
Abstract
The severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) infection may produce a systemic disease, the coronavirus disease-19 (COVID-19), with high morbidity and mortality. Even though we do not fully understand the interaction of innate and adaptive immunity in the control and complications of the viral infection, it is well recognized that SARS-CoV-2 induces an immunodepression that impairs the elimination of the virus and favors its rapid dissemination in the organism. Even less is known about the possible participation of inhibitory cells of the innate immune system, such as the myeloid-derived suppressor cells (MDSCs), or the adaptive immune system, such as the T regulatory cells (Tregs). That is why we aimed to study blood levels of MDSCs, as well as lymphocyte subpopulations, including Tregs, and activated (OX-40+) and inhibited (PD-1) T lymphocytes in patients with mild COVID-19 in comparison with data obtained from control donors. We have found that 20 hospitalized patients with COVID-19 and no health history of immunosuppression had a significant increase in the number of peripheral monocytic MDSCs (M-MDSC), but a decrease in Tregs, as well as an increase in the number of inhibited or exhausted T cells, whereas the number of activated T cells was significantly decreased compared with that from 20 healthy controls. Moreover, there was a significant negative correlation (r = 0.496) between the number of M-MDSC and the number of activated T cells. Therefore, M-MDSC rather than Tregs may contribute to the immunosuppression observed in patients with COVID-19.
Introduction
In December 2019
We now know that SARS-CoV-2 infection may produce a systemic disease, named by the WHO as the coronavirus disease-19 (COVID-19), with high morbidity and mortality, characterized by interstitial pneumonia and SARS, that may progress to multiple organ dysfunction and death (28,33). COVID-19 was declared as pandemic on March 11th, 2020 due to the rapid and uncontrolled expansion worldwide, with more than 188 million cases and more than 4 million deaths in 192 countries at this moment.
The virus produces an early immune suppression by mechanisms not fully understood, but there is a reported lymphopenia in most cases (16,26), which is probably a common feature of other viral infections, although a direct specific effect caused by SARS-CoV-2 should be considered, since it has been shown that some lymphocytes express the ACE2 receptor (the molecule used for the virus to enter the cells) on their Surface (31). Moreover, the virus may enter the cells through other receptors such as CD147 (30). Lymphocyte subpopulations have also been found altered in COVID-19 patients, specially CD4 and CD8 (3,21). Lymphopenia is even worse when the disease progresses to distress respiratory syndrome (24). Actually, decreases in CD4+ T cells, CD8+ T cells, B cells, and natural killer (NK) cells have been observed in peripheral blood of the COVID-19 patients (25,29).
T cells are crucial in the antiviral immune responses. Patients with COVID-19 present an increase in the expression of inhibitory molecules, such as PD-1 or CTLA-4 producing an ineffective immune response (2,34,35). That is why it has been pointed out that even though blockade of the innate immune response and inflammation might be useful in patients with rapid deterioration, it should be taken into account that severe lymphopenia, T cell exhaustion, and consequent adaptive immunosuppression present in these patients may worsen the consequences of targeting innate immunity, including the antibody response (27). Actually, CD4+ T cells are known to promote the production of virus-specific antibodies by activating T-dependent B cells. Therefore, immune modulation has been proposed for the treatment of COVID-19 (6), and it has been found crucial to control and eliminate infection, using adaptive and innate immune responses (7).
Different mechanisms for the lymphopenia in COVID-19 have been recently reviewed (9), including apoptosis, pyroptosis, autophagy, thymus suppression, bone marrow impairment, cytokine storm, activation induced cell death, and antibody-dependent or dendritic cell-dependent killing of lymphocytes. Nevertheless, a possible role of myeloid-derived suppressor cells (MDSCs) has been proposed in the immune suppression of this disease, since expansion of granulocytic MDSCs (G-MDSC) in patients infected with SARS-CoV-2 has been recently described (1). In fact, the role of MDSCs in viral infection has been recently reviewed (19) and, even though the knowledge is limited, broad therapies targeting MDSCs to attain a better balance of antiviral immunity versus immunopathology have been proposed.
MDSCs are immature cells of myeloid origin that express surface markers such as CD33 and CD11b, whereas they are negative for HLA-DR. Besides, they can either express CD14 (monocytic MDSCs [M-MDSC]) or CD15 (G-MDSC) (1). Their immunomodulatory effect has been studied with more detail in the mechanisms of cancer immunoescape (8). They have an immunosuppressive effect on T cells, NK, and dendritic cells by different mechanisms, including the production of free radicals, cytokines, and arginase (4,5,8,20). One of the mechanisms of immune suppression by MDSCs is the induction of the immunosuppressor phenotype in T lymphocytes (TL), T regulatory cells (Tregs), which have also been studied in the immunology of cancer (22). Tregs inhibit the cytotoxic response by inducing the expression of immunosuppressive cytokines, and inhibitory surface proteins, such as CTLA-4 or PD-1.
Therefore, MDSCs and Tregs are good candidates to look for new mechanisms of immunosuppression and new therapeutic approaches in COVID-19. We have analyzed MDSCs, Tregs, as well as B cells, NK, and T cells (CD4+ and CD8+), including the number of activated and inhibited or exhausted TL in peripheral blood from 20 COVID-19 patients and compared with control data from 20 healthy donors.
Materials and Methods
Patients
We have studied the immunological characteristics of peripheral blood leukocytes from 20 COVID-19 successive patients admitted to the Virgen Macarena University Hospital. Twenty healthy donors (from a cohort obtained before the pandemic) were used as control. The study was approved by the Institutional Review Board (ref. MDSC-Treg_COVID-19 Code N. 0908-N-20) according to the ethical principles included in Declaration of Helsinki 1964 (2013 update), and all patients provided oral consent.
Blood samples were obtained within 48 h of the admission. A confirmed COVID-19 case was defined as a patient admitted with respiratory and/or systemic symptoms and positive for real-time reverse-transcriptase polymerase chain reaction (RT-PCR, Allplex 2019-nCoV Assay, Seegene, South Korea) assay for nasal and pharyngeal swab specimens.
Monoclonal antibodies
Antibodies were obtained from Becton Dickinson Immunocytometry Systems (BDIS, San Jose, CA) and were used at the manufacturer's recommended concentrations.
MDSCs: PerCP-Cy5.5 Mouse Anti-Human CD45 (ref no. 564105), APC-Cy7 Rat Anti-CD11b (ref no. 557657), PE Mouse Anti-Human CD33 (ref no. 555450), PE-Cy7 Mouse Anti-Human HLA-DR (ref no. 560651), FITC Mouse Anti-Human CD14 (ref no. 555397), and APC Mouse Anti-Human CD15 (ref no. 551376).
Tregs: Treg kit (ref no. 560249), including PerCP Mouse Anti-Human CD4, PE Mouse Anti-Human CD127, and FITC Anti-Human CD25.
Activated and inhibited TL: FITC Mouse Anti-Human OX-40 (CD134) ref no. 555837, PerCP-Cy5.5 Mouse Anti-Human CD8 (ref no. 565310), PE-Cy7 Mouse Anti-Human CD4 (ref no. 557852), APC Mouse Anti-Human PD-1 (CD279) (ref no. 558694), APC-H7 Mouse Anti-Human CD3 (ref no. 560176).
Flow cytometric analyses in whole blood samples
Lymphocyte subpopulations (T, B, NK, CD4, CD8) were measured by flow cytometry in the routine laboratory using the BD FACSCanto™ flow cytometry system (Becton Dickinson). Peripheral blood samples were collected from subjects in EDTA-K3 tubes to determine MDSCs, Tregs, and OX40+ and PD-1+ TL counts. Cell populations were determined by flow cytometry of whole blood using the BD FACSCanto flow cytometry system. M-MDSC were determined as CD45+CD11b+CD33+HLA-DR-CD14+CD15−, G-MDSC as CD45+CD11b+CD33+HLA-DR-CD14−CD15+ (Fig. 1), Tregs as CD4+CD25highCD127−, activated T cells were determined as CD3+CD4+OX40+PD-1- and CD3+CD8+OX-40+PD-1-, and inhibited T cells as CD3+CD4+OX-40-PD-1+ and CD3+CD8+OX-40-PD-1+. Leukocyte and lymphocyte counts were obtained from hematologic counts (Sysmex CS-1000). The absolute numbers of different leukocyte subsets were calculated by multiplying the percentages with total leukocyte count.
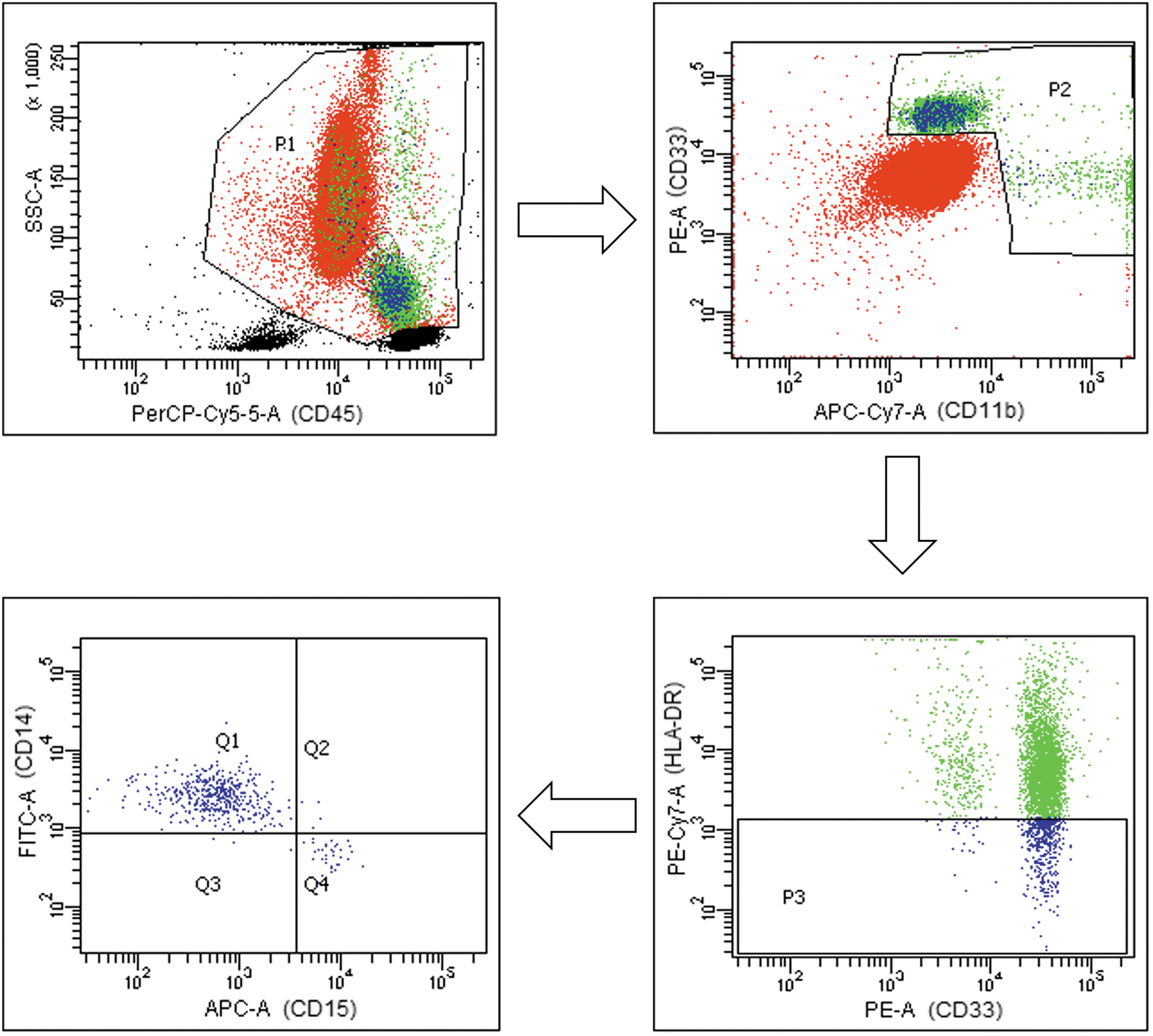
MDSC gating strategy in peripheral blood from COVID-19 patients and healthy controls. Representative plots of the gating strategy for MDSC identification. In SSC/CD45 plot lymphocytes were excluded. Then, CD33+ and CD11b+ cells were selected. Next, in CD33/HLA-DR plot, HLA-DR were selected. Finally G-MDSC are defined as CD15+ and M-MDSC as CD14+. COVID-19, coronavirus disease-19; G-MDSC, granulocytic myeloid-derived suppressor cells; MDSC, myeloid-derived suppressor cell; M-MDSC, monocytic myeloid-derived suppressor cells.
Data analysis
Statistical analysis was performed by SPSS 25.0 software package (SPSS, Inc., Chicago, IL). Normal distribution of analyzed variables was checked by watching histogram, box plot, Q-Q plot-, and the outcomes of normality tests of Shapiro–Wilk. Due to absence of normality, the cell distributions of patients and controls were compared using nonparametric Mann–Whitney U test. Data shown are median and confidence intervals. Statistically significant differences were considered at the 95% level of confidence (p ≤ 0.05). Spearman correlation was used to analyze the association between variables.
Results
Clinical characteristics of COVID-19 patients
Twenty COVID-19 patients and 20 healthy donors were included in the study. Similar age (median, 95% CI) was found in patients: 66.00 (59.01–73.49) to that in control group: 68.2 (62.6–73.6) years old, but higher Male/female ratio percentage of 65%/35 in patients and 50%/50% in control patients and control subjects, respectively. Clinical characteristics of patients are shown in Table 1.
Characteristics of Patients Included
Data are presented as median (interquartile range) or No. (percentage).
Hematological data
COVID-19 patients had no significant changes in total blood leukocytes and platelet levels (Table 2). However, patients have significant increase in the blood concentration of neutrophils and monocytes, whereas we have found a significant decrease in total lymphocyte concentration.
Peripheral Blood Parameters from Coronavirus Disease-19 Patients Compared with Those from Healthy Donors
Data are median (95% confidence interval).
p < 0.05 significant differences in Mann–Whitney U test.
COVID-19, coronavirus disease-19; NK, natural killer; TL, T lymphocytes.
Lymphocyte subpopulations
Lymphocyte subpopulations (T cells, helper and cytotoxic, B cells, and NK) were analyzed by Flow cytometry. As shown in Table 1, COVID-19 patients had significantly decreased T cells, both helper (CD4+) and cytotoxic (CD8+). B lymphocytes and NK were also found decreased in COVID-19 patients, whereas neutrophils were found significantly increased in COVID-19 patients. Monocytes and platelet numbers were not significantly changed in COVID-19 patients.
MDSC and Tregs in COVID-19 patients
Figure 2 shows the median and confidence interval of the MDSC (left panel) and Treg (right panel) number/μL in peripheral blood from COVID-19 patients compared with control healthy donors. Significant increase in MDSCs was observed in COVID-19 patients. This change was due to an increase in M-MDSC, since no differences were found in G-MDSC. The absolute M-MDSC number was threefold higher than in controls. In contrast, the number of Tregs was found to be decreased threefold in peripheral blood from COVID-19 patients compared with control healthy donors.
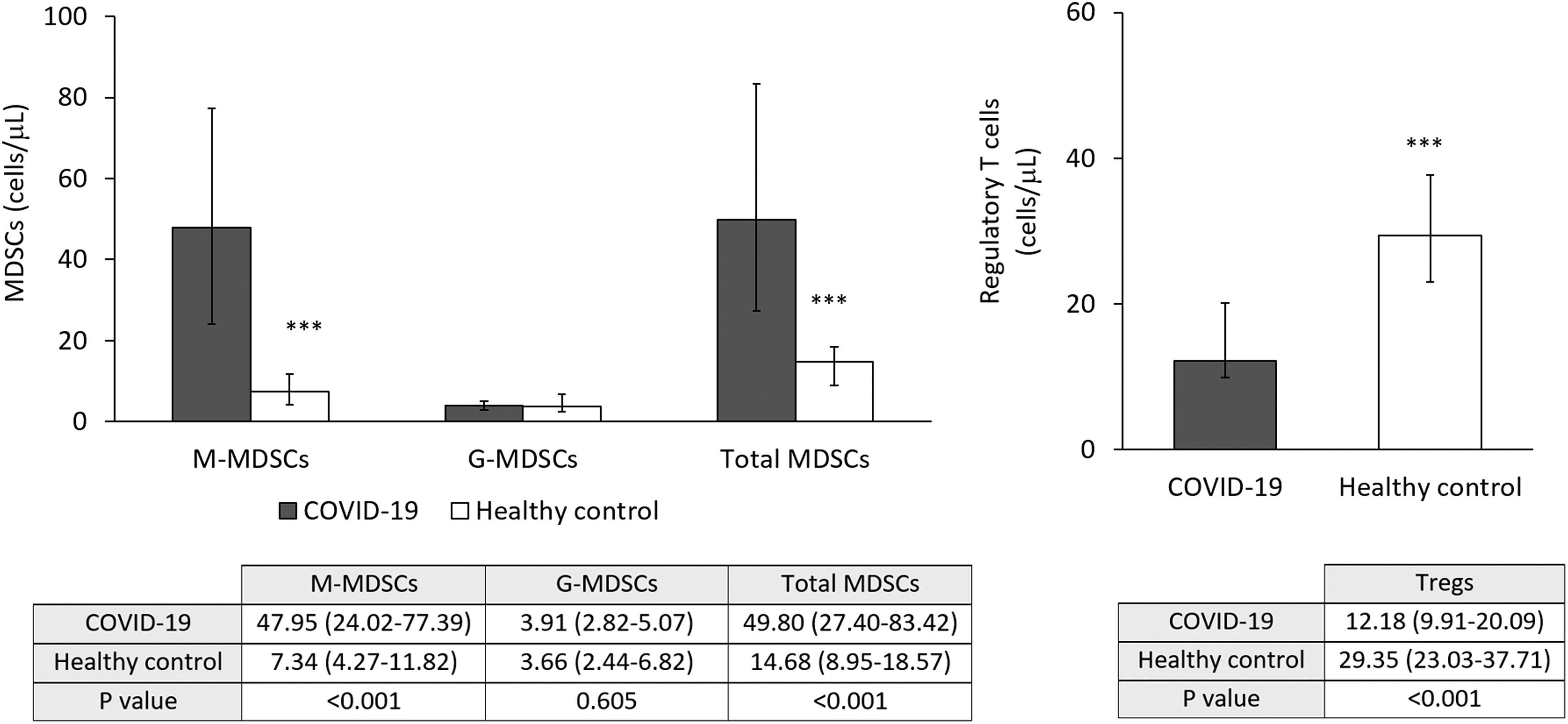
MDSC and Treg in peripheral blood from COVID-19 patients and healthy controls. Left panel: Increased total MDSCs and M-MDSC but not G-MDSC in samples from COVID-19 patients. ***p < 0.01. Right panel: Decreased number of Treg in samples from COVID-19 patients. ***p < 0.01. Treg, T regulatory cell.
Activated and exhausted TL in COVID-19 patients
As shown in Figure 3 (left panel) activated TL (OX-40+) were decreased in COVID-19 patients, compared with healthy control subjects. This difference was due to the decrease in both activated CD8+ TL and the number of activated CD4+ TL in COVID-19 patients, although the decrease was higher in CD8+ TL. Moreover, there was a negative correlation between the number of M-MDSC and activated TL (r = -0.496 [p = 0.031]).
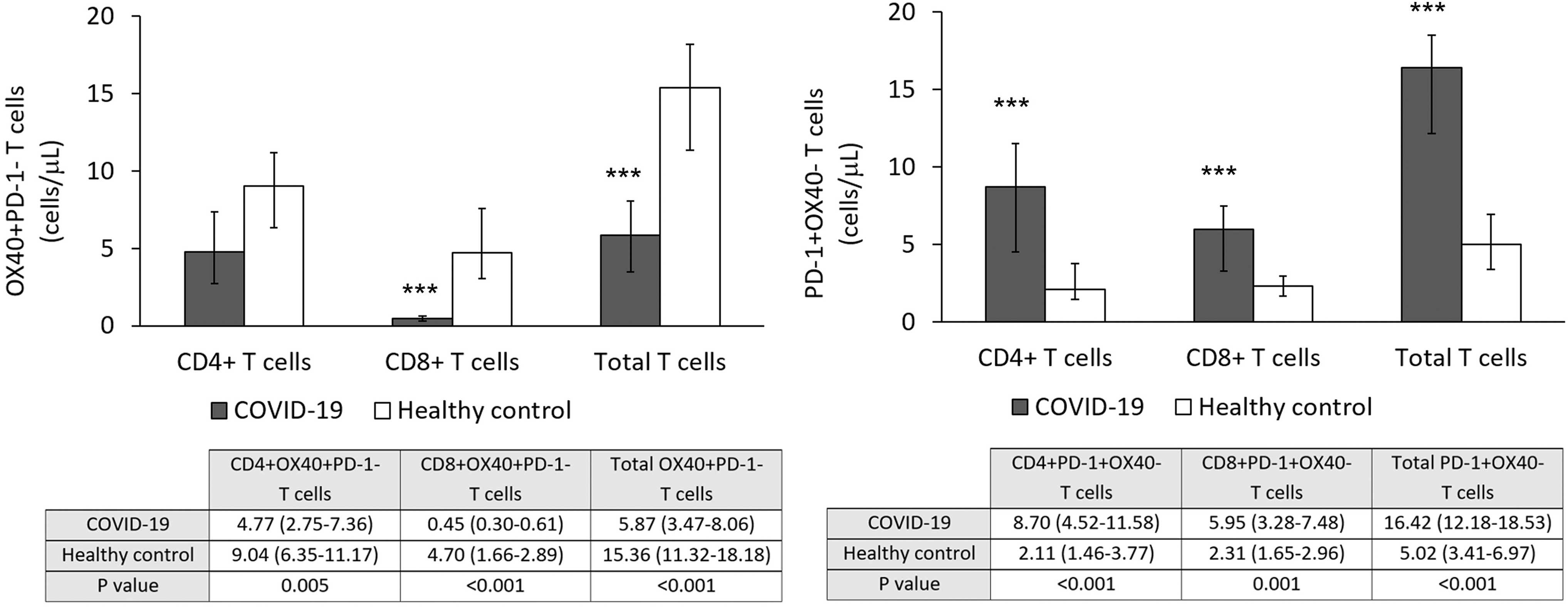
MDSCs, Tregs, and activated and exhausted T cells in peripheral blood from COVID-19 patients and healthy controls. Left panel: Decreased number of activated CD8+ but not CD4+ TL in peripheral blood of COVID-19 patients. ***p < 0.001. Right panel: Increased number of exhausted or inhibited TL, both CD4+ and CD8+. ***p < 0.001. TL, T lymphocytes.
Finally, the number of exhausted T cells (expressing PD-1) in peripheral blood from COVID-19 patients was significantly increased in both CD4+ and CD8+ TL compared with that observed in control subject, as shown in Figure 2 (right panel).
Discussion
SARS-CoV-2 infection may produce a systemic disease named COVID-19, which has been found to produce immunopathological characteristics (29). Peripheral blood leukocyte populations were consistently similar to those previously found in Covid-19 patients; increased neutrophil number, low lymphocyte count, and mild increase in monocyte number (13,29).
Lymphopenia is the most frequent and notable laboratory dysfunction in patients with COVID-19 (13,16,26). We have also found lymphopenia in all the COVID-19 patients studied, and the lymphopenia affects every lymphocyte subset as previously described (T cells, both CD4+ and CD8+, B, and NK cells) (11,25,29). This effect of the viral infection could be due to a direct effect of the virus, although only a small percentage of lymphocytes seem to express ACE2 receptors (31), or indirect, by cell redistribution through chemotaxis, or apoptosis by activating some signaling, as previously suggested for SARS-CoV-1 (32). Besides, cytokines produced upon SARS-CoV-2 infection (particularly IL-6 and IL-8) may impair the ability of T cells to prime dendritic cells, thus leading to a failure to promote an effective adaptive immune response (12,23). In this line, we have found decreased activated and increased exhausted T cells as previously described (34,35). Nevertheless, our hypothesis was that the SARS-CoV-2 virus could recruit innate suppressive immune cells such as MDSCs to participate in the immunodepression of COVID-19 patients. In fact, a general role of MDSCs has been proposed in viral infection (19), in a similar way to that described for cancer (8). In this context, an increase in MDSCs has recently been found in COVID-19 patients (1), although they found only expansion of G-MDSC, and not M-MDSC, as we have found. There must be some technical differences that may explain this discrepancy. For instance, we have determined MDSCs in total blood, whereas the previous study analyzed isolated peripheral blood mononuclear cells by density gradient centrifugation. Nevertheless, this discrepancy further supports further studies of MDSC in COVID-19 patients. In any case, the role of MDSCs in COVID-19 is not specific, since it may be a general mechanism of immunosuppression in viral infection (19), which may be mediated by the alteration of adaptive and/or innate immune response, as suggested for influenza virus (10).
In contrast, Tregs, which are promoted by MDSCs, have been found decreased in the COVID-19 patients studied. This result is consistent with previous data regarding the dysregulation of immune response in patients with COVID-19 (21), but other groups have found increased number of Tregs, especially in severe COVID-19 patients. We have no clear explanation for this result of decrease in Treg number. Eventually, the lymphopenic effect of the SARS-CoV-2 infection may also affect the Treg population. In contrast, it could be that the effect of MDSC on the production of Tregs needs further exposition to the infection and evolution of the disease, or alternatively, Tregs may be sequestered in inflammed tissue such as the inflamed lung and that is why the number is even lower in peripheral blood. This is a point that warrants further investigation.
A possible limitation of our study is the low number of patients studied. Nevertheless, data seem clear enough to conclude that viral infection with SARS-CoV-2 in COVID-19 patients produces lymphopenia that affects every lymphocyte subset and decreases the number of activated CD8 and CD4 lymphocytes. In fact, there was a significant inverse correlation between the number of M-MDSCs and the number of activated TL. Moreover, the infection also increases the number of exhausted CD4 and CD8 lymphocytes, which further impair the immune response.
We have not carried out further analytical determinations since the course of the disease in the patients studied was benign. Further studies with a higher number of patients could help to consider the parameters analyzed as prognostic markers.
Finally, according to the data obtained in the present work, these immunosuppressive effects observed in COVID-19 patients may be, at least in part, mediated or helped by the observed increase in M-MDSC in these patients.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
V.S.-M. received support from Junta de Andalucia and University of Seville (PAIDI CTS-151 group), A.P. and J.R.-B. receive support for research from Plan Nacional de I+D+i 2013–2016 and Instituto de Salud Carlos III, Subdirección General de Redes y Centros de Investigación Cooperativa, Ministerio de Ciencia, Innovación y Universidades, Spanish Network for Research in Infectious Diseases (REIPI RD16/0016/0001), cofinanced by European Development Regional Fund “A way to achieve Europe,” Operative Program Intelligence Growth 2014–2020. LdC-M received support from the Asociación de Mujeres con Cáncer de Mama (AMAMA Sevilla), Fundación Sandra Ibarra, and Asociación de Cáncer de Mama de Brenes.