
Abstract
Porcine reproductive and respiratory syndrome virus (PRRSV) induces apoptosis through the activation of death receptors, including cell-surface Fas receptor. The aim of this study was to investigate the impact of intradermal (ID) and intramuscular (IM) vaccination with a commercial PRRSV-modified live vaccine in piglets on Fas-related apoptosis. The study included 104 suckling piglets from a commercial farrow-to-finish pig farm, suffering from positive unstable PRRSV status. Animals were assigned in four groups: group A—Porcilis PRRS ID-vaccinated pigs, group B—Porcilis PRRS IM-vaccinated pigs, group C—Diluvac ID adjuvant-administered pigs, and group D—Diluvac IM adjuvant-administered pigs. Vaccines were administered at 2 weeks of age. Blood samples were collected from the same pigs at 4, 7, and 10 weeks of age. Sera were examined by quantitative real-time reverse transcription-PCR (qRT-PCR) for PRRSV and by ELISA for soluble Fas (sFas). At 4 weeks of age, all groups were negative qRT-PCR for PRRSV; at 7 weeks only group A was negative; and at 10 weeks all groups were positive. sFas was significantly increased in groups C (4 vs. 7, 4 vs. 10, and 7 vs. 10 weeks) and D (7 vs. 10 weeks). Significant differences among groups were noticed only at 10 weeks (A vs. C, A vs. D, B vs. C, B vs. D). A significant positive and moderate correlation between PRRSV viral load and Fas level was observed. In unvaccinated piglets, increased serum sFas levels reveal apoptotic suppression compared with vaccinated piglets. In the latter, vaccine-derived antibodies limit the infection and may attribute to the reduced Fas expression, suggesting a weak induction of lymphocyte-mediated cytotoxicity.
Introduction
Porcine reproductive and respiratory syndrome virus (PRRSV) remains a major cause of significant economic loss in the swine industry (44). PRRSV causes reproductive failure in the breeding herd and respiratory disease and growth retardation in the pigs (65). Vaccination is considered as a crucial measure for the prevention and control of PRRSV infection. It is also the most cost-effective strategy for all sizes of pig farms compared with other control strategies (28,45).
Intradermal (ID) vaccination has the advantage of targeting antigen-presenting cells in the epidermis near skin-draining lymph nodes (46,57). Many studies have already proved the safety and efficacy of ID vaccination against major pathogens in swine, such as PRRSV (2,33,34,50), porcine circovirus 2 (PCV2) (49), Aujeszky's disease virus (12,61), and Mycoplasma hyopneumoniae (4,55). Combined with needle-free administration, ID vaccination is more animal friendly and prevents accidental transmission of pathogens caused by reusing needles as well as muscle damage by broken needles in the muscle.
Previous studies demonstrated that route of administration of the vaccine does not affect the degree of virological protection, as pigs intradermal (ID) vaccinated did not show any differences in terms of viral load in their blood and clinical signs when compared with animals intramuscular (IM) vaccinated (33,34).
Previous reports confirmed that a PRRSV-modified live vaccine (MLV) administered through the intradermal route can efficiently induce a protective response in pigs subsequently exposed to a genetically diverse PRRSV isolate even when applied in field conditions (33). This means that vaccine administration through either the intradermal or the intramuscular route reduces the viral titers of PRRSV in the blood and in BAL fluid, can confer some degree of virological cross-protection to heterologous isolates of PRRSV (26). Also, a recent study suggested that a European type 1 (PRRSV-1) PRRSV MLV administered by either IM or ID can provide partial heterologous protection after challenge with a highly pathogenic PRRSV type 2 (PRRSV-2) strain, either alone or combined with PRRSV-1, as demonstrated by less lung tissue lesions and reduced viremia (31).
In addition, needle-free applications have demonstrated some advantages such as safety for the executor, painlessness for the pig, absence of histological lesions, and no iatrogenic transmission of pathogens (33). It has been demonstrated that 28 h after vaccination, IM-vaccinated piglets against PCV2 had lower levels of blood C-reactive protein and blood haptoglobin compared with IM-vaccinated ones (56).
Another advantage is that ID vaccine administration might represent an alternative to enhance the efficacy of vaccination against PRRSV, as it induced lower IL-10 levels after vaccination and more IFN-γ-SC after the experimental challenge of animals with a heterologous viral strain alone or even combined with a homologous strain infection (31). Notable welfare-associated advantages of ID vaccination versus IM vaccination of piglets have been demonstrated in recent studies. ID vaccination against M. hyopneumoniae (14) and PCV2 (56) resulted in fewer postvaccination behavior and activity changes in suckling piglets compared with intramuscular vaccination.
Pulmonary alveolar and/or intravascular macrophages are known as the major target cells for PRRSV in the lungs (58). The studies on PRRSV infection and replication in host alveolar macrophages continue to give conflicting results as to its effect on innate cell activation, antigen presentation, and the triggering of acquired immunity (20,21,23,25,27,59,60). It has been widely demonstrated that PRRSV causes apoptosis in different organs (lungs, testes, lymph nodes, and thymus) as well as in cell lines (5,6,11,22,24,48,51,52). On a morphological basis, the apoptotic cells are assumed to be alveolar and pulmonary intravascular macrophages and mononuclear cells in the alveolar septa. Double labeling experiments indicate that most apoptotic cells are noninfected (48,52). An infection of the lungs with PRRSV has some peculiar cellular features that may be attributed to apoptosis.
First, it is characterized by changes in the population of bronchoalveolar monocytes/macrophages, consisting of a reduction of the population of well-differentiated macrophages and an influx of new monocytes (21). In general, apoptotic pathways can be divided into two nonexclusive signaling cascades: extrinsic pathway and intrinsic pathway (7). Extrinsic or “death” receptor-mediated apoptosis is initiated when a soluble ligand (such as tumor necrosis factor [TNF]; Fas ligand [FasL]; or TNF-related apoptosis-inducing ligand [TRAIL]) binds to specific receptors on the cell surface (1,38).
The cell-surface Fas receptor, also termed APO-1 or CD95, is a member of the TNF and nerve growth factor family of receptors (39,47). Fas expression is observed in a variety of immune and nonimmune cells (62). Previous studies have covered the mechanisms underlying cell death induced by Fas/FasL interaction (1,19,32,43).
The aim of this study was to investigate the Fas (APO-1)-related apoptosis in piglets after intradermal (ID) and intramuscular (IM) vaccination with a PRRSV MLV in comparison with nonvaccinated piglets.
Materials and Methods
Ethics
Throughout this clinical study, every procedure was performed according to the Guide for the Care and Use of Agricultural Animals in Research and Teaching as well as the Code of Practice for the Conduct of Clinical trials for Veterinary Medical Products; all animals were housed and handled in compliance with National and European animal Welfare requirements (8,9,10,42). All procedures and animal use in this study were approved by the Ethical Committee of the Faculty of Veterinary Medicine, School of Health Sciences, University of Thessaly (Approval number: 98/December 19, 2019).
Farm
General requirements for the selection of the farm were the presence of a PRRSV laboratory-confirmed infection and good farm management practice (i.e., accurate and up-to-date administration and all-in/all-out system).
This study was carried out in a farrow-to-finish commercial pig farm (commercial hybrids of Large White × Landrace). The capacity of the farm was 150 sows under production, located in Central Greece. A grandparent nucleus of 10 sows was kept in the farm for producing its own gilts. The farm facilities included farrowing houses, flat-deck units, growing and finishing houses, one mating-pregnancy (dry period) stable with individual stalls (35th–105th day of pregnancy), breeding stock house of group housing (>35th day of pregnancy), breeding stock house of group housing for noninseminated gilts, a feed mill, and an artificial insemination (AI) laboratory. The herd practiced a 1-week batch production system.
The weaning piglets were allotted equally according to the body weight and sex at random at flat-deck batteries for piglets in a climate-controlled postweaning stable. Weaned sows were housed in individual stalls, and they were inseminated twice with fresh semen from the same boar. Semen collection, dilution, and storage were performed on the farm (system “Do-it-yourself AI”). At 30–35 days of gestation, sows were moved to group housing rooms where they were housed in 10 animal groups. Sows were fed individually using separate feeding stalls. Gilts were also housed individually during the first month of gestation.
The routine vaccination program of breeding stock included vaccinations against PRRSV, Aujeszky's disease virus, swine influenza virus, porcine parvovirus, Erysipelothrix rhusiopathiae, Escherichia coli, and Clostridium perfringens. Gilts were vaccinated at age of 180th and 210th days and the sow population followed a 6–60 vaccination plan against PRRSV with the commercial vaccine (Porcilis® PRRS, MSD Animal Health). The weaned piglets were vaccinated against M. hyopneumoniae and PCV2 at 21 days of age. All breeding females were treated with a single ivermectin injection 14 days before farrowing; boars were treated twice a year.
Feed provided to the animals was self-prepared meal, based on a corn/barley/wheat-soya combination, depending on the season. Breeding animals received differentiated meal during gestation and lactation, based on the needs of the animals for each production stage.
Animals had ad libitum access to drinking water. There was one drinking nipple per sow during lactation and one nipple in each crate during gestation. Housing facilities were equipped with a fully automated temperature and humidity control system, as well as an automated feeding system.
Farm history—pretrial period
One month before the beginning of the trial, blood samples were collected from the breeding stock (12 total blood samples from gilts, sows in the dry period, and in lactation), as well as from suckling, weaning, growing, and finishing pigs (8 blood samples at age of 2, 4, 7, 10, 13, 17, and 21 weeks). The presence of PRRSV infection was investigated by quantitative real-time reverse transcription-PCR (qRT-PCR), and the wild-type PRRSV strain circulating in the study farm was characterized by complete ORF5 (Gp5) gene sequencing and phylogenetic analysis, as described below under the section “Laboratory examinations.”
Animals
In total, 104 suckling piglets of 2 weeks of age were included in the study and were divided into 4 groups (Table 1) and 2 replicates, including 13 piglets per group (2 replicates × 13 piglets × 4 = 104 piglets). Only healthy suckling piglets of 2 weeks of age were included. Diseased piglets, piglets significantly underweight, runts, and animals with abnormalities (e.g., hernias) were excluded. All piglets were identified with two unique numbered ear tags. Piglets belonging to all groups were mixed and housed as usual.
Results of qRT-PCR in Serum Samples of the Studied Animals at Ages of 4, 7, and 10 Weeks
PRRS ID, intradermal vaccination with Porcilis PRRS; PRRS IM, intramuscular vaccination with Porcilis PRRS; PRRSV, porcine reproductive and respiratory syndrome virus; qRT-PCR, quantitative real-time reverse transcription-PCR.
Study design
Experimental material
Porcilis PRRS (MSD Animal Health) was the commercial vaccine used in the study for the immunization of piglets against PRRSV. The vaccine was administered through both intramuscular and intradermal ways as stated in product SPC. It is a live attenuated lyophilized vaccine containing per dose of 2 mL (intramuscular application) or 0.2 mL (intradermal application) of reconstituted vaccine: Lyophilisate (active substance)—live attenuated PRRSV strain DV: 104.0–106.3 TCID50 (50% tissue culture infective dose), solvent (Diluvac Forte® adjuvant)—dl-α tocopheryl acetate: 75 mg/mL. In control groups, only Diluvac Forte, containing dl-α tocopheryl acetate was administered.
Experimental groups, dosage, and administration
The following four groups were created: group A—Porcilis PRRS ID, group B—Porcilis PRRS IM, group C—Diluvac ID, and group D—Diluvac IM. Piglets belonging to groups A and B were vaccinated at 2 weeks of age with 1 dose of Porcilis PRRS, diluted in 0.2 mL of Diluvac Forte and injected intradermal (ID), or diluted in 2 mL of Diluvac Forte and injected intramuscular (IM). Piglets belonging to groups C and D were not vaccinated against PRRSV. Only 1 dose of the recommended volume of adjuvant (Diluvac Forte) was administered to each animal belonging to both latter groups; specifically, 0.2 mL for ID-negative controls (group C) and 2 mL for IM-negative controls (group D).
Per treatment group, an IDAL (IntraDermal Application of Liquids) vaccinator was used for needle-free intradermal vaccination of ID groups. For the vaccination of the IM groups, an automatic syringe, with a fixed 2 mL volume, was used. A new, clean needle (size 7, ∼0.9 × 13 mm) was used for each IM trial group.
All other treatments of the growing-fattening pigs (preventive oral or injectable treatments and vaccinations) were recorded for each treatment and control group.
Samplings
During the trial, blood samples were collected from three same ear-tagged pigs of each group for two replicates, at the age of 4, 7, and 10 weeks, as shown in Figure 1.

An overview of experimental design, samplings, and tests performed.
Laboratory examinations
Blood serum samples obtained from both the pretrial period and the trial (investigation) period underwent nucleic acid extraction using the PureLink®Viral RNA/DNA Mini Kit (Invitrogen, Carlsbad, CA). Extracts were tested for PRRSV genome, using a previously described TaqMan probe-based qRT-PCR (30). Reactions were carried out on a CFX96® Real-Time System (Bio-Rad Laboratories, Hercules, CA). The obtained cycle threshold (Ct) values were used as viral load estimates regarding the samples obtained during the trial period.
In addition, the wild-type PRRSV strain circulating in the study farm was characterized by complete ORF5 (Gp5) gene sequencing and phylogenetic analysis, including sequence comparison with the vaccine strain used in the study. In brief, a qRT-PCR–positive RNA extract was reverse transcribed into cDNA using random hexamers. The viral ORF5 sequence was amplified by PCR, employing the primers Gp5-SeqUp (5′-ATGAGGTGGGCYACAACCATYGC-3′) and Gp5-SeqDo (5′-AGGGCRTATATCATTATRGGTGTGTATGT-3′). Sanger sequencing of the purified product was performed on an ABI 3730xl Genetic Analyzer (Applied Biosystems, Darmstadt, Germany). The obtained ORF5 sequence was assembled and imported into MEGA 6 software (53).
Additional homologous sequences of representative European-type viral strains were retrieved from GenBank, including the sequence of the strain contained in the vaccine used (strain DV; Acc. no. KF991509), and were included in the dataset.
The sequences were aligned by MUSCLE. The best fitting nucleotide substitution model was determined based on the lowest Bayesian information criterion score. A maximum-likelihood tree was constructed using the Kimura 2-parameter model with gamma-distributed rates across sites (K2+Γ) and the nearest neighbor interchange tree searching strategy. The reliability of the hypothesis was assessed using bootstrap resampling with 1000 replications. Pairwise genetic distance calculation was performed by MEGA 6.
Sera were also tested for soluble Fas (sFas) levels using a commercially available ELISA Kit (Pig Factor-related Apoptosis, ELISA kit Porcine FAS/APO-1, Cusabio) following the manufacturer's instructions.
Statistical analysis
Data were expressed as mean ± standard deviation. Comparisons were performed using Kruskal–Wallis's test while the Mann–Whitney U test was used for post hoc/multiple comparisons. Comparisons between more than two measurements of the same group during the time were performed using Friedman's test and Wilcoxon's signed-rank test as a post hoc test. All tests were two sided. p < 0.05 was considered to indicate a statistically significant difference. Statistical analysis was performed in IBM SPSS 21 (IBM Corp., Armonk, NY).
Correlation between viral load as determined by qRT-PCR and the Fas levels in the blood was estimated by Spearman's correlation coefficient (ρ), employing the commercial MedCal 9.2 software (MedCalc Software, Mariakerke, Belgium). Statement of significance was based on p ≤ 0.05. The strength of the relationship was evaluated as follows: ρ ≤ 0.35—weak correlations, 0.36 to 0.67—moderate correlations, and 0.68 to 1.0—strong correlations (55).
Results
Virological testing
During the pretrial period, the presence of PRRSV infection was confirmed by qRT-PCR in blood serum samples. At an age of 4, 7, 10, and 13 weeks, 100% of the tested pigs were qRT-PCR positive. The full-length ORF5 sequence of the wild-type PRRSV strain detected in the study farm during the pretrial period was successfully determined. Comparison of the obtained sequence with the respective sequences of other European genotype strains available in GenBank revealed nucleotide sequence identity of 90.7% to the strain DV; that is, the strain included in the MLV used in the study (acc. no. KF991509) and of 90.8% to “Lelystad virus”; that is, the prototype European genotype strain (acc. no. M96262) (Fig. 2).
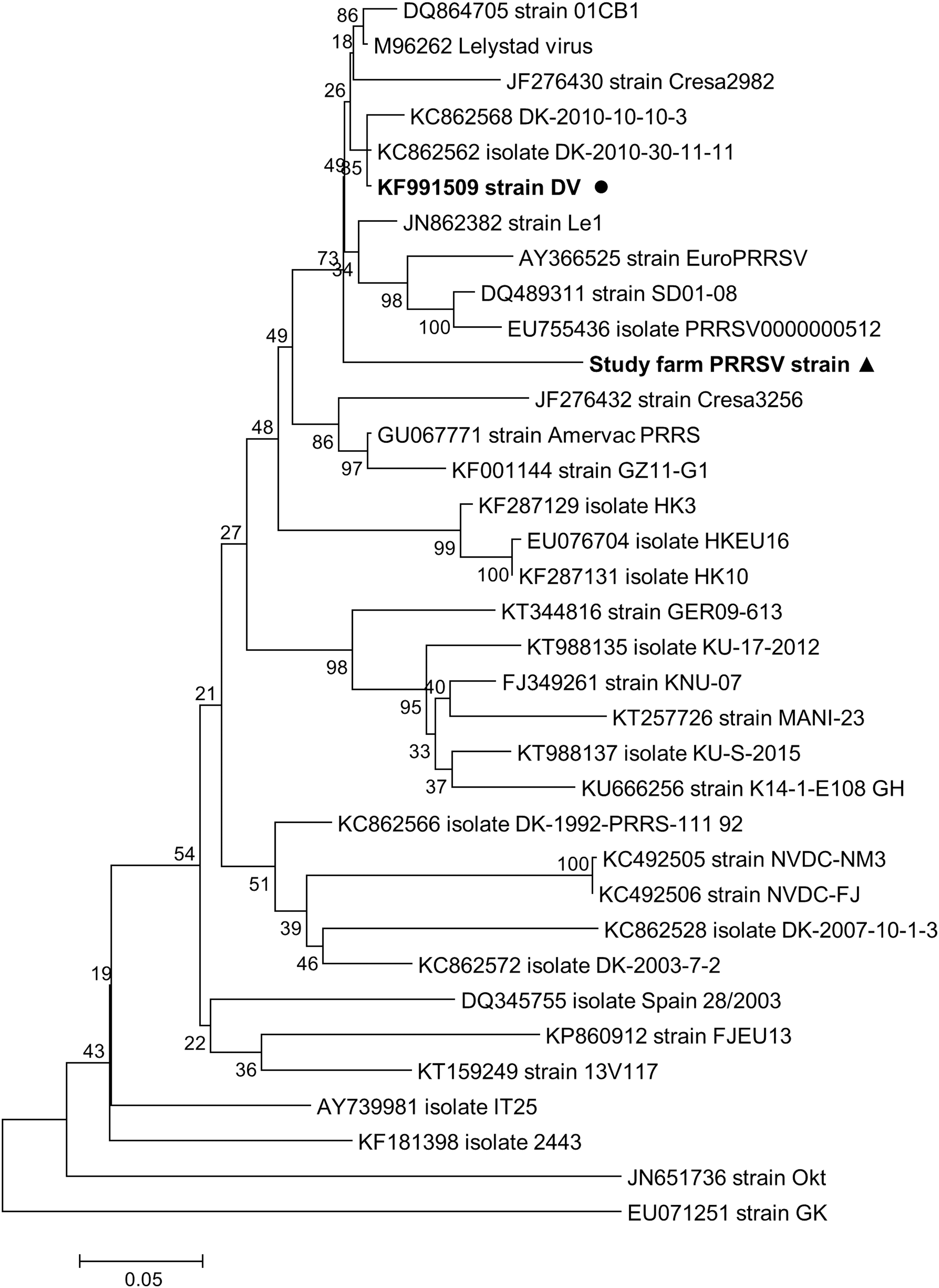
Phylogenetic tree inferred with ML analysis based on complete ORF5 (Gp5) sequences of selected European-type viral strains. Each taxon label includes GenBank accession number and isolate/strain name. The Kimura 2-parameter model with gamma-distributed rates across sites (K2+Γ) was selected as the best fitting nucleotide substitution model for this dataset. The tree is rooted at the midpoint, and the numbers indicated on the branches are nonparametric bootstrap probabilities generated from 1,000 iterations. The scale bar indicates nucleotide substitutions per position. The sequence obtained from the wild-type virus circulating in the study farm is indicated with a triangle, and the sequence of the DV vaccine strain is indicated with a circle. ML, maximum likelihood.
The results of qRT-PCR analysis in the blood samples obtained during the study period are shown in Table 1. Ct values are presented in Figure 3. Samples obtained at 4 weeks of age were negative in all groups. At 7 weeks, only group A was negative, and at 10 weeks all groups were positive. However, a narrower Ct value range and a slightly lower mean Ct value were observed in group A compared with group B at 10 weeks (Fig. 3).

Ct values of qRT-PCR in serum samples at ages of 7 and 10 weeks of PRRS ID- and IM-vaccinated piglets in comparison with unvaccinated piglets. Ct, cycle threshold; PRRS ID, intradermal vaccination with Porcilis PRRS; PRRS IM, intramuscular vaccination with Porcilis PRRS; qRT-PCR, quantitative real-time reverse transcription-PCR.
Fas
The results for Fas (pg/mL) are shown in Table 2 and Figure 4. No differences in sFas levels were observed in groups A and B throughout the study. Levels of sFas increased in group C during time (4 vs. 7 weeks—p = 0.028; 4 vs. 10 weeks—p = 0.028; 7 vs. 10 weeks—p = 0.046) and D (7 vs. 10 weeks—p = 0.028). Significant differences among groups were noticed only at 10 weeks (A vs. C—p = 0.010; A vs. D—p = 0.055; B vs. C—p = 0.010; B vs. D—p = 0.037).
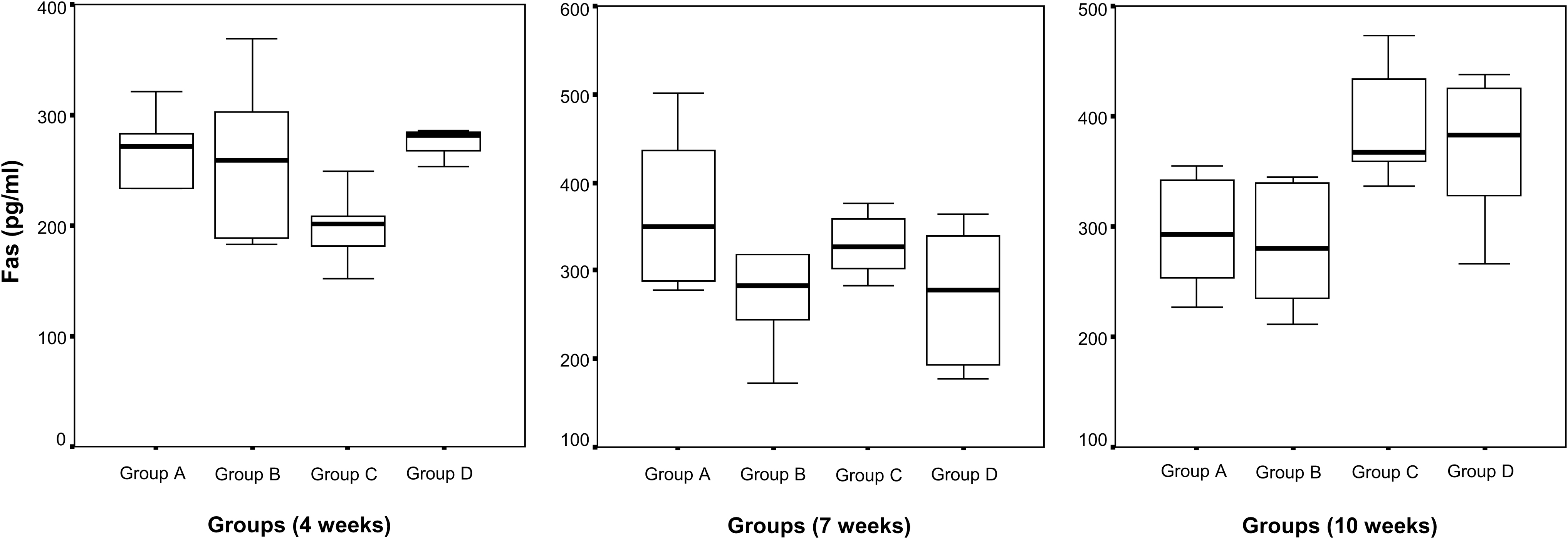
Fas levels (pg/mL) in all groups at ages of 4, 7, and 10 weeks of PRRS ID- and IM-vaccinated piglets in comparison with unvaccinated piglets.
Results of Fas (APO-1) Levels (pg/mL) in Serum Samples at Ages of 4, 7, and 10 Weeks
NS, nonsignificant; SD, standard deviation.
Correlation between PRRSV viral load and Fas
Data analysis revealed a statistically significant positive and moderate correlation between PRRSV viral load and Fas level (ρ = 0.639, p < 0.0001).
Discussion
Previous studies have determined the regulatory roles of PRRSV in apoptosis that is important in the pathogenesis of PRRSV (16,18,24,51,52). In our study, increased serum sFas levels in unvaccinated piglets were recorded, revealing apoptotic suppression compared with vaccinated piglets. In the latter, vaccine-derived antibodies limit the infection and may attribute to the reduced Fas expression, suggesting a weak induction of lymphocyte-mediated cytotoxicity. Finally, higher sFas levels were observed with aging, possibly due to persistent PRRSV infection.
Virus-infected cells involved apoptosis in eliminating themselves to prevent viral replication, viral dissemination, or persistent infection (41). Thus, it is vital for viruses to disable host cell apoptosis at an early stage of the infectious cycle, thereby avoiding premature cell death and allowing viral replication. On the contrary, apoptosis at a late infection stage could function promotively to virus spread by breaking down infected cells or killing uninfected cells of the immune system (15).
Former studies demonstrated biphasic apoptosis during virus infection (17,64). In contrast with a typical acute viral infection resolving in 1–2 weeks by an effective clearance, PRRSV gives rise to a long-term infection, characterized by prolonged viremia and persistent infection. The immune response to PRRSV is partially ineffective, including a relatively low or altered activation of the inflammatory and innate response, which has an important role in accounting for the persistence of PRRSV infection and its low and retarded activation of acquired immune response (3,29,37,63). PRRSV-1 has been shown to possess a dichotomous apoptotic role in manipulating apoptosis, which is inhibited early but induced late in infection (6).
Fas/FasL demonstrated a major role in the induction of apoptosis in immune and bronchial epithelial cells (13,35,40). Fas/FasL interactions induce apoptosis of immune cells, including T, B cells, and macrophages, and cell death is often associated with activated stages of immune cells. Activation-induced cell death is a mechanism to inhibit excessive immune responses and to terminate ongoing immunity.
The analysis of our results demonstrated for the first time a significant positive and moderate correlation between PRRSV viral load and Fas levels. This finding supports that PRRSV interferes with the induction of cellular innate immune response, pathways of which are tied closely with that of apoptosis (36). This may explain why there is a delayed immune response in PRRSV-infected pigs, and elucidation of the mechanisms of interference may provide clues into novel intervention strategies to combat PRRSV infection (36).
In conclusion, our study indicated increased serum sFas levels in unvaccinated piglets, which could reveal apoptotic suppression compared with vaccinated piglets. No significant differences were noticed for serum sFas levels between ID and IM vaccination. Moreover, in the latter, vaccine-derived antibodies limit the infection and may attribute to the reduced Fas expression, suggesting a weak induction of lymphocyte-mediated cytotoxicity.
Footnotes
Author Disclosure Statement
G.M. was employed by MSD Animal Health from April 2018 to April 2019.
Funding Information
This work was partially supported by Intervet Hellas AE—MSD Animal Health through the Research Committee of the University of Thessaly (code: 5168, Scientific Responsible: Associate Professor V. Papatsiros).