
Abstract
Declined immune response is the main cause of decreased potency of the influenza vaccine in the elderly, regardless of virus mutations. Herein, we hypothesized that the addition of α-tocopherol to the influenza vaccine formulation might increase vaccine potency and efficacy. Hemagglutinin of the H1N1 virus was formulated in Alum and α-tocopherol, and then aged (16–20-month-old) and young (6–8-week-old) mice were immunized subcutaneously two times with 2-week intervals with 5 μg of different vaccine formulations. Two weeks after the final boosting, IFN-γ and IL-4 cytokines were assessed by using ELISA. Humoral immune responses were assessed by hemagglutination inhibition (HI). In addition, vaccine efficacy was determined by intranasal viral challenge of mice using mouse-adapted H1N1 virus. Our results showed that the new vaccine formulation improved IFN-γ and IL-4 responses in the experimental mice. However, the increase was evident mainly in the aged group and, to some extent, in the young group. Results from the HI assay showed that α-tocopherol in the vaccine formulation could increase HI activity in both young and aged mice. Furthermore, α-tocopherol, as an adjuvant, increased the protectivity of the influenza vaccine in both aged and young groups through the decreased lung viral load and increased survival rate of the experimental mice. In conclusion, it seems that α-tocopherol can not only be used as an appropriate adjuvant for aged people, but also empower old and worn out cells to increase the effectiveness of the vaccine in the elderly.
Introduction
Influenza is a life-threatening viral infection negatively affecting the respiratory tract (13,41). The highest influenza-related mortality happens in the elderly (15). According to the CDC report, ∼70–85% and 50–70% of the regular influenza-related deaths and hospitalizations happen in individuals who were 65 years old, respectively. Therefore, influenza is regularly very genuine for individuals aged 65 and more years. Despite the presence of different types of influenza, the new types can spread globally and cause influenza pandemics with increased case numbers (13,43). The influenza virus appears with symptoms such as cough, sore throat, fever, body aches or diarrhea, and vomiting (45). The symptoms may not be detected first before testing. The virus causes million severe cases, a great number of deaths, and subsequent economic losses each year.
Vaccination is a rational approach to combat influenza infection (27). Although immunization and vaccination are the foremost successful intercession, jumbles between circulating infections and immunization strains decrease antibody adequacy (37,45). Higher doses of the influenza antigen may increase resistant reaction (43). Although a wide variety of vaccines with different efficacies have been made so far, none of them have been able to completely prevent the spread of the disease, especially in aged people. Therefore, there is an urgent need for vaccine optimization to achieve a suitable protection rate, especially for elderly.
Vitamin E is a lipid solvent vitamin comprising of eight normal isoforms, specifically α, β, δ, γ isoforms of tocopherol and α, β, δ, γ isoforms of tocotrienol (3,10). α-Tocopherol, as a bioactive form of vitamin E, is a vital antioxidant (18,20), which is used in a variety of cases, such as cancer prevention and therapy, treatment of Alzheimer, and application effects on skin (11). Recent studies have uncovered cellular and systemic mechanisms, showing that vitamin E might be appropriate treatment strategy for a diverse population of aged individuals (40). Animal and human studies exhibited that higher levels of vitamin E, as compared to the presently prescribed levels, are vital to balance immune and inflammatory responses in aged people (31).
In this study, we hypothesized that the addition of α-tocopherol to the influenza vaccine formulation may reinforce the responding cells of aged mice. In turn, this may result in an increase in the immune response to the vaccine. For this purpose, hemagglutinin of the H1N1 virus was formulated in Alum and α-tocopherol, followed by subcutaneous immunization of aged (16–20-month-old) and young (6–8-week-old) mice two times with the newly formulated vaccine. Lastly, the potency and efficacy of the newly developed vaccine were assessed in the young and aged mice.
Materials and Methods
Animals
Female Balb/c mice, including aged (16–20-month old) and young (6–8-week old) groups, were purchased from Pasteur Institute of Iran (Karaj, Iran). Mice were kept for 1 week before the experiment, given access to food and water ad libitum, and maintained in a light/dark cycle with lights on from 6:00 to 18:00 h. All procedures, before and during the experimental period and animal handling, were performed in agreement with the standard protocols of the Pasteur Institute of Iran.
Vaccine formulation
Purified hemagglutinin of the H1N1 influenza (Influenza Antigen A/California/7/2009 [H1N1]-cell derived) was purchased from The National Institute for Biological Standards and Control (NIBSC, United Kingdom). Each vial contained 50 μg of purified hemagglutinin, which was used for the vaccine formulation and immunoassay. Molecular grade aluminum hydroxide was provided from Pasteur Institute of Iran (Karaj, Iran). α-Tocopherol was purchased from Sigma Company.
For vaccine formulation, Alum (200 μg as one dose) was admixed with hemagglutinin (5 μg for each dose) at PH = 6 according to their isoelectric points. The mixture was vortexed for 1 h at room temperature (RT), and allowed for absorption of the antigen on the Alum adjuvant. Then, α-tocopherol was dissolved in ethanol alcohol and added to the vaccine formulation at three doses, including 1, 5, and 10 mg, for each dose of the vaccine under continuous vortexing. After achieving a homogenized suspension, the reaction was stopped and the vaccine was used for immunization. In this formulation, a dose of vaccine contained 5 μg of hemagglutinin, 200 μg of Alum, and α-tocopherol at the desired dose (1, 5, and/or 10 mg/dose).
Experimental groups and immunization
In this study, young and aged mice were used as experimental animals. The young (Table 1) and aged (Table 2) mice were divided into 8 groups, consisting of 15 mice in each group. The groups of experimental young and aged mice were immunized as below:
Experimental 6–8-Week-Old Mice as Young Cohort
HA, hemagglutination assay.
Experimental 16–20-Month-Old Mice as Aged Mice Cohort
Group-1: Mice immunized with 5 μg of hemagglutinin-Alum.
Group-2: Mice immunized with 5 μg of hemagglutinin-Alum-α-tocopherol-1 mg.
Group-3: Mice immunized with 5 μg of hemagglutinin-Alum-α-tocopherol-5 mg.
Group-4: Mice immunized with 5 μg of hemagglutinin-Alum-α-tocopherol-10 mg.
Group-5: Mice immunized with Alum-α-tocopherol-1 mg as a control group.
Group-6: Mice immunized with Alum-α-tocopherol-5 mg as a control group.
Group-7: Mice immunized with Alum-α-tocopherol-10 mg as a control group.
Group-8: Mice immunized with PBS as a control group.
All mice were vaccinated subcutaneously two times on days 0 and 14. All vaccine volumes were 200 μL and each mouse received 5 μg of hemagglutinin, as an immunogen, in the vaccine formulation.
IFN-γ and IL-4 cytokine ELISA
ELISA was used to assess the levels of IFN-γ and IL-4 cytokines in the immunized mice. For this purpose, blood samples were taken from the mice 16–18 h after the final immunization. Afterward, the mouse sera were harvested using centrifugation and then stored at −70°C until cytokine assay. Quantitative IFN-γ and IL-4 cytokine assessment were carried out with mouse IFN-γ and IL-4 commercial ELISA kits (Mabtech, Sweden) concurring to the manufacturer's protocol. The amount of each cytokine was calculated concurring to its standard curve and presented as pg/mL for each individual mouse.
Viral propagation and titration with hemagglutination assay
The Madin-Darby canine kidney (MDCK) cell line was cultured in a minimum essential medium (Sigma) supplemented with 10% fetal bovine serum,
Hemagglutination inhibition
Two weeks after final immunization, sera from the mice were collected to determine the potency of neutralizing antibodies through masking the HA antigen in the hemagglutination inhibition (HI) assay.
Experimental sera were inactivated by incubation at 56°C for 30 min. Then, nonspecific inhibition of hemagglutination was abrogated by adding 225 μL of a 25% kaolin suspension to inactivated sera. The samples were vortexed and incubated for 20 min at RT. After centrifugation for 2 min at 6,000 g, 50 μL of the supernatants were collected and serially diluted in PBS in U-bottom 96-well plates (Greiner, Germany). Then, the PR8 virus (4 U) was added to each well and the plates were incubated at RT for 1 h. Afterward, 0.5% chicken red blood cells were added to all wells and incubated for 30 min at RT. The HI titer for each mouse was reported as the reciprocal of the highest serum dilution that completely inhibited hemagglutination (22).
Experimental challenge
The viral titer needed to kill 50% of inoculated mice (MLD50) was calculated through anesthetizing 6 to 8-week-old BALB/c mice (5 mice/group) with 50 μL of the serial 10-fold diluted influenza virus. Mice were weighed and checked daily for 14 days postinfection and regarded as dead when they lost 25% of their body weight. MLD50 titers were then calculated using the karber method. For the challenge study, the mice were gently anesthetized 3 weeks after the final immunization with a mixture of Ketamine/Xylazine and challenged intranasally with 100 MLD50 of PR8 [A/Puerto Rico/8/34 (H1N1)] influenza viruses (22).
Viral load
Four days after viral challenge, the mice were dislocated and their lungs were removed and homogenized mechanically in Dulbecco's modified Eagle's medium containing 0.1% bovine serum albumin. The samples were filtered and served as viral challenge stocks. MDCK cells were cultured in 96-well plates and the diluted homogenized lung samples were (10-fold serially) added to the culture. The plates were incubated 48 h at 37°C in an atmosphere of 5% CO2. Once the cytopathic effect was detected, the virus titer of each sample was calculated as 50% tissue culture infection dose (TCID50) by the Reed–Muench method (2).
Survival rate
The mortality rate of mice was recorded daily after the viral challenge. The mice were monitored for 14 days after the viral challenge.
Statistical analysis
The experiments of this project were performed in duplicate or triplicate. Graph Pad prism version 6 software was used for statistical analysis of raw data. One-way ANOVA and Tukey test with 95% confidence interval were used for statistical comparison between the experimental groups. p-Value less than p < 0.05 was considered as significant difference. Kaplan–Meier and log rank test were used for survival analysis.
Results
IFN-γ cytokine response in aged mice
Assessment of IFN-γ, as the major Th1 pattern cytokine, showed that the vaccine formulated in α-tocopherol leads to increased IFN-γ, as compared with mere vaccine (Fig. 1). In fact, the addition of α-tocopherol to the vaccine formulation, at all three concentrations of 1, 5, and 10 mg, resulted in a significant increase in the IFN-γ level, as compared with the mere vaccine (p = 0.0234, p = 0.0144 and p = 0.0085, respectively). The comparison of IFN-γ responses, among the three concentrations, showed that the vaccine formulated in 1 mg of α-tocopherol leads to an increased level of 2.5% to 5%, when compared with those formulated in 5 and 10 mg.
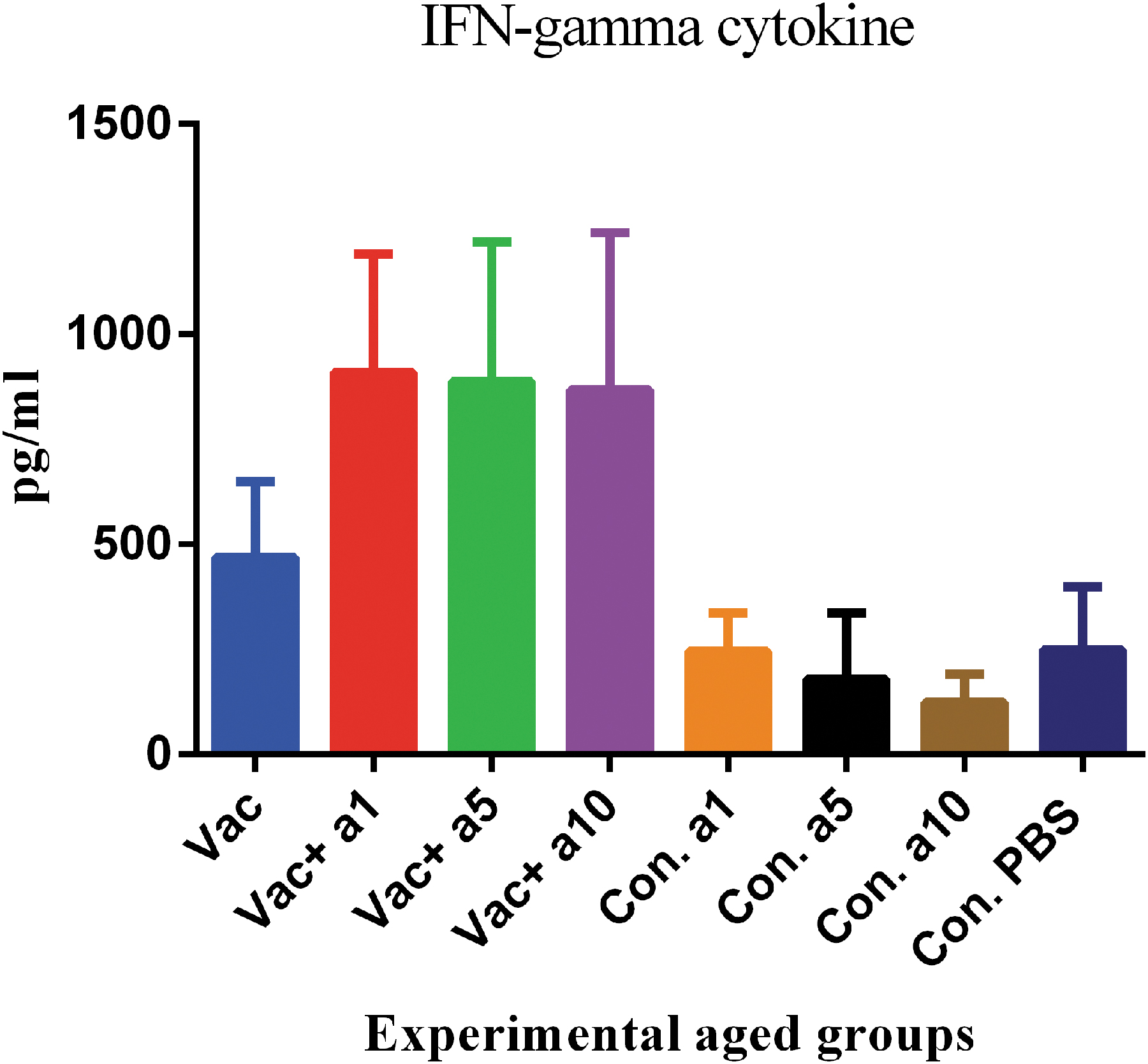
IFN-γ cytokine response of aged mice after two shots of vaccine. The addition of α-tocopherol to the vaccine formulation at the doses of 1, 5, and 10 mg caused a significant increase in the IFN-γ levels compared with the mere vaccine (p = 0.0234, p = 0.0144, and p = 0.0085, respectively).
IFN-γ cytokine response in the young mice
Assessment of IFN-γ levels in the young mice showed that the vaccine formulated in α-tocopherol could increase the IFN-γ level, as compared with the mere vaccine; however, this was not statistically significant. In fact, the addition of α-tocopherol to the vaccine formulation, at all three concentrations of 1, 5, and 10 mg, caused a slight increase in the IFN-γ levels, as compared with the mere vaccine, which did not show a statistically significant difference with the vaccine group alone (p = 0.9502, p = 0.8392, and p = 0.9939, respectively). In addition, the slight increase in the IFN-γ cytokine, in the new formulation with concentrations of 1, 5, and 10 mg of α-tocopherol, was 5.8%, 10.2%, and 8.2%, when compared with the mere vaccine, respectively (Fig. 2).

IFN-γ cytokine response in the young mice after vaccination. Influenza vaccine formulated with α-tocopherol increased the IFN-γ cytokine level as compared with the mere vaccine, but was not statistically significant. In fact, α-tocopherol at the doses of 1, 5, and 10 mg caused a slight increase in IFN-γ cytokine response as compared with the mere vaccine (p = 0.9502, p = 0.8392, and p = 0.9939, respectively).
IL-4 cytokine response in the aged mice
Examination of IL-4, as the main cytokine characteristic of the Th2 pattern, in the aged mice showed that the vaccine formulated in α-tocopherol, at concentrations of 1, 5, and 10 mg, causes a nonsignificant increase in cytokine levels, as compared with the mere vaccine (p = 0.9825, p = 0.9990, and p = 0.5619, respectively). Additionally, the increased level of IL-4 in the new formulation with concentrations of 1, 5, and 10 mg of α-tocopherol was 41%, 17.8%, and 25.56%, when compared with the mere vaccine, respectively (Fig. 3).

IL-4 as a Th2 response in the aged mice. The influenza vaccine formulated with α-tocopherol at the doses of 1, 5, and 10 mg caused a nonsignificant increase (41%, 17.8%, and 25.56%, respectively) as compared with the mere vaccine (p = 0.9825, p = 0.9990, and p = 0.5619, respectively).
IL-4 cytokine response in the young mice
Evaluation of IL-4, as a cytokine of the Th2 pattern, in the young mice indicated that the vaccine formulated in α-tocopherol at concentrations of 1, 5, and 10 mg showed no significant changes, as compared with the mere vaccine (p = 0.9411, p = 0.9995, and p = 0.9999, respectively) (Fig. 4).
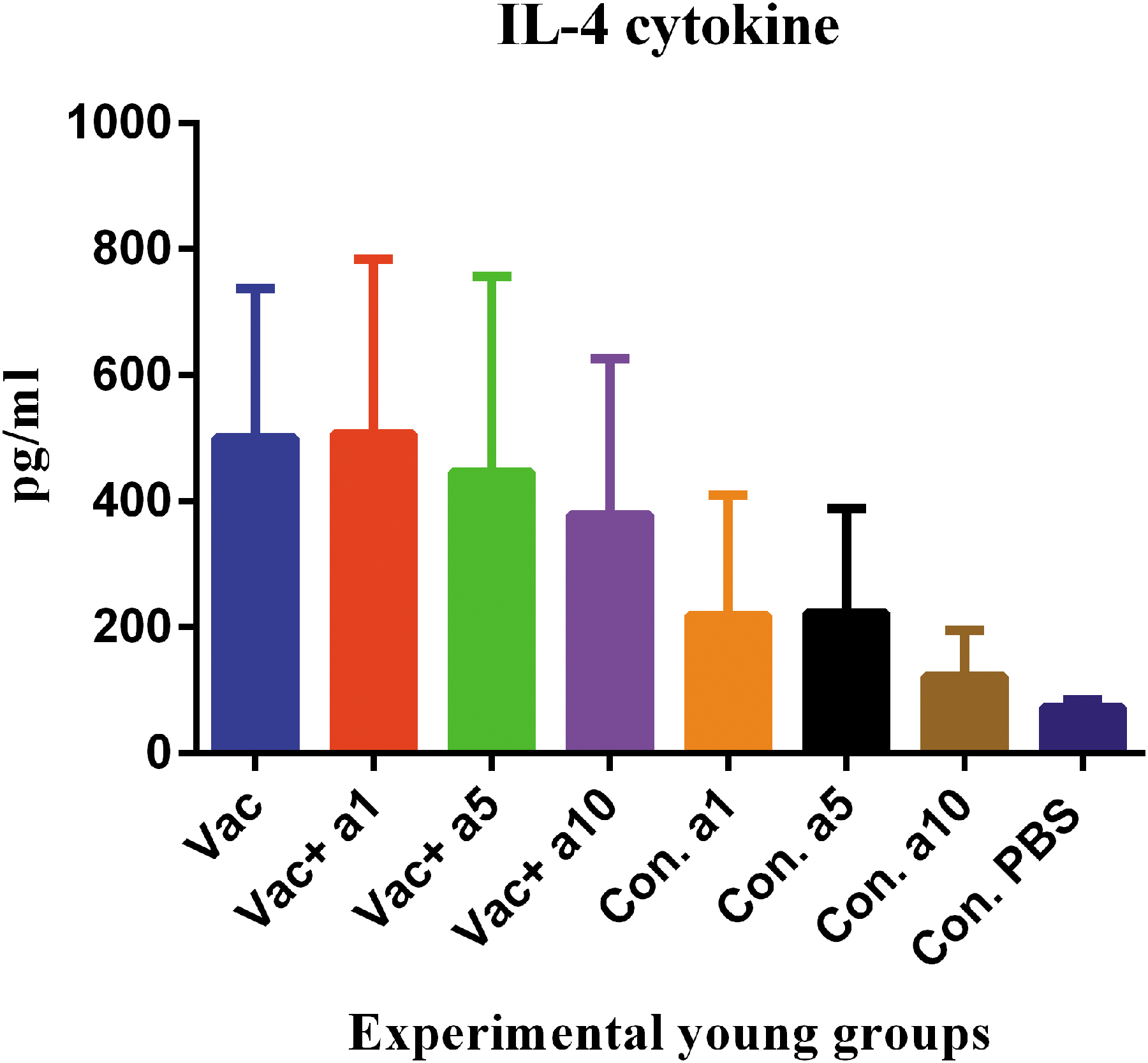
IL-4 response in the young mice. Influenza vaccine formulated with α-tocopherol at the doses of 1, 5, and 10 mg showed no significant changes, as compared with the mere vaccine (p = 0.9411, p = 0.9995, and p = 0.9999, respectively).
HI potency of aged mice
The antibody response was evaluated using its potency in the virus neutralization and inhibition of hemagglutination. Functional evaluation of the antibodies produced after vaccination showed that the vaccine, formulated in α-tocopherol, at concentrations of 1, 5, and 10 mg, significantly increased its ability to inhibit hemagglutination in these groups, as compared with the mere vaccine (p = 0.0001, p = 0.0001, and p = 0.0649, respectively). Furthermore, it seems that the vaccine formulated in 1 and 5 mg of α-tocopherol was more effective than that formulated in 10 mg in inducing this type of response (p = 0.0397 and p = 0.0302, respectively) (Fig. 5).
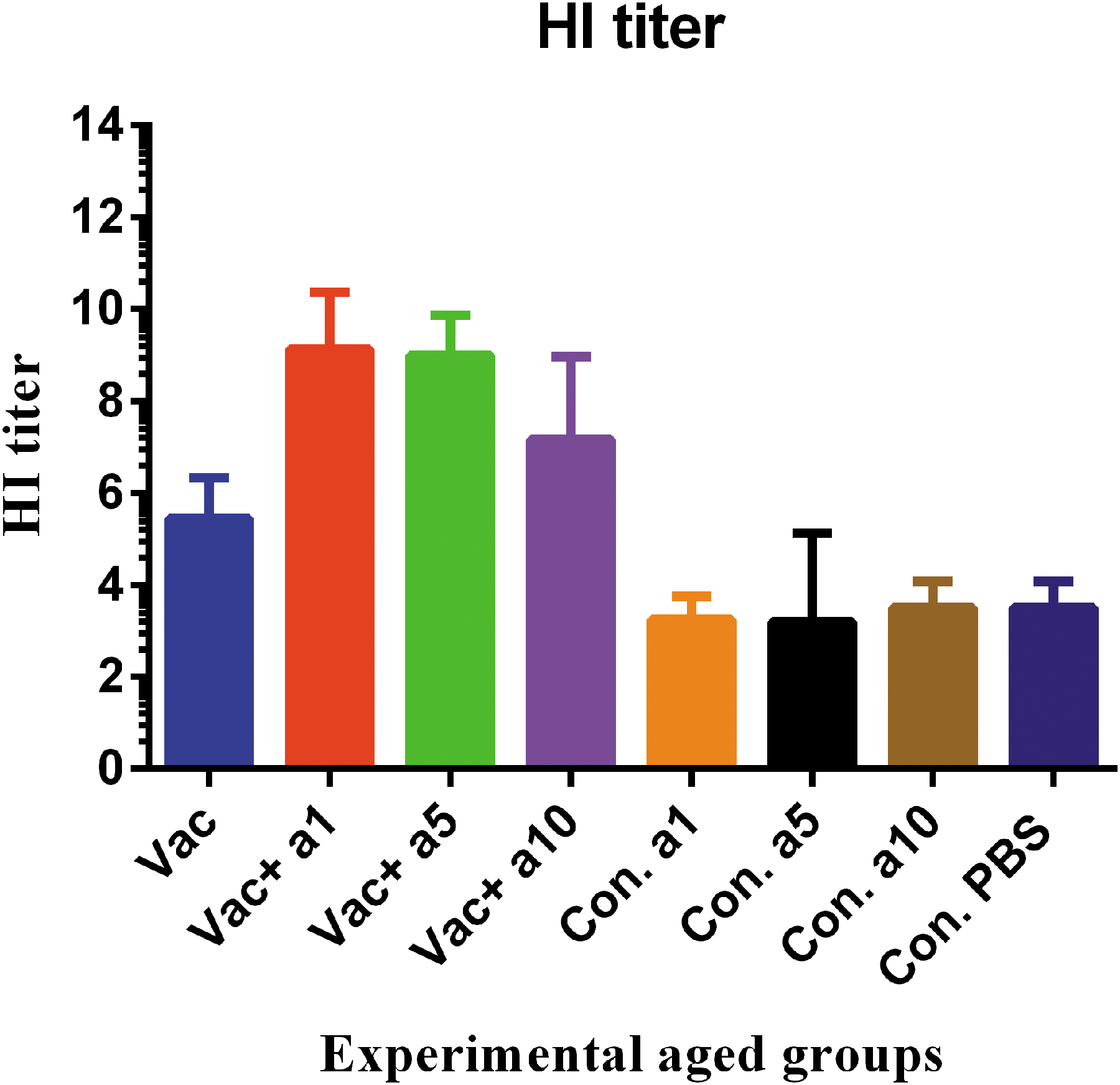
HI potency of sera from the aged mice. The function of antibodies produced in the old mice after vaccination in the hemagglutination inhibiting process showed that the influenza vaccine formulated with α-tocopherol at the doses of 1, 5, and 10 mg significantly increased the HI activity in these groups as compared with the mere vaccine (p = 0.0001, p = 0.0001, and p = 0.0649, respectively). HI, hemagglutination inhibition.
HI of young mice
Examination of antibody responses in young mice using the HI test showed that the vaccine formulated in α-tocopherol at concentrations of 1, 5, and 10 mg increased the ability to HI, as compared with the mere vaccine, but only at a concentration of 1 mg showed a significant difference (p = 0.0083, p = 0.7315, and p = 0.9204, respectively). In addition, it appears that the vaccine formulated in α-tocopherol at the concentration of 1 mg is more effective in inducing this response, as compared with that formulated in other concentrations; however, this exhibited no statistically significant difference (p = 0.6448 and p = 0.2315, respectively) (Fig. 6).
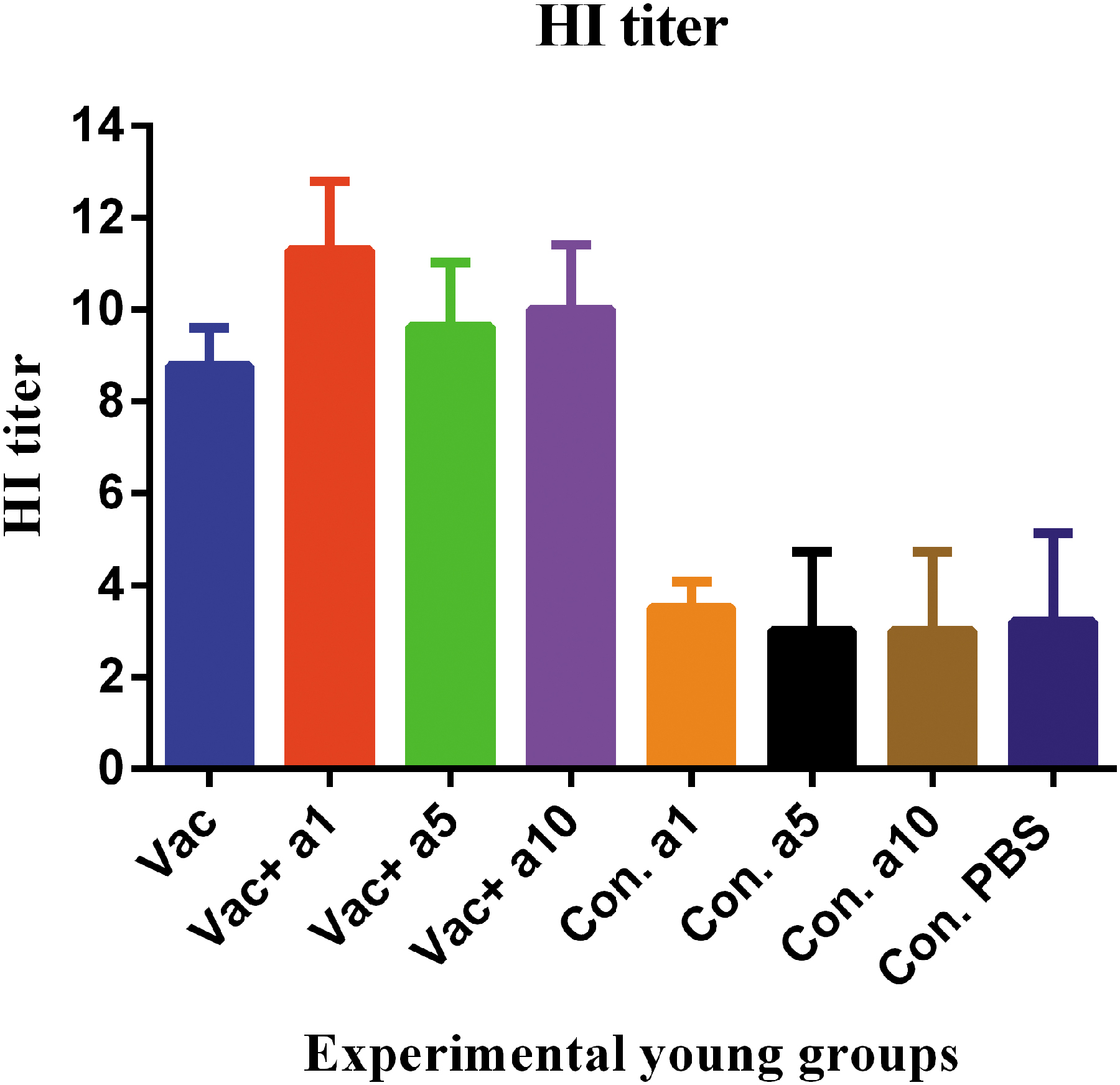
HI activities of experimental young mouse sera. Results showed that the influenza vaccine formulated with α-tocopherol at the doses of 1, 5, and 10 mg increased the ability to HI as compared with the mere vaccine but only at the dose of 1 mg was highly effective, showing a significant difference versus the mere vaccine (p = 0.0083, p = 0.7315, and p = 0.9204, respectively).
Lung viral load in the aged mice
To evaluate the potency of different vaccine formulations in the control of viral infection, the viral load was evaluated in the lung tissue from the challenged mice. Examination of viral load showed that the vaccine formulated in α-tocopherol at concentrations of 1, 5, and 10 mg leads to decreased viral loads in these groups, as compared with the mere vaccine (p = 0.8017, p = 0.9103, and p = 0.0187, respectively); however, this effect was statistically significant only at a concentration of 10 mg of α-tocopherol (Fig. 7). In addition, it appears that the vaccine formulated in α-tocopherol at a concentration of 10 mg was more effective in controlling the viral load than that formulated at the doses of 1 and 5 mg, but this decreased viral load was not statistically significant (p = 0.2849 and p = 0.4196, respectively).

Lung viral load in the aged mice after intranasal virus challenge with mouse adapted H1N1 virus. Assessment of the viral load in the lung tissue of old mice showed that the influenza vaccine formulated with α-tocopherol at a dose of 10 mg decreased the virus load in these groups as compared with the mere vaccine (p = 0.0187).
Lung viral load in the young mice
Examination of the viral load in the lung tissue of young mice showed that the vaccine formulated in α-tocopherol at concentrations of 1 and 5 mg leads to decreased viral loads in these groups, as compared with the mere vaccine (p = 0.0001 and p = 0.0377, respectively); however, no clear effect was found at the dose of 10 mg, when compared with the mere vaccine (p = 0.9999).
Additionally, it appears that α-tocopherol at a concentration of 1 mg was more effective, among the three concentrations, in the controlling of the viral load as compared with the vaccine formulated in α-tocopherol at concentrations of 5 and 10 mg (p = 0.0001 and p = 0.1307, respectively) (Fig. 8).

Viral load in the lung tissue of young mice after experimental intranasal challenge. Influenza vaccines formulated with α-tocopherol at the doses of 1 and 5 mg decreased the viral load in these groups as compared with the mere vaccine (p = 0.0001 and p = 0.0377, respectively), but not at the dose of 10 mg.
Survival rate of aged mice
The survival rate of experimental aged mice after viral challenge showed that different vaccine formulations were effective in the reducing mortality rate of aged mice (Fig. 9). In fact, survival results showed that mice immunized with the mere vaccine resulted in a survival rate of 35%, while the influenza vaccine formulated with α-tocopherol at concentrations of 1, 5, and 10 mg increased the survival rate in these groups as compared with the mere vaccine (55.5%, 48.6%, and 57.1%, respectively). It also appears that among the three concentrations, the influenza vaccine formulated with α-tocopherol at concentrations of 1 and 10 mg was more effective in the increased survival rate than that formulated with α-tocopherol at a concentration of 5 mg; however, the increase in the survival rate was about 10% and not significant.

The survival rate of aged mice after intranasal challenge with the mouse adapted H1N1 virus. Survival results showed that mice immunized with the mere vaccine resulted in a survival rate of 35% and the addition of α-tocopherol to the vaccine formulation at the doses of 1, 5, and 10 mg increased the survival rate in these groups as compared with the mere vaccine (55.5%, 48.6%, and 57.1%. respectively).
Survival rate of young mice
The survival rate of young mice after viral challenge showed that different vaccine formulations were effective in the decreased mortality rate of young mice. In fact, survival results showed that vaccination in the young mice group resulted in a survival rate of 62.5%, while the influenza vaccine formulated with α-tocopherol at the concentrations of 1, 5, and 10 mg led to the increased survival rate in these groups, as compared with the mere vaccine (87.5%, 87.5%, and 77.14%, respectively). It also appears that among these three concentrations, the influenza vaccine formulated with α-tocopherol at concentrations of 1 and 5 mg was more effective in increasing survival rates, as compared with that formulated with α-tocopherol at a concentration of 10 mg, but the increase in survival rate was about 10% and not significant (Fig. 10).
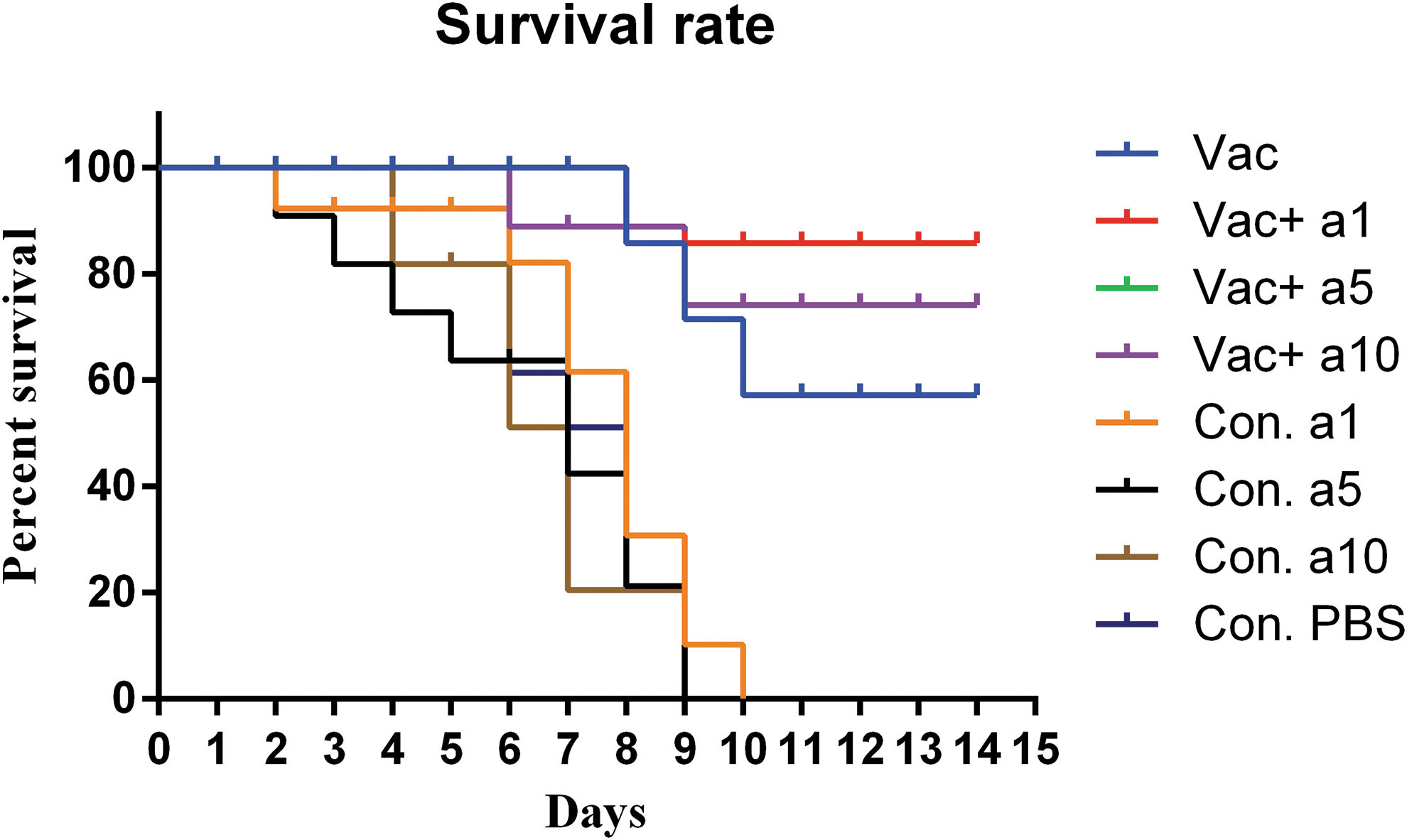
The survival rate of young mice after H1N1 viral challenge. Vaccination of young mice resulted in a survival rate of 62.5% and the influenza vaccine formulated with α-tocopherol at doses of 1, 5, and 10 mg increased the survival rate as compared with the mere vaccine (87.5%, 87.5%, and 77.14%, respectively).
Discussion
Although a variety of vaccines have been developed for influenza, there is a significant annual death from the influenza virus. This is because of low responses of the influenza virus to the available vaccines due to the decline in the immune response of aged people, resulting in increased mortality in these individuals (38,39). Therefore, there is a great need for development of a more effective influenza vaccine in the elderly. Herein, α-tocopherol was added to the influenza vaccine formulation to improve immune responses. For stronger responses to the vaccine, we hypothesized that empowering the responding cells may change the potency of the influenza vaccine in the aged mice. Therefore, we designed this study and vaccinated both elderly and young mice to explore the potency and efficacy of the vaccine in these different immunophysiology groups.
It is clear that T cell function and cytokine responses, especially IFN-γ response, dramatically decrease with growing ages (28,46). In fact, aging is followed by a decreased capacity to generate the IFN-γ cytokine response; this can explain the increased incidence and case fatality in the elderly people caused by viruses and intracellular pathogens (34). Therefore, restoring T cell function and cytokine response in the elderly may improve the response to the vaccines and, in the other words, increase vaccine potency, especially the influenza vaccine (28). Results from the IFN-γ cytokine response, as a Th1 pattern, showed that the vaccine formulated in α-tocopherol leads to a dramatic increase in the IFN-γ cytokine in the aged mice; nevertheless, no dramatic increase was observed in the young mice.
A study conducted on vitamin E supplementation in healthy elderly subjects showed that vitamin E is able to improve Th1 and cellular immune responses in aged people (29). In the study carried out by Han et al., vitamin E supplementation of old mice infected with the influenza virus showed an increase in the IFN-γ cytokine production and protectivity effect (17).
A study conducted by Meydani et al. on supplementation with vitamin E in healthy elderly showed a 65% increase in the delayed-type-hypersensitivity (DTH) reaction, as a parameter of cellular immune response. Other findings from this study indicated that a level of vitamin E greater than the routine level enhances T cell-mediated function in healthy elderly persons (32). In addition, several other studies confirmed positive effects of vitamin E on the polarization to the Th1 pattern in the elderly, as we achieved in the influenza vaccine model as an adjuvant (25,30,44).
IL-4 measurements showed an increase of about 72% in the aged, but not in the young mice, indicating that α-tocopherol in the aged group improved Th2 immune responses. This finding is consistent with other studies demonstrating improvement of IL-4 by α-tocopherol as an adjuvant (9,26,35). It is clear that Th2 responses, through secretion of critical cytokines, provide all needs for antibody responses (6). We also assessed antibody responses against the influenza vaccine through the potency in the engagement of their ligand on the virus particle and the ability to block the virus binding to the bird red blood cells. In addition, the HI test was performed to evaluate the antibody response for the vaccinated mice (16).
Results from the HI test showed that the addition of α-tocopherol to the vaccine formulation results in an increase in the antibody response in both aged and young mice, demonstrating its more inhibitory effect on HI assay. It appears that α-tocopherol showed the highest inhibitory effect at the dose of 1 mg in aged and young groups. This finding showed α-tocopherol concentration is a critical parameter in the vaccine formulation to improve the humoral immune response.
Various studies demonstrated the positive effect of α-tocopherol on humoral immune responses (14,24). In a study on adjuvant activity of α-tocopherol in a DNA vaccine model of H1N1 influenza Karlsson et al. showed a significant increase in humoral immune responses, when compared with the mere vaccine (21). Our results are consistent with this finding, in which α-tocopherol improved humoral immune responses in both young and aged mice. A study on supplementation with vitamin E in healthy elderly showed a sixfold increase in the antibody titer to hepatitis B and tetanus vaccine compared with the placebo group.
In addition, results from this study revealed that individuals with higher levels of serum α-tocopherol (vitamin E) after supplementation had stronger humoral immune responses to hepatitis B and DTH response. It appears that a higher level of vitamin E greater than the routine dose is required to improve immunologic parameters in healthy elderly persons (32).
Antibodies are important tools for neutralizing viral particles and preventing the virus from binding to the target cells (6). In addition, this mechanism can decrease the viral load and, thereby, decrease the mortality rate. After immunogenicity studies, the efficacy of the vaccine in the young and old mice was assessed by experimental intranasal influenza virus challenge to show the protectivity effect of vaccine formulations in both young and aged mice, as two different immunophysiology groups. Results from the viral load in the lungs of mice showed that α-tocopherol used in the vaccine formulation led to a decreased viral load in both young and aged mice. Here, the outcome of α-tocopherol as adjuvant on the vaccine efficacy was different in young and aged mice.
In fact, α-tocopherol at a concentration of 10 mg showed the lowest viral load in the aged mice, while 1 and 5 mg of α-tocopherol were found to be the lowest viral load in the young mice. This finding showed the α-tocopherol concentration is a critical parameter in the vaccine formulation for the elimination of viral particles in the lungs; in the vaccine formulation, the dose of this molecule should be carefully considered according to the immunophysiology of the host for aged and/or young population. Several studies demonstrated that α-tocopherol could decrease viral loads by various mechanisms, especially through modulation of cellular and humoral immune responses (8,12).
The survival rate of experimental young and aged mice showed the protectivity effect of novel vaccine formulation in the survival of challenged mice. Immunization of aged mice with the mere vaccine led to a surviving rate of 35%, while α-tocopherol, at the doses of 1, 5, and 10 mg, resulted in the increased survival rate to 55.5%, 48.6%, and 57.1%, respectively. This finding demonstrated that the α-tocopherol-formulated vaccine increased not only vaccine immunogenicity in the aged mice, but also vaccine efficacy. The survival rate of young mice was 62.5% after being immunized with mere vaccine, while α-tocopherol at the doses of 1, 5, and 10 mg increased the survival rate to 87.5%, 87.5%, and 77.14%, respectively. This finding demonstrated that α-tocopherol in the vaccine formulation was useful for young mice to increase the vaccine efficacy.
Various strategies have been reported to increase vaccine efficacy in the elderly, such as changes in the immunization route, increases in the vaccine dose, and presentation of new adjuvants (4, 42). In a study, Onchocerca volvulus activation-associated secreted protein-1 (Ov-ASP-1) was used as a potential adjuvant for the influenza vaccine, which improved the protection rate in the elderly (19).
In another study, a split H1N1 influenza vaccine admixed with the TLR4 agonist, the glucopyranosyl lipid adjuvant formulated in a stable oil-in-water emulsion, improved vaccine efficacy in the aged mice (5). Here, α-tocopherol in the influenza vaccine formulation improved immunogenicity and efficacy of the influenza vaccine in the both aged and young mice. However, this formulation was more effective in the polarization toward the Th1 response in the aged group rather that young mice. We strongly believe that the difference in the IFN-γ cytokine response in aged and young group may be due to the immunophysiology of aged mice, and the mechanism of α-tocopherol, as a vaccine adjuvant in elderly, may induce a rejuvenation mechanism (7) and/or antioxidant activity (23) that act and restore the function of aged cells to some extent like a young cell.
Conclusion
In general, it appears that α-tocopherol can be used as a potential adjuvant in vaccine formulation for the purposes of the aged group and can also empower old cells to increase the effectiveness of the vaccine in the aged people. Results from this study encouraged us to follow this point that targeting the immunophysiology of immune cells from aged people, using novel adjuvants, may result in normalization of immunophysiology status of responding cells, thereby improving immune responses to vaccines and also vaccine efficacy.
Footnotes
Acknowledgments
The authors are grateful to Borna Zist Pazhohan Knowledge Company and their staff for funding and technical support for this project. We thank Dr. Akbar Khorasani for vaccine formulation from the Department of FMD of Razi Vaccine and Serum Research Institute of Iran (Karaj, Iran).
Author Disclosure Statement
Y.E. is from the Academic Center for Education, Culture and Research (Tehran, Iran), Tehran University of Medical Sciences (Tehran, Iran), and Islamic Azad University (Tehran, Iran); Y.V. is from Islamic Azad University; P.K. is from the Academic Center for Education, Culture and Research and Tehran University of Medical Sciences; M.A.-A. and A.A. are from Larestan University of Medical Sciences (Larestan, Iran); M.T. is from the Agricultural Research Education and Extension Organization (Karaj, Iran); and M.M. is from the Academic Center for Education, Culture and Research, Tehran University of Medical Sciences, and Pasteur Institute of Iran (Tehran, Iran), all where education and research are the primary functions.
Funding Information
This study was supported by Department of Research and Development of Contract no. 98-03-12 with Borna Zist Pazhohan Knowledge Company and also partially by a grant from Pasteur Institute of Iran.