
Abstract
New strategies to increase the immune response to HIV-1 vaccine using immunological adjuvants such as Toll-like receptor agonists are needed. In this study, HIV-1 p24-Nef and conjugated form of the vaccine candidate to type-A flagellin (FLA) were injected in the BALB/c mice in different routes. Two weeks after the last immunization, lymphocyte proliferation was measured by the BrdU method. The IL-4 and IFN-γ levels, as well as the total IgG antibody and its isotypes titer, were evaluated by the enzyme-linked immunosorbent assay method. The IFN-γ ELISPOT was also performed. Our data showed that the HIV-1 p24-Nef alone and conjugated to type-A flagellin (FLA) significantly increased lymphocyte proliferation responses as well as higher levels of cytokines and IFN-γ producing lymphocytes and the level of humoral immune responses compared with the control groups. The cell-mediated immune responses through the subcutaneous route and humoral immune responses through the intramuscular route were significantly higher in the conjugated form than in the mere vaccine candidate. In conclusion, when the FLA as an adjuvant is constructed in the HIV-1 vaccine candidate, it could effectively improve both humoral and cellular immune responses. Furthermore, modification in the vaccine formulation could change the optimal route of vaccine inoculation.
Introduction
Vaccines are an important product to the control of infectious diseases. To elicit an appropriate and robust immune response, it seems that an adjuvant can increase the immune response against an antigen or main vaccine through various mechanisms (13).
Owing to the beneficial effect of microbial products, research on the use of Toll-like receptors (TLRs) agonists as a new type of adjuvants has concentrated. The binding of these molecules to their receptors on the surface of immune cells, especially antigen-presenting cells (APCs), leads to activation of these cells and triggers inflammatory responses and stimulates naive T cells and polarization into Th1/Th2 immune platforms (11). This characteristic in terms of access to an effective immune response to a vaccine is important, therefore, certain TLR agonists currently have been used with success in formulations of some commercial vaccines (14).
The bacterial flagellin, such as Pseudomonas aeruginosa, has a sequence that can bind to TLR-5 and irritate it extremely. The protein that can act as a ligand for TLR-5 (12,32,34) can induce innate immune responses that are required to set up the acquired immune response, including the production of proinflammatory cytokines and chemokines by lymphoid and nonlymphoid cells, influx of B and T lymphocytes into secondary lymphoid organs, dendritic cells (DCs) activation and induce direct activation of T lymphocytes (14).
Flagellin in many types of bacteria was shown to stimulate strong innate and acquired immune responses and is considered as an immunological adjuvant and even an effective vaccine in humans and nonhuman primates (17,31). However, the protection conferred by the immune system depends on both vaccine formulation and type of immune response, and so control of these two parameters is important for vaccination strategies.
Successful immunization depends on many parameters that can affect vaccine efficacy that one of the most important of them is vaccine administration-related parameters that include immunization route, number of administrations, and intervals between administrations (30). The optimum route of immunization can trigger the highest level of immune responses to establish protectivity (4). Also, the route of delivery could affect the localization of the vaccine that may impact the priming of immune cells as well as consequential local and systemic immune responses (35).
Since the interaction between the vaccine and APCs in terms of base and mechanism in the different injection sites is different, routes of immunization influence the immune response to an antigen (1,4). In each route of administration, APCs and DCs uptake vaccine and present to the T lymphocytes in the lymphoid organs after processing, but according to the injection site, immune polarization and the level of response may vary (26). Thus, the characteristics and power of induced and memory T cells response are dependent on T lymphocytes activation that, in turn, is dependent on the nature of resident APCs (7).
Expression of homing molecules on the lymphocytes is greatly influenced by the microenvironment of inducing the immune response. DCs in each region conduct expression of homing receptors on the T lymphocytes that migrate into those points (6). It seems the nature of the T cell homing receptor is affected by tissue microenvironment and tissue-specific DCs, and the microenvironment of immune system immunization gives specific traffic characteristics to the responsive CD8+ T cells that may be the property of regional APCs (6,21). Also, differences in the nature of the APCs in each route have different effects on T lymphocytes assisting pattern.
In a way that has been observed in the dermal route, CD8+ T cells do not need CD4+ T cells to help, whereas in the intraperitoneal route, CD8+ T cells require CD4+ T cells because of dendritic and Langerhans cells in the dermal route and peritoneal macrophages in the intraperitoneal route that are involved in the presentation of antigens. These macrophages cannot directly activate CD4+ T cells, whereas DCs directly can (2).
Also, studies have shown that the parameters of the memory B lymphocytes that are associated with protective immunity can be related to the injection route of the vaccine (19), and even the way that vaccine is injected can determine which tissue responds. For example, in the intravenous route, the spleen, and in the intramuscular (i.m) route, lymphoid organs and spleen are responding tissue (1,7). Thus, according to the differences in DCs phenotype and their basic function at the beginning of immune responses, the selection of vaccine injection site is particularly important.
In this study, HIV-1p24-Nef peptide vaccine as a vaccine model in two formulas as mere vaccine and conjugated to flagellin protein in the subcutaneous (s.c), intradermal (i.d), and i.m routes was used. Herein, we hypothesized that the change in the vaccine formulation may affect its effective route, so after immunization of experimental animals, immunological parameters of each formulation of vaccine and immunization routes were compared.
Materials and Methods
Experimental mice and animal ethics
Six- to 8-week-old inbred female BALB/c mice (n = 51) were purchased from Pasteur Institute of Iran (Karaj, Iran). The mice were housed for 1 week before the experiments, given free access to food and water, and maintained in a standard light/dark cycle (12 h/12 h) and at 20–22°C. All the mouse experiments were done under the animal care and use protocol of Pasteur Institute of Iran and Shahrekord University of Medical Sciences with ethics code 89-11-3.
HIV-1 p24-Nef fusion peptide
The HIV-1 p24-Nef fusion peptide from p24 (aa159–173) and Nef (aa102–117) proteins was synthesized according to the solid-phase method by GL Biochem Company (Shanghai, China). The peptide was purified using high-performance liquid chromatography (HPLC) to obtain a peptide with a purity exceeding 95% for experiments.
Production and purification of the recombinant flagellin
In previous studies, the type-A flagellin (fliC) gene was cloned and expressed as the histidine-tagged protein in a bacterial expression system (8,27). In brief, the P. aeruginosa fliC gene was first cloned into the pET28a expression vector and then expressed in Escherichia coli BL21 (DE3) and purified by Ni2+ affinity chromatography. The purified protein was passed through the LPS (lipopolysaccharide) removal resin (Pierce High Capacity Endotoxin Removal Spin Column, 0.5 mL, #88274; Thermo Fisher Scientific, Inc.), and finally the LPS level was measured by Limulus amebocyte lysate (Lonza) test.
Conjugation procedure
For covalent conjugation of the recombinant flagellin (FLA) to HIV-1 p24-Nef peptide based on the previous studies, the standard protocol was followed (17,34). In brief, in a separate container, 1 mM FLA was reacted with an excess amount of acetic anhydride as a carboxyl protecting group agent for at least 1 h. The protected peptide was separated by a dialysis bag to remove any impurities. In another container, 1 mM HIV1p24-Nef was activated by excess amounts of (1-ethyl-3-(3-dimethylamino) propyl carbodiimide, hydrochloride) and 10 mg N-hydroxy sulfo succinamide (Sulfo-NHS) in phosphate-buffered saline (PBS), and the reaction mixture was stirred for at least 10 min.
Afterward, the protected FLA was gently added drop wise into the activated HIV1p24-Nef container and the reaction mixture was mixed for the next 48 h. The fidelity of conjugation and the finishing time for the reaction completion were estimated by thin-layer chromatography (TLC). After dialysis, the conjugate molecules were purified by Sephadex G-75 gel filtration column (GE Healthcare, Life Sciences, Swaziland), and then verified by HPLC and TLC (data not shown).
Vaccine preparation
In this case, the HIV-1 p24-Nef-FLA conjugate and p24-Nef peptide were formulated in Montanide ISA70 adjuvant (SEPPIC, France). In brief, all of the vaccine candidates were admixed with Montanide ISA70 at the ratio of 3:7 and vigorously vortexes for 1 h to achieve a homogeneous suspension. After this, 100 μL of the mixture containing 20 μg of the vaccine candidate was used for immunization purpose.
Experimental groups and immunization schedule
The inbred BALB/c mice were divided into nine different groups as follows: Group 1: HIV-1 p24-Nef-FLA immunized s.c (n = 6) Group 2: HIV-1 p24-Nef-FLA immunized i.d (n = 6) Group 3: HIV-1 p24-Nef-FLA immunized i.m (n = 6) Group 4: HIV-1 p24-Nef immunized s.c (n = 6) Group 5: HIV-1 p24-Nef immunized i.d (n = 6) Group 6: HIV-1 p24-Nef immunized i.m (n = 6) Group 7: PBS immunized s.c (n = 5) Group 8: PBS immunized i.d (n = 5) Group 9: PBS immunized i.m (n = 5)
The regimen of all experimental groups was emulsified in Montanide ISA70 adjuvant, and experimental BALB/c mice on day 0 were immunized with 100 μL of vaccine containing 20 μg of the vaccine candidate through the mentioned routes. Also, the control groups were injected with the same route of immunization accordingly. All experimental groups were boosted on the 21st and 42nd days after immunization by the same condition.
Lymphocyte proliferation assay
Three weeks after the third immunization, the spleens of experimental mice (n = 5–6) were removed under sterile conditions and suspended in sterile cold PBS containing 2% fetal bovine serum (FBS) and pen/strep. Red blood cells were lysed with lysis buffer, and after washing the cells by centrifugation, the cell suspension was adjusted to 2 × 106 cells/mL in RPMI-1640 (Gibco, Germany) supplemented with 10% FBS, 4 mM
Phytohemagglutinin-A (5 μg/mL; Gibco), unstimulated wells, and culture medium were used as a positive control, negative controls, and blank, respectively. All experiments were done in triplicate. After 72 h of cell culture, 20 μL of BrdU (Roche, Germany) was added to each well and the plates were further incubated at 37°C for 18 h. After incubation, the plates were centrifuged at 300 g for 10 min, the supernatant was carefully aspirated and the plates were dried at 60°C for 30 min, and 200 μL of fixation/denaturation buffer was added to each well for 30 min.
The plates were aspirated and 100 μL of the anti-BrdU-horseradish peroxidase (HRP) conjugate was added and incubated for 2 h at room temperature. Afterward, the plates were washed five times with PBS, and then tetramethylbenzidine (TMB) substrate was added to the wells and incubated for 5 min in the dark at room temperature. The reaction was stopped by adding 100 μL of 2 N H2SO4. The OD450nm of each well was determined by the enzyme-linked immunosorbent assay (ELISA) plate reader (BioTek). The optical density (OD) of blank wells was subtracted from all other wells and then stimulation index (SI) was calculated according to the following formula:
ELISA of IFN-γ and IL-4 cytokines
Three weeks after the final boosting, a total of 4 × 106 spleen cells in complete RPMI-1640 were placed on each well of the 24-well plate and stimulated in vitro with 10 μg/mL of the p24-Nef fusion peptide and incubated at 37°C in 5% CO2. The other wells containing spleen cell suspension of experimental mice were cultured without antigen recall.
Three days postantigen stimulation, supernatants of stimulated and unstimulated wells were removed and the concentration of IFN-γ and IL-4 was estimated by ELISA kits (Quantikine; R&D Systems) according to the manufacturer's instruction. The concentration of each sample (pg/mL) was calculated according to the standard curve and the absolute cytokine production of each mouse was calculated by subtracting the stimulated well from the unstimulated well. Furthermore, the ratio of IFN-g/IL-4 was calculated and reported as a criterion of cytokine pattern.
Frequency of IFN-γ-producing cells by ELISPOT assay
The frequency of IFN-γ-producing splenocytes was analyzed by the ELISPOT assay (Mabtech, Stockholm, Sweden). In brief, a total of 4 × 105 spleen cells were placed on each well of the 96-well ELISPOT microplate, and in vitro stimulation was carried out with 10 μg/mL of the p24-Nef peptide. PHA was placed on some wells as a positive control of stimulated cells and wells containing unstimulated cells were used as negative controls. The plates were incubated at 37°C in 5% CO2 for 24 h. After in vitro restimulation, the plates were washed five times with washing buffer. Afterward, 100 μL of antimouse IFN-γ in PBS containing 0.5% FBS+T20 was added to the wells and incubated for 2 h at room temperature.
The plates were washed five times with washing buffer and incubated for 1 h at room temperature with 100 μL of 1:1,000 diluted streptavidin-conjugated alkaline phosphatase. After final washing, spots were developed by adding 100 μL of BCIP/NBT substrate to wells and incubated for 30 min at room temperature in dark conditions. Then, the plates were rinsed three times with distilled water and dried at 4°C.
Spots were counted using stereomicroscope (Nikon, Japan). The number of IFN-γ-producing lymphocytes was calculated by subtracting the spots from stimulated wells with an unstimulated one. The number of IFN-γ-producing cells is adjusted to 1 × 106 in the figure.
ELISA of specific total IgG antibodies and their isotypes
Specific antibodies were determined by an optimized indirect ELISA method reported previously (31). In brief, 100 μL of 10 μg/mL of the p24-Nef fusion peptide in PBS was added into 96-well ELISA Maxisorp plates (Nunc, Naperville, IL) and incubated for 24 h at 37°C. The wells were washed with PBS containing 0.05% Tween 20 (washing buffer) and blocked for 1 h at 37°C with 5% skimmed milk in PBS (blocking buffer).
Plates were washed with washing buffer and 100 μL of 1:100 diluted sera (1:100 dilution was optimized in setting up ELISA) was added to each well and incubated at 37°C for 2 h. The wells were washed five times with washing buffer and incubated for 2 h with 100 μL of 1/7,000 dilution of antimouse HRP conjugate (Sigma). The wells were washed five times and incubated for 30 min with 100 μL of TMB substrate in the dark. The reaction was stopped by adding 100 μL of 2 N H2SO4 and color density was measured at OD450nm with an ELISA plate reader (BioTek). Detection of specific IgG1 and IgG2a subclasses was performed by using goat antimouse IgG1 and IgG2a secondary antibodies (Sigma) according to the manufacturer's instruction.
Statistical analysis
All experiments were carried out in triplicate and the data are expressed as means ± standard deviation of each experiment. All statistical analyses were done by the Mann–Whitney U test. In all of the cases, p-values <0.05 were considered statistically significant.
Results
Lymphocyte proliferation assay
To evaluate the lymphocyte proliferation response, splenocytes were restimulated in vitro for 72 h with 10 μg/mL of P24-Nef peptide, and proliferation was detected utilizing the BrdU method. As shown in Figure 1, immunization of mice by vaccine candidate alone or conjugated with FLA in the different routes, that is, s.c, i.d, and i.m significantly increased lymphocyte proliferation in comparison with the control groups (p < 0.001).
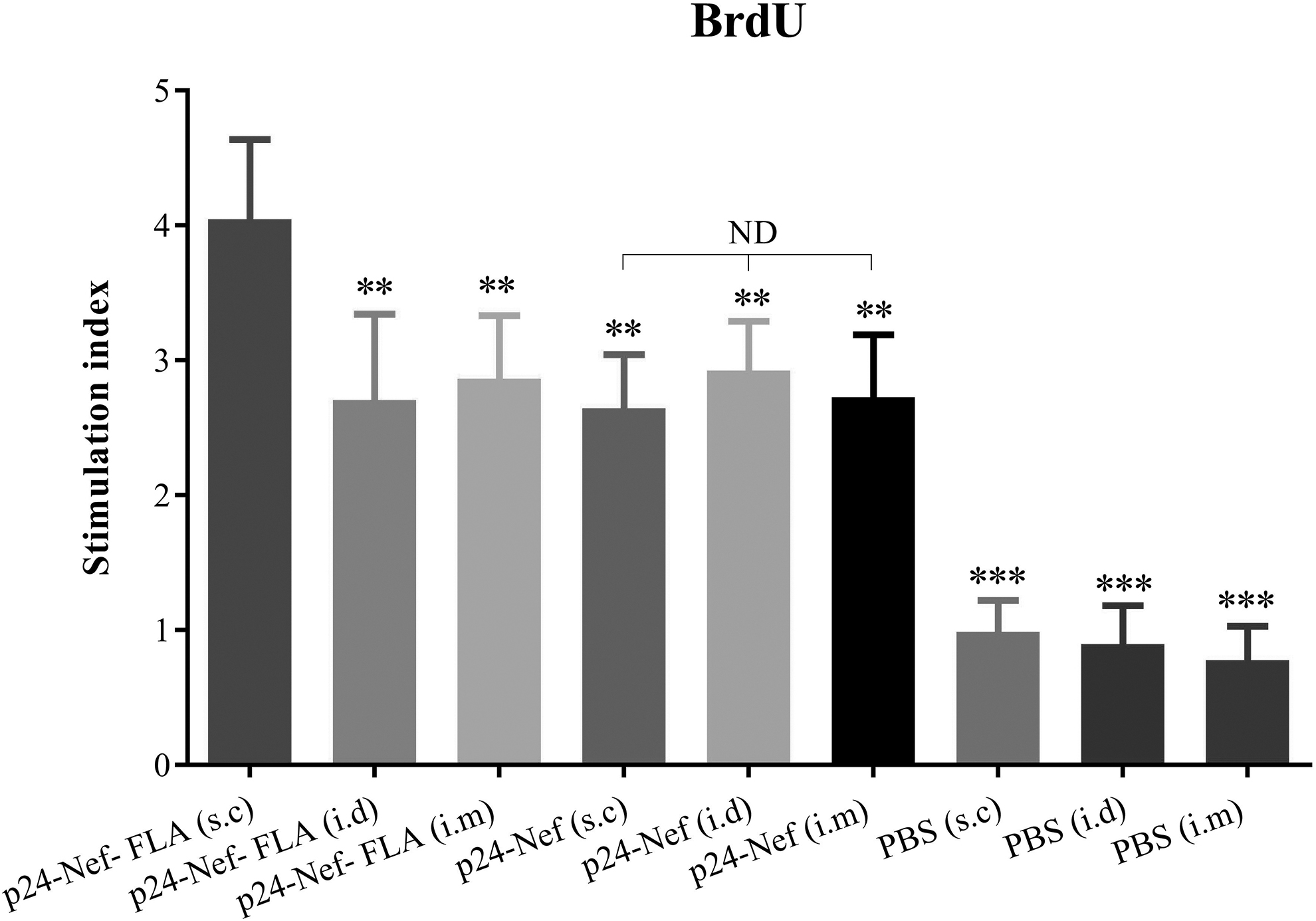
Lymphocyte proliferation response of splenic cells after antigen recall using the BrdU method. After the immunization course, splenocytes were harvested and restimulated in vitro with HIV-1 P24-Nef peptide for 72 h and lymphocyte proliferation was then quantitated using a commercially available BrdU proliferation kit as described in the Materials and Methods section. Proliferation is presented as the stimulation index of individual mice and values are represented as mean ± SD of experimental mice. Asterisks denote the groups that were statistically significant (**p < 0.01, ***p < 0.001) compared with the non-asterisk group. ND indicates not detectable differences. SD, standard deviation.
Administration of vaccine candidate formulated in the FLA protein in the s.c compared with i.m. and i.d with the same vaccine formulation and compared with vaccine candidate without FLA in the s.c, i.d, and i.m routes significantly increased lymphocyte proliferation response (p < 0.01). Administration of vaccine candidate HIV-1 p24-Nef in the different routes of immunization, that is,s.c, i.d, and i.m indicated no significant difference among the three groups (p > 0.1).
IL-4 and IFN-γ cytokines pattern
Results of IFN-γ cytokine (Fig. 2A) assay revealed that immunization with HIV-1 p24-Nef alone or conjugated with FLA molecule in s.c, i.d, and i.m. in comparison with control groups significantly increased the level of IFN-γ response (p < 0.001). Administration of vaccine candidate conjugated with FLA in the s.c route significantly increased IFN-γ release compared with i.d and i.m routes (p = 0.004). Although administration of vaccine candidate without FLA molecule in the i.d route showed the highest IFN-γ responses versus s.c and i.m. routes, no significant difference was observed between these groups (p > 0.1).
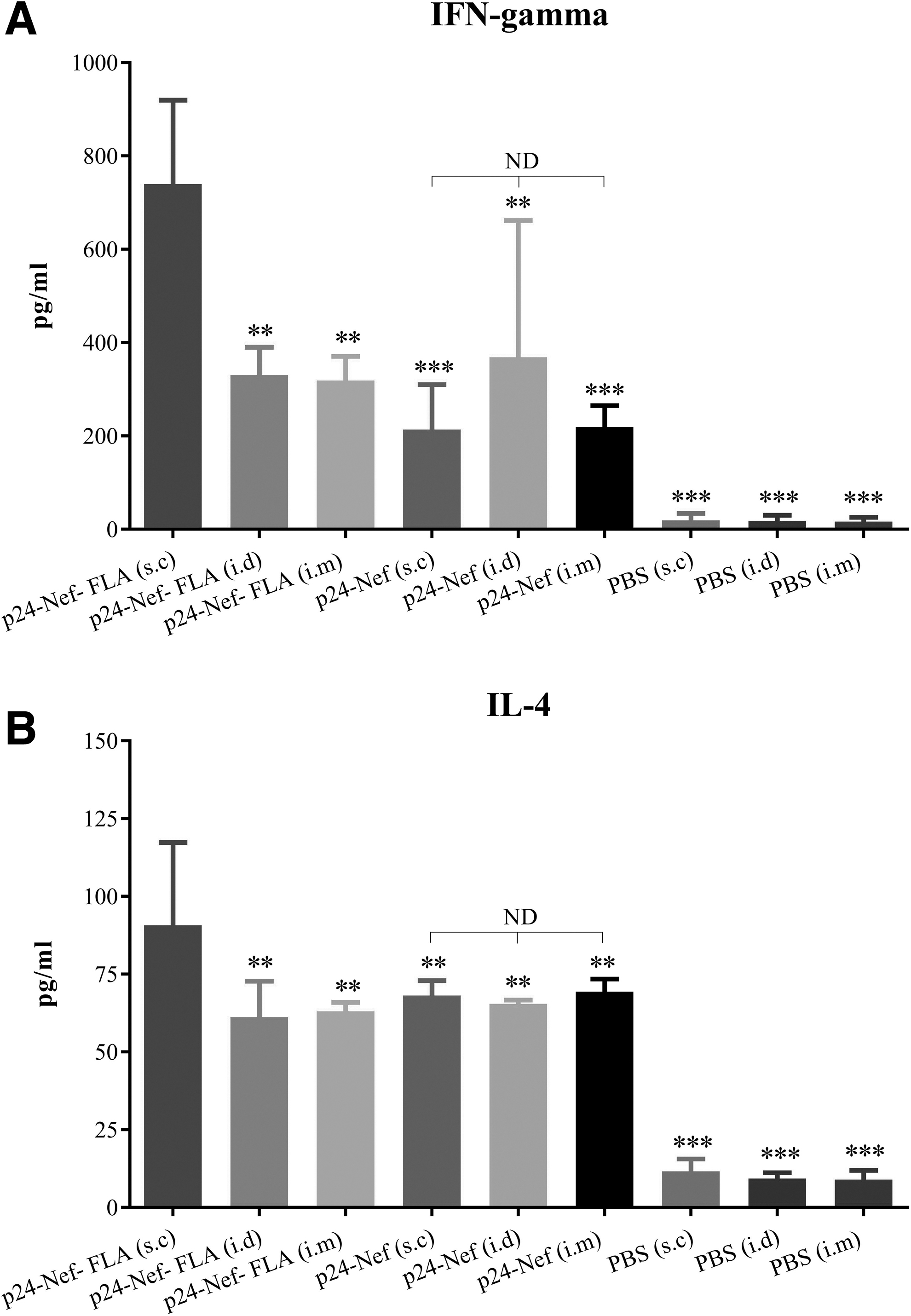
ELISA of IFN-γ
Evaluation of IL-4 cytokine response in the vaccination groups showed that injection of candidate peptide vaccine alone and conjugated with FLA molecule in the s.c, i.d, and i.m routes significantly increased the level of IL-4 responses compared with the control groups (p < 0.001) (Fig. 2B). Injection of vaccine candidate conjugated to FLA in the s.c route significantly increased IL-4 response compared with i.d and i.m routes with the same formulation of vaccine (p = 0.004).
Immunization with HIV-1 p24-Nef vaccine candidate in the different routes of immunization, that is, s.c, i.d, and i.m showed no significant difference in the induction of IL-4 cytokine response (p > 0.3).
IFN-γ cytokine ELISPOT assay
The frequency of IFN-γ-secreting lymphocytes was evaluated using the ELISPOT assay. Administration of vaccine candidate alone or conjugated to FLA significantly augmented number of IFN-γ-secreting lymphocytes as compared with the control groups (p < 0.05). Also, immunization of mice with vaccine candidate conjugated to FLA in the s.c route significantly increased IFN-γ-producing lymphocytes compared with i.d and i.m routes with the same vaccine formulation (p < 0.009). Also, immunization with HIV-1 p24-Nef in the s.c, i.d, and i.m. routes indicated no significant difference among the three groups (p > 0.1) (Fig. 3).
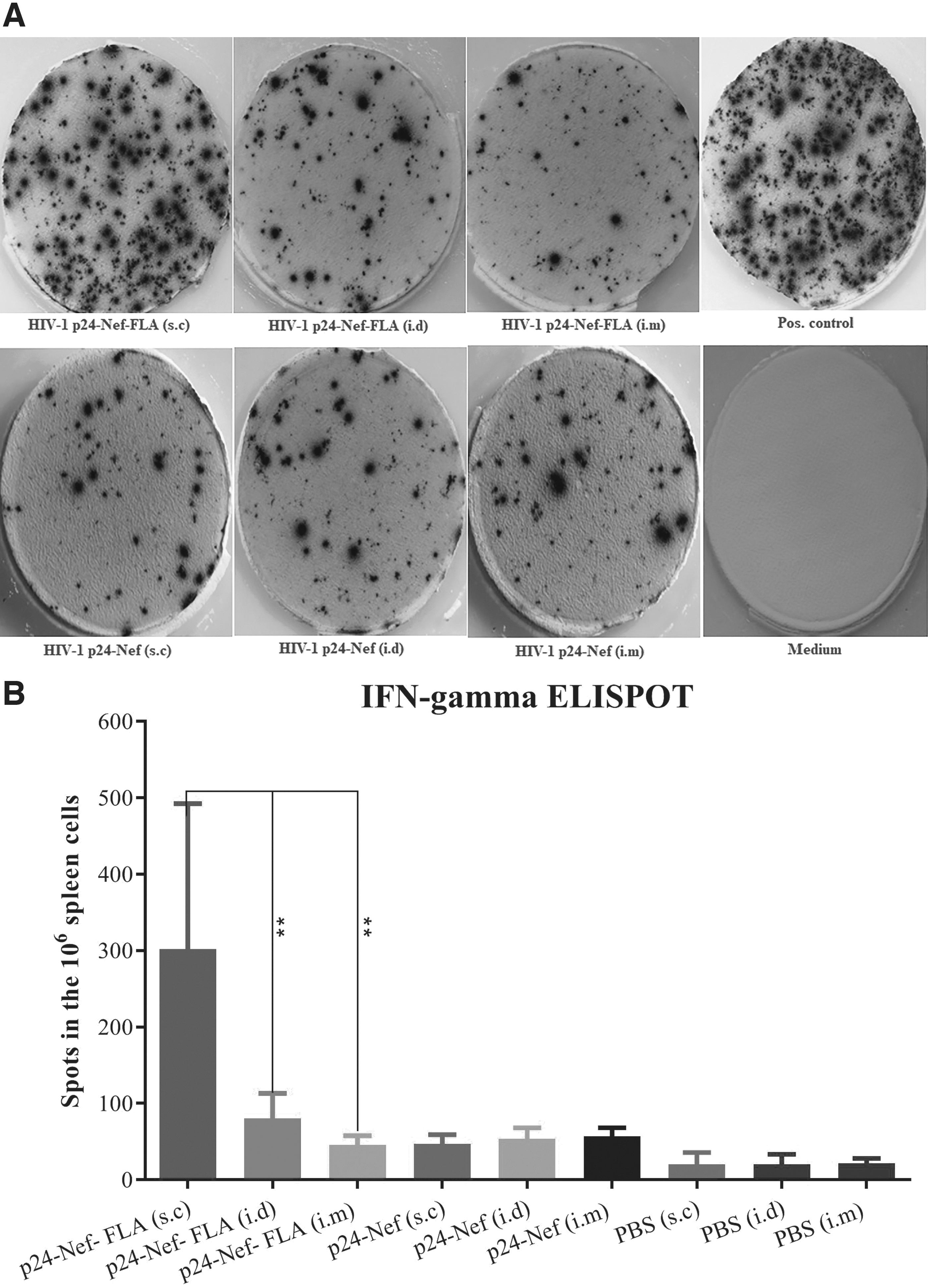
Quantification of the frequency of IFN-γ-producing cells in ELISPOT. Enumeration of IFN-γ-excreting splenocytes in response to mitogen and peptide stimulation. Splenocytes were isolated from the spleens of experimental mice. Splenocytes were either treated with medium (no stimulus control), the mitogen PHA, or HIV-1 P24-Nef peptide for 24 h and enumerated using ELISPOT assays specific for IFN-γ. Representative wells are shown
Specific total IgG response
Assessment of specific IgG response shows that immunization of mice with candidate vaccines with or without flagellin induced a high level of specific IgG antibody responses in the s.c, i.d, and i.m. routes, which show significant differences with control groups (p < 0.006) (Fig. 4). The level of specific total IgG in mice immunized with the conjugated vaccine to FLA in the i.m. route showed a significant difference with immunized groups with the conjugated vaccines in the s.c and i.d routes (p < 0.006). Immunization with the mere vaccine in the s.c, i.d, and i.m. routes indicated no significant differences among the three groups in the induction of specific total IgG responses (p > 0.05).
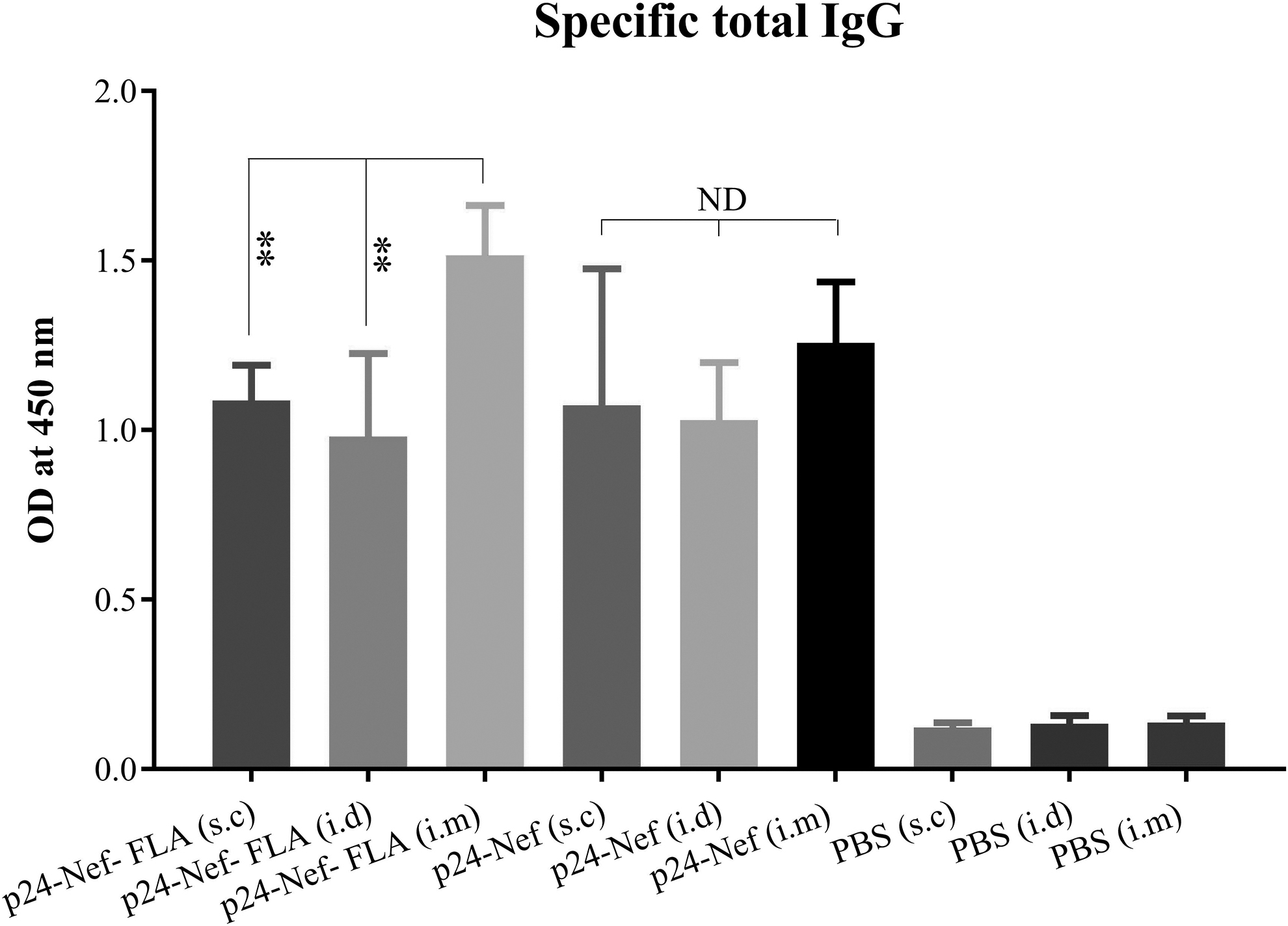
Specific total IgG antibody responses in the experimental groups. Experimental mice were immunized with vaccines with different formulations and a specific total IgG antibody was evaluated with an indirect ELISA method. Data are shown as the mean ± SD. Asterisks represent the statistically significant groups and ND indicates not detectable differences (p < 0.05).
Specific IgG1 and IgG2a immune responses
Results of specific IgG1 and IgG2a isotyping showed that immunization with conjugated and mere vaccine increased IgG1 and IgG2a isotypes level in all three routes of injection compared with the control groups (p < 0.01). The results of specific IgG1 responses showed significant differences in the groups that received the vaccine conjugated to FLA in the i.m route compared with the groups with the conjugated vaccine in the s.c and i.d routes (p < 0.02). Also, immunization with a mere p24-Nef vaccine candidate in all three routes of immunization does not show significant differences in the induction of IgG1 responses among each other (p > 0.05) (Fig. 5A).
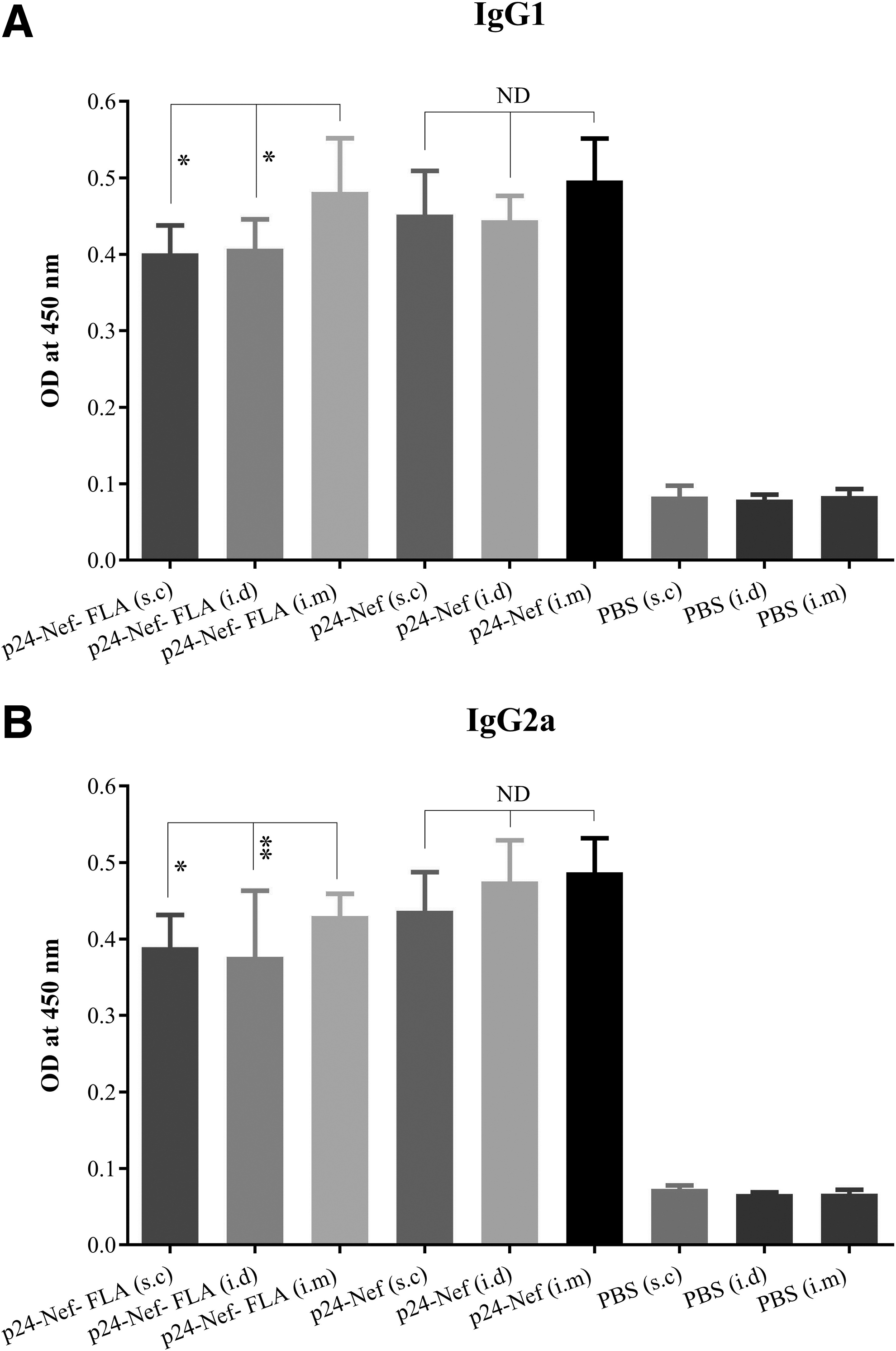
Detection of specific IgG1
Administration of the conjugated vaccine with FLA in the i.m. route showed a significant increase of specific IgG2a level versus i.d and s.c routes with the same vaccine formulation (p = 0.007 and p = 0.03, respectively). Vaccination with HIV-1 p24-Nef vaccine candidate in the s.c, i.d, and i.m. routes revealed no significant difference among the three groups in the induction of IgG2a isotype response (p > 0.05) (Fig. 5B).
Discussion
Improvement of vaccine efficacy is a hot topic in vaccinology (33). Immune modulators and adjuvants are important components of modern vaccines and are needed to improve the vaccine's immunogenicity. One of the problems for subunit-based vaccines is that when administered alone (without adjuvant), they do not properly stimulate protective immunity against infectious agents. It has been demonstrated that activation of TLRs can result in the stimulation of APCs and enhanced B and T cells activation. Nowadays, TLR agonists are considered promising vaccine adjuvant candidates. The use of flagellin adjuvant as the TLR-5 agonist beside vaccine candidate by stimulation of innate immune responses can be a useful strategy to increase the immunogenicity and also the efficacy of vaccine candidate (32).
Besides vaccine formulation, the optimum route of immunization can probably improve vaccine efficacy (22). Herein, we hypothesized that the modification in the vaccine formulation may change its effective route of immunization. So, here two formulations of HIV-1 p24-Nef vaccines (vaccine conjugated to FLA molecule and mere HIV-1 p24-Nef vaccine) were evaluated in the s.c, i.d, and i.m routes of immunization to assess our hypothesis.
Results from lymphocyte proliferation assay as a criterion of cellular immune response function (10) showed that immunization with HIV-1 p24-Nef formulation had an equal induced effect on the lymphocyte proliferation in the s.c, i.d, and i.m routes, but immunization with HIV-1 p24-Nef-FLA in the s.c route showed a significant increase in the lymphocyte proliferation in comparison with i.m and i.d routes with similar vaccine formulation. This finding shows that with a change in the vaccine formulation and route of immunization, promising results could be obtained by stimulating the lymphocyte proliferation, and thus, an effective cellular immune response will be established.
A major difference between T cell proliferation rates in different routes of immunization appears to lie in the range of antigens recognized by responding T cells. It has been proven that repertoire of T cells involved in the immune response against an immunogen may vary in each route of immunization. In fact, T cells from one route of immunization may recognize a relatively narrow range of antigens, whereas in another route of immunization, a more wide range of antigens may be recognized and more repertoire of T cells may be activated (30). In addition, the different level of lymphocyte proliferation rate in the different routes of immunization may be due to the nature of APCs in these different locations that are functionally distinct and have different prime activities and thereby effect on immune response patterns (16).
Administration of vaccine candidate conjugated with FLA in the s.c route significantly increased IFN-γ release compared with i.d and i.m routes. Whereas vaccine candidate without FLA molecule in the i.d route showed the highest IFN-γ responses versus s.c and i.m. routes but was not statistically significant. Interestingly, results of the IFN-γ ELISPOT assay demonstrated that the frequency of IFN-γ-producing lymphocytes of vaccine candidate conjugated to FLA in the s.c route significantly increased compared with i.d and i.m routes, whereas immunization with HIV-1 p24-Nef in the s.c, i.d, and i.m. routes indicated no significant differences. The results obtained here were in concert with the results of the lymphocyte proliferation test so that the conjugation of FLA to HIV-1 vaccine candidate was versatile in the induction of the highest Th1 response through the s.c route.
Because of 24 h in vitro stimulation in ELISPOT assay, the responding cells considered as effector memory T cells (24) that reflect immunization with FLA-formulated vaccine in the s.c route developed the highest number of effector memory T cells that produce IFN-γ cytokine.
These findings show that the route of immunization and vaccine formulation not only affect the type of immune responses but also determine the frequency of memory T cell formation.
Our finding is parallel to various studies that were performed before (7). The previous study showed that the route of immunization and the location of encountering naive T cells to the antigen determine the fate of memory T cells, and in each route of immunization a distinct pattern of memory T cells and also the type of immune responses would be developed (25).
Injection of HIV-1 p24-Nef-FLA in the s.c route significantly increased IL-4 cytokine response compared with i.d and i.m routes with the same formulation, whereas immunization with HIV-1 p24-Nef in the different routes of injection showed no significant difference in the induction of IL-4 cytokine response. This finding shows that the use of FLA as an adjuvant in the vaccine formulation changes its effective route in the induction of Th2 cytokine pattern, whereas mere vaccine shows the same potency in all of the immunization routes to induce Th2 response. Our previous study confirmed that the FLA is a strong inducer for antibody-mediated immune response, and a single dose of this immunogen could cause a protective immunity against P. aeruginosa in the burn model of infection (9).
Results from total IgG response in the experimental groups showed that the i.m route of the conjugated form of the HIV-1 p24-Nef significantly raised specific antibody response compared with i.d and s.c routes with the same formulation, whereas immunization of mice with HIV-1 p24-Nef in different routes showed no significant difference in terms of inducing a humoral immune response. This finding demonstrated that FLA as an adjuvant could increase Th2-dependent immune response against vaccine candidate as reported by various studies (5,20,27).
In addition, injection of conjugated vaccine candidate following the i.m route could have the highest boosting effect on the humoral immune response. Higher level of antibody response depends on the function of Tfh responses, its interaction with local APCs, and also vaccine stability after deposition at the injection site (23,28,29). Regardless of the role of local immune cells in the improvement of humoral immune responses, deep muscle immunization, due to keeping intact the antigen and more time releasing property, resulted in more exposure time for immune cells that may be accompanied with higher immune responses (15,18).
Results of specific IgG1 and IgG2a antibody responses revealed that immunization with HIV-1 p24-Nef-FLA in the i.m route significantly raised IgG1 and IgG2a responses compared with the i.d and s.c routes with the same formulation, whereas no significant difference was observed in the induction of IgG1 and IgG2a responses after immunization of mice with HIV-1 p24-Nef through different routes. Since the muscle tissue contains very few antigen-presenting resident cells (3) and i.m. vaccination (compared with other routes) can lead to further involvement of lymph nodes and spleen, it could be concluded that a higher percentage of antigens will be exposed to B lymphocytes and, therefore, a high level of induction of the humoral immune response will be established (15).
Conclusion
In summary, change in vaccine formulation and delivery strategies for eliciting optimum immune response is important for developing effective prophylactic vaccines against infectious diseases, which are considered as global concerns for infectious disease. In this study, we found that the change in vaccine formulation through conjugation of the vaccine candidate to the flagellin protein could lead to changes in the immunological characteristics. We concluded that any change in the vaccine formulation could effect on the effective route of immunization, which is very important in vaccine optimization. The changes might alter the vaccine efficacy, which could eventually lead to access to the most potent vaccine.
Footnotes
Acknowledgment
The authors thank Mrs. Zahra Shahosseini for expert technical assistance.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by a grant (Grant No. 595) from the Pasteur Institute of Iran and a grant from Shahrekord University of Medical Sciences (Grant No. 925).