-NTCP cells to evaluate the effect of HBV infection on the production of this cytokine. The cell culture supernatant of HBV-infected and Poly(I:C)- and/or Poly(I:C)/LyoVec-stimulated HepG2-NTCP cells at 14 dpi was tested for production of IL-8; supernatant of agonist-unstimulated and HBV-uninfected and/or HBV-infected cells served as control (Mock) (Fig. 4). Our results showed that IL-8 production was significantly increased after Poly(I:C) and Poly(I:C)/LyoVec treatment, confirming the functionality of TLR3 and RIG-1/MDA5 pathways in HepG2-NTCP cells. Although HBV infection did not induce the production of IL-8 (Mock, HBV+), viral infection was able to significantly reduce the secretion of this cytokine after Poly(I:C) stimulation. In addition, HBV infection also reduced IL-8 secretion after Poly(I:C)/LyoVec stimulation, although this effect was not statistically significant (Fig. 4).
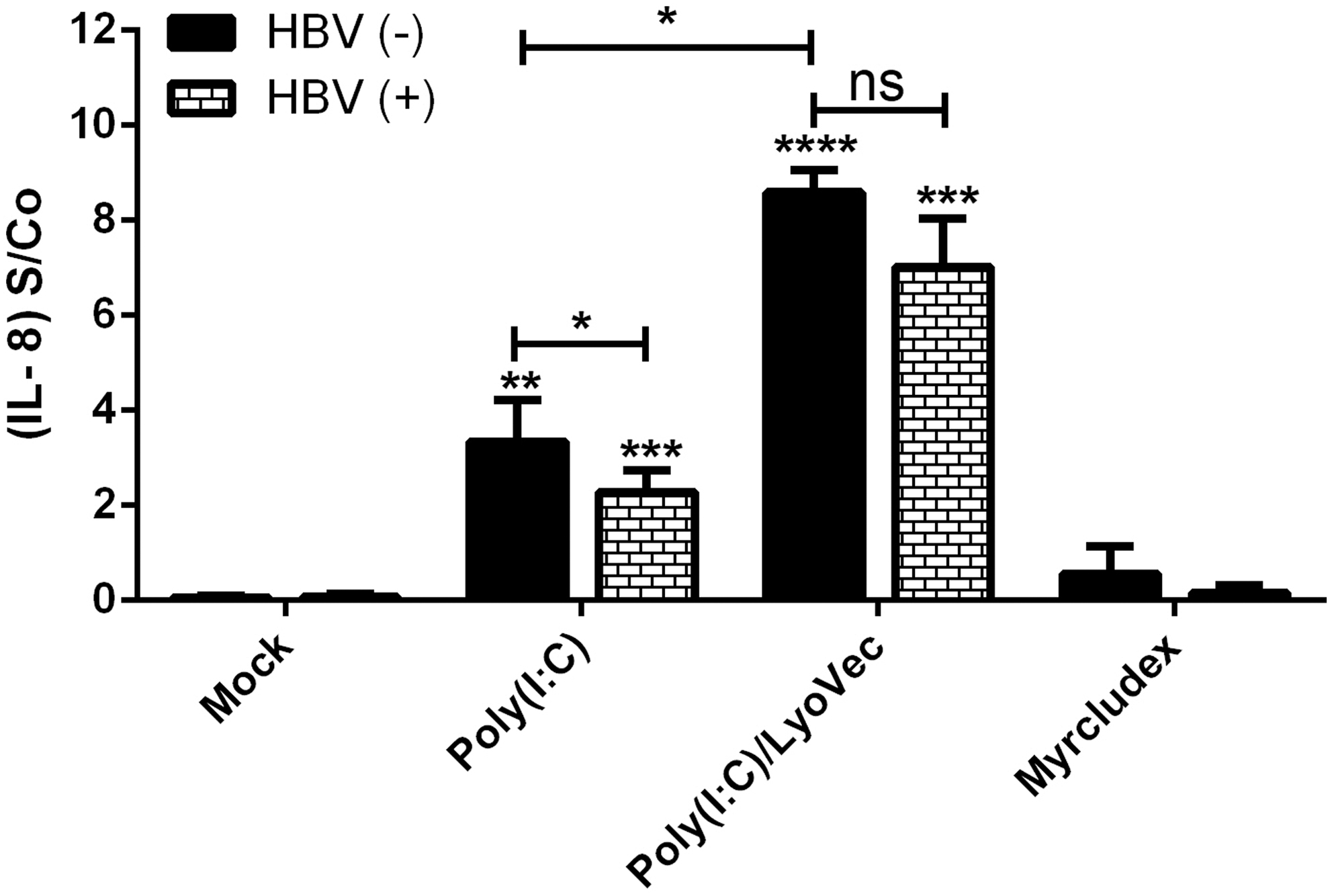
Secretion of IL-8 after HBV infection and stimulation with TLR/RLR ligands. IL-8 was measured in culture supernatants of TLR/RLRs-stimulated HepG2-NTCP cells in the presence or absence of HBV infection. HBV (+) stands for HBV-infected HepG2-NTCP cells that have been treated with TLR/RLR ligands at 5, 9, and 13 dpi, and their supernatants were collected at 14 dpi for measuring IL-8 level by ELISA. HBV (−) stands for TLR/RLRs ligand-treated HepG2-NTCP cells with no HBV infection. Mock stands for HepG2-NTCP cells [HBV (+) and/or HBV (−)] that were not stimulated with TLR/RLR ligands. Results are given as mean ± SD (n = 3) and differences were considered statistically significant compared with the Mock when p-values were p < 0.05 (*), p < 0.01 (**), p < 0.001 (***), and p < 0.0001 (****). Asterisks directly above individual bars indicate statistical difference in comparison with Mock, whereas asterisks between two bars indicate statistical difference between HBV (+) and HBV (−) groups.
The influence of Poly(I:C) and Poly(I:C)/LyoVec stimulation on the pattern of PRR and APOBEC gene expression in HepG2-NTCP cells
The expression levels of TLR3, RIG-1/MDA5, and APOBEC3 genes were determined in HepG2-NTCP-stimulated cells before and after HBV infection. Our results showed that Poly(I:C)/LyoVec significantly increased the expression level of RIG-1, MDA5, TLR3, A3F, A3G, and A3H (Fig. 5A–C, G–I). An increased expression of TLR3 after stimulation with Poly(I:C)/LyoVec indicates that direct addition of Poly(I:C) to the culture of cells is not the only route that Poly(I:C) can engage TLR3 in hepatocytes. In fact, transfection of Poly(I:C) (using LyoVec) had greater effect on TLR3 expression than direct addition of it to the culture of HepG2-NTCP cells. Therefore, we observed that Poly(I:C)/LyoVec not only induced its intracytoplasmic receptors, which are RIG-1 and MDA5, but also upregulated the expression of TLR3 in HepG2-NTCP cells (Fig. 5A–C). In addition, the expression level of MDA5 and TLR3 was significantly increased after Poly (I:C) stimulation (Fig. 5B, C).
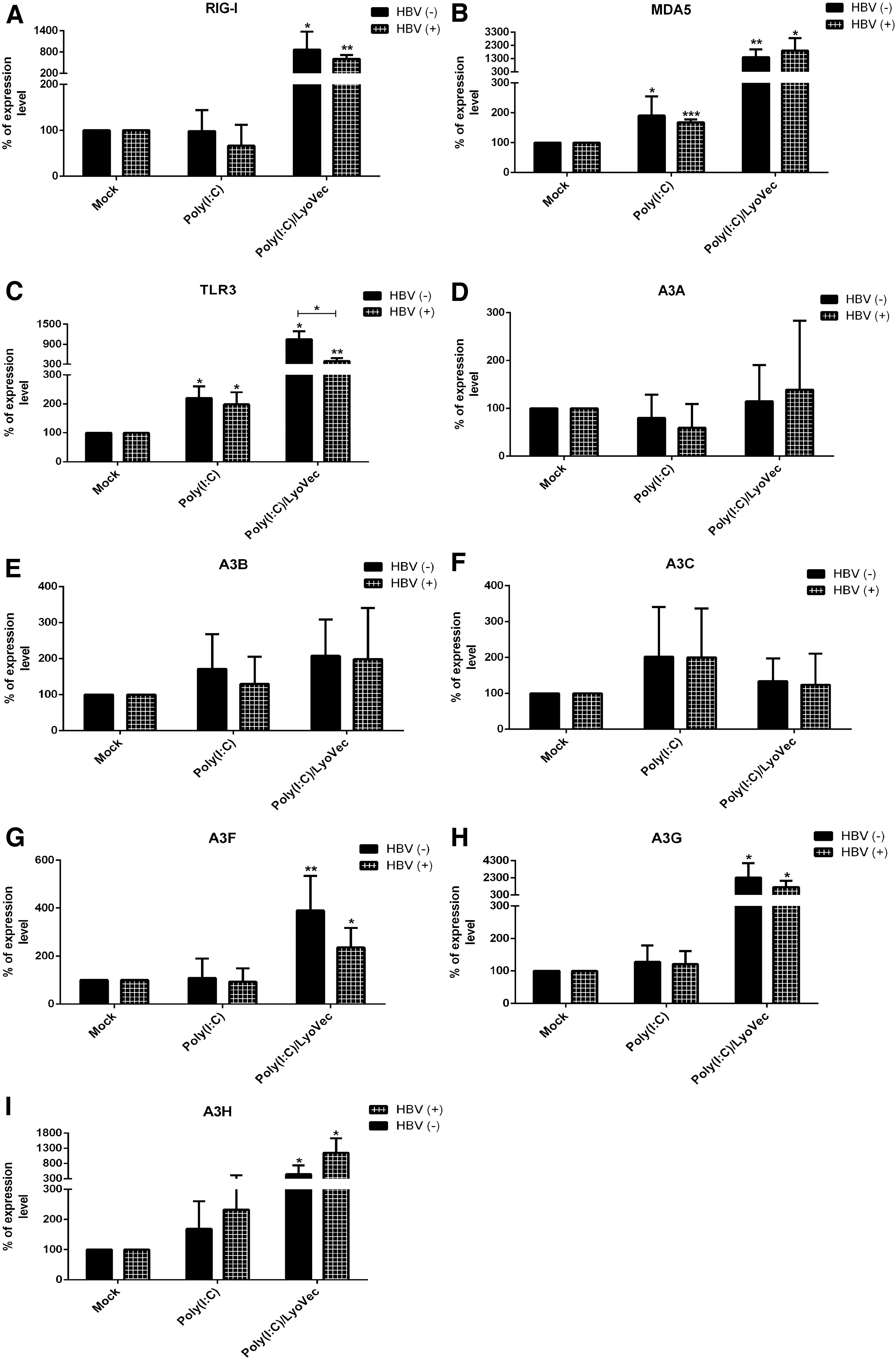
Stimulatory effect of PRR ligands on the expression level of TLR, RLR, and APOBEC3 family genes
Interestingly, HBV infection reversed the effect of Poly(I:C)/LyoVec and significantly reduced the stimulatory effect of this ligand on expression of TLR3 (Fig. 5C). In addition, HBV infection reduced the expression of A3G and A3F genes after Poly(I:C)/LyoVec treatment, but this reduction was not significant (Fig. 5G, H).
Discussion
Both innate and adaptive immunity are involved in host defense against HBV infection. Involvement of HBV-specific T cell and B cell responses in controlling HBV infection is well documented (11). However, the importance of innate immunity in eliminating HBV infection has been explored in recent years and need to be further investigated. TLRs are mediators that can link innate and adaptive immunity, and stimulation of TLRs by PAMPs or corresponding ligands leads to activation of antiviral mechanisms, including intracellular antiviral pathways, production of IFNs, and also secretion of proinflammatory cytokines (38). Experimental results from animal models of HBV infection and also in vitro models have shown that stimulation of TLRs by cognate agonists can lead to an antiviral response, resulting in the suppression of HBV replication. For instance, GS-9620, a TLR-7 agonist, has a strong antiviral effect in woodchuck and chimpanzee models. In addition, its safety and efficacy has been investigated in a phase II clinical trial in human (14,21,26). Results of a study have shown that TLR8 ligand ssRNA40 selectively activates innate immune cells in liver to produce antiviral cytokine IFNγ in the liver of both healthy and chronically infected HBV subjects (15). Addition of TLR agonist to the conventional HBV vaccine has also resulted in increasing the titer of anti-HBs antibody and enhancement of affinity maturation process to increase the avidity of anti-HBs antibody (25). Therefore, TLR agonists could be a promising candidate for the treatment of chronic HBV infection and further studies are necessary to determine the details of molecular mechanism of their action. Two recent studies have determined the TLR/RLR functionality of HepaRG cells and primary human hepatocytes (PHH) after stimulating with cognate ligands to eliminate HBV infection (23,24). However, there is no detailed information regarding the innate immune response after TLR/RLR ligand stimulation in infected HepG2-NTCP cells.
APOBEC molecules are cytidine deaminases that play an important role in defense against viruses, such as HIV and HBV (34). In spite of the antiviral role of APOBECs, correlation between A3G activity and generation of drug resistance mutations in the HIV genome has recently been reported, which suggests that APOBECs may simultaneously aid host and the virus: as a preexisting innate immunity element that induces mutation in the virus genome and limits the virus replication, APOBEC3G was found to boost the ability of HIV to gain resistance during drug treatment (27). Recent studies have shown that treatment of HBV-infected hepatocytes with different cytokines, such as TNF-α, IFNγ, and IL-1, efficiently inhibited HBV infection through increasing APOBEC gene expression level (36,39). However, there is no report of the effect of TLR/RLR ligands on expression of these genes in hepatocytes. Therefore, to determine the details of the underlying mechanisms by which TLR/RLR ligands inhibit HBV infection, it is necessary to determine the effect of TLR/RLR ligands on expression of A3 family genes in hepatocytes.
First, we determined the expression pattern of A3 gene family and also TLR and RLR genes in differentiated HepG2-NTCP cells. Our results showed that RIG-1 and MDA5 were highly expressed, and TLR1, 3, 6, and 9 were moderately expressed, whereas TLR2, 4, 7, and 10 expression levels were very low or almost undetectable in HepG2-NTCP cells (Fig. 2A). Among the A3 gene family, A3B, A3C, and A3F mRNA expression levels were more than others, and A3C had the highest expression level. The rest of A3s, including A3A, A3DE, A3G, and A3H, expression levels were very low or almost undetectable (Fig. 2B). There is no report of the expression pattern of A3 gene family in HepG2-NTCP cells. However, a recent study evaluated the expression pattern of TLR and RLR genes expression in HepG2 cells. Their results showed that there is a high expression level of RIG-1, MDA5, TLR2, TLR4, and TLR6, followed by moderate expression of TLR3 and TLR5, and low expression of TLR8 and TLR9, and almost undetectable expression of TLR7 and TLR10 (23).
Pam3CSK4, Poly(I:C), LPS, Resiquimod or R848, CPG ODN 2006, and Poly(I:C)/LyoVec were used in this study to, respectively, stimulate/engage TLR1/2, TLR3, TLR4, TLR7/8, TLR9, and RIG-1/MDA5 in HepG2-NTCP cells. In addition to evaluating the effect of these ligands on HBV infection in HepG2-NTCP, which there is no report so far, effect of the ligands with the highest HBV inhibitory effect on innate immunity genes, including A3 gene family, TLR, and RLR (RIG-1 and MDA5) mRNA expression level was also assessed. Our results showed that all TLR/RLR ligands, except Pam3CSK4 and CpG, were able to inhibit HBV infection based on significant reduction in HBsAg and HBeAg levels (Fig. 3A, B). However, the only ligands that were able to significantly inhibit HBV DNA and cccDNA were TLR3 and RIG-1/MDA5 ligands (Fig. 3C, D), and thus we continued the rest of our study with only Poly(I:C) and Poly(I:C)/LyoVec, which were able to inhibit all four HBV infection markers.
Next, to confirm the functionality of TLR3 and RIG-1/MDA5 in HepG2-NTCP cells, the secretion of IL-8 was assessed after stimulating them with their corresponding ligands. As expected from the pattern of TLR expression in HepG2-NTCP cells, Poly(I:C) and Poly(I:C)/LyoVec significantly induced secretion of IL-8 in HepG2-NTCP cells (Fig. 4). Interestingly, HBV infection was able to significantly suppress IL-8 secretion from Poly(I:C)-stimulated cells. The cell viability was determined by Trypan Blue dye exclusion test and there was no significant differences between the treated and untreated cells (data not shown). To the best of our knowledge, this is the first report of the inhibitory effect of HBV on production of IL-8 after TLR3 stimulation and further studies are needed to address the details of these mechanisms.
It has been previously shown that A3B, A3C, A3G, and A3F could strongly restrict HBV replication by editing the HBV genome, as evidenced by inducing G to A hypermutation, and also a nonediting mechanism (3,17,22,31,32). In addition, the inhibitory effect of some cytokines, such as IFNγ and TNF-α, on HBV infection have been shown to be through the induction of A3 gene family (39). Since there is no report on the effect of TLR ligands on induction of A3 gene family, we decided to evaluate whether the inhibitory effect of Poly(I:C) and Poly(I:C)/LyoVec on HBV infection could be through induction of A3 gene family.
Our results showed that Poly(I:C)/LyoVec significantly increased mRNA expression of A3 genes, such as A3F, A3G, and A3H (Fig. 5G–I). Therefore, the inhibitory effect of this ligand on HBV infection might be through induction of A3F, A3G, A3H, TLR3, and RIG-1/MDA5 genes or IL-8 secretion. Interestingly, HBV infection reversed the effect of Poly(I:C)/LyoVec and significantly reduced the stimulatory effect of this ligand on expression of TLR3 (Fig. 5C). Although HBV infection caused a reduced trend in the expression of A3F and A3G after Poly(I:C)/LyoVec treatment, this reduction was not significant (Fig. 5G, H).
Poly(I:C) is another ligand that could suppress HBsAg, HBeAg, HBV DNA, and cccDNA in our study (Fig. 3A–D). Significant expression level of MDA5 and TLR3 (Fig. 5B, C) and also significant secretion of IL-8 (Fig. 4) were observed after stimulation with Poly(I:C), whereas there was no evidence of increasing the expression of A3 family genes. Therefore, the inhibitory effect of this ligand on HBV infection is likely independent from stimulation of A3 family and might be through induction of other mechanisms, such as TLR3, MDA5, and IL-8 signaling. Interestingly, HBV infection was able to significantly suppress IL-8 secretion from Poly(I:C)-stimulated cells.
Our data are in accordance with experiments performed on dHepaRG cells, which showed Poly(I:C)-LMW/LyoVec and Poly(I:C)-LMW were able to suppress total intracellular HBV DNA, HBeAg, and HBsAg secretion in HBV-infected dHepaRG cells; it is noteworthy that TLR ligand treatment in that study was performed before infecting the cells with HBV (23), whereas TLR ligand stimulation in our study was performed after establishment of HBV infection. The next study conducted by the same research group on dHepaRG cells and PHH showed that there is an increase in production of IL-6 and IP-10 mainly in response to Pam3CSK4, Poly(I:C)-HMW, LPS, and FSL, whereas the expression pattern of these cytokines was not the same after ligand stimulation in dHepaRG cells and PHH. However, they showed that the antiviral effect of TLR2 and TLR3 ligands is not due to type-I IFN and IL-6 production. In addition to Poly(I:C)-HMW, other ligands, such as Pam3CSK4 and LPS treatments, also significantly decreased both total intracellular HBV DNA and HBeAg secretion in both HBV-infected dHepaRG cells and PHH. However, this study did not include Poly(I:C)-HMW/LyoVec (24). In addition, results of studies on HBV transgenic mice showed that Poly(I:C) is able to inhibit HBV infection, and stimulation of TLR3 pathway has been identified as a potential therapeutic target (13).
Although these recent studies have shown that TLR2 stimulation by Pam3CSK4 in HepaRG cells and PHH could efficiently suppress HBV infection (23,24), another study conducted on transgenic mice showed that TLR2 ligands had little effect on virus replication (13). Our results also showed that stimulation of TLR2 in HepG2-NTCP cells had no effect on HBV infection (Fig. 3), which could be due to the low expression level of TLR2 on HepG2-NTCP cells (Fig. 2A) or impaired TLR2 signaling in this cell line.
Although our results showed that the expression level of TLR4 and TLR7 in HepG2-NTCP cells was very low (Fig. 2A), LPS and R848 significantly reduced HBsAg and HBeAg, but did not affect HBV DNA and cccDNA (Fig. 3). Previous studies have shown a strong inhibition of HBV infection after LPS treatment in PHH, dHepaRG cells, and primary woodchuck hepatocytes, which might be due to the high expression level of TLR4 in these cells (24,40). Result of previous studies showed that R848 and ssRNA, as TLR8 ligands, and also CL264, as TLR7 ligand could not affect HBV infection in HepaRG cells and PHH (23,24), whereas Imiquimod, as TLR7 ligand, significantly inhibited HBeAg and HBV DNA only in dHepaRG cells, but not PHH (24). In contrast, our results showed that R848 significantly reduced HBsAg and HBeAg in infected HepG2-NTCP cells (Fig. 3A, B).
In our study, no inhibitory effect was observed on HBV infection after CpG treatment (Fig. 3). Lack of inhibitory effect on HBV infection by this ligand might be due to the low expression level of TLR9 in HepG2-NTCP cell. A recent study showed that using CpG during HBV inoculation in dHepaRG could prevent the infection, while this inhibitory effect did not correlate with TLR9 activation and/or CpG class. The results of this study showed that this inhibitory effect of CpG was IL-6, IP-10, and TLR9 independent, and perhaps binding of CpG to HBV particles could strongly interfere with HBV entry by coating HBV virions and preventing their interaction with cellular receptor (1). In our study, however, CpG treatment was performed after establishment of HBV infection in HepG2-NTCP cells. Therefore, whether interfering with HBV entry is the only mechanism by which CpG inhibits HBV infection remains to be further investigated. Moreover, using different protocol of treatments, different classes of CpG, and different patterns of innate immunity in these cell lines should be evaluated.
Our results showed that HBV infection could decrease TLR3 expression level and also secretion of IL-8 after Poly(I:C)/LyoVec and Poly(I:C) stimulation, respectively, in hepatocytes (Figs. 4 and 5C). Given the role of TLR3 signaling in restricting HBV infection, it seems a wise strategy that HBV uses to suppress this aspect of innate immunity during Poly(I:C)/LyoVec treatment in HepG2-NTCP cells.
This study is the first report of the effect of TLR/RLR ligands on the induction of APOBEC genes expression in hepatocytes. Based on our results, Poly(I:C)/LyoVec induced the expression of A3F, A3G, and A3H genes in HepG2-NTCP cells. According to the significant inhibitory effect of Poly(I:C)/LyoVec and Poly(I:C) on cccDNA and induction of innate immune response genes, these ligands could be considered as a promising candidate for immunotherapy in CHB patients.
Footnotes
Authors' Contributions
S.A.-A., H.S., M.M.A., M.M., J.K., and F.G.-S. are from Tehran University of Medical Sciences; F.S. is from Tehran University of Medical Sciences and Avicenna Research Institute; and M.J-T. is from Avicenna Research Institute, all where research and education are the primary functions.
Acknowledgments
The authors are grateful to Professor Urlike Protzer, and Professor Christoph Seeger for providing cell lines. They thank Professor Michael Nassal, Professor Masamichi Muramatsu, Dr. Yuchen Xia, Dr. Julie Lucifora, Dr. Suzanne Faure-Dupuy, and Dr. Koichi Watashi for scientific consultation. They also thank Professor Stephan Urban for generously providing Myrcludex B and Theresa Asen for technical support.
Author Disclosure Statement
No competing financial interests exist. This article does not contain any studies with human participants or animals performed by any of the authors.
Funding Information
This study was supported by grants from Tehran University of Medical Sciences (grant numbers: 97-02-27-37246 and 97-02-27-37247) and Iran National Science Foundation (INSF) (grant numbers: 95836480 and 97015900).