
Abstract
There are hundreds of coronaviruses, most of which circulate among animals, yet there are seven types that infect humans. Three of them can cause severe acute respiratory illness—SARS-CoV, SARS-CoV-2, and MERS-CoV. Other HCoV-229E, HCoV-OC43, HCoV-NL63, and HCoV-HKU1 usually cause only mild to moderate upper respiratory tract infections. These four coronaviruses are called seasonal, because they are continuously circulating among human population and are responsible for up to 30% of all respiratory tract infections. Genetically, these low-pathogenic types are related to SARS-CoV-2. That is why questions concerning the cross-reactivity and cross-neutralization between antibodies against different types of coronaviruses have been raised. We addressed these questions by using enzyme-linked immunosorbent assays and targeted next-generation sequencing (NGS). We established the upper respiratory infection etiology for three patients who had been vaccinated with Sputnik V and tested positive on anti-SARS-CoV-2 antibodies. The symptoms included sore throat, nasal congestion, and myalgia. Their blood serum was analyzed for anti-SARS-CoV-2 antibodies in dynamics: before vaccination, and after the first and second dose of the vaccine. After the second dose, all patients were positive for IgG antibodies against SARS-CoV-2. The targeted NGS panel sequencing data analysis showed that these patients were infected with common coronavirus HCoV-OC43. These results suggest that S protein-targeted vaccine-induced antibodies against SARS-CoV-2 are not protective against seasonal coronavirus HCoV-OC43.
Introduction
Coronaviruses are a diverse group of viruses mainly infecting animals. Nevertheless, some of them cause infections in humans. Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) is a novel member of the Coronaviridae family. The current COVID-19 pandemic once again showed the importance of animal coronaviruses to humans. Seasonal coronaviruses such as HCoV-229E, HCoV-OC43, HCoV-NL63, and HCoV-HKU1 are widely spread among the human population and responsible for 15–30% of common cold cases in human adults mainly causing mild upper respiratory tract diseases (5).
However, in infants, the elderly, and immunocompromised individuals, they may cause severe diseases, such as pneumonia. HCoVs cause epidemics every 2–3 years with high reinfection probability due to short-lasting immunity. The antibodies against seasonal HCoVs are common in humans. Recently immune response against SARS-CoV-2 was observed in individuals without any exposure history to the virus, indicating possible cross-reactivity of an immune response against seasonal HCoVs and SARS-CoV-2 (3).
The major target of neutralizing antibodies is the S protein of SARS-CoV-2. N protein also acts as a strong antigen to stimulate immune response during the infection (8). SARS-CoV-2 and SARS-CoV appear to have antibody cross-reactivities against S-protein (6). Other studies showed the evidence for significant cross-reactivity between antibodies to SARS-CoV-2 and seasonal HCoVs (7), but there is still no evidence for cross-protective immunity.
Here we suggested that vaccine-induced antibodies against S protein of SARS-CoV-2 are not protective against another member of the Coronaviridae family HCoV-OC43. We examined three cases of HCoV-OC43 infection in patients, who previously tested positive on IgG against SARS-CoV-2. The etiology of infection was established using the next-generation sequencing (NGS)-based method that included all known human coronaviruses.
Materials and Methods
Patients and samples
The mean age of patients was 35 years (range 29–40) (Table 1). Common symptoms among individuals were a sore throat, nasal congestion, and myalgia. All participants provided written informed consents. Nasopharyngeal swabs were collected from adults with upper respiratory tract infections in February 2021. The swabs were stored at −65○C. Vacuum-sealed test tubes with coagulation activators Vacuette (Greiner Bio-One, Austria) were used for venous blood sampling. Test tubes with blood were centrifuged at 2000 × g for 15 min. Blood plasma separated from erythrocyte mass was divided into aliquots and stored at −80°C.
Study Participant Information
Enzyme-linked immunosorbent assays
Titers of IgG antibodies against SARS-CoV-2 were determined with a semiquantitative enzyme-linked immunosorbent assay Antigma G (Generium, Russia). In total, 100 μl of serum 1:10 diluted was incubated in plate wells for 30 min at 37°C followed by washing with phosphate-buffered saline (PBS) four times, adding 100 μL of human IgG conjugated with horseradish peroxidase, incubation for 30 min at 37°C, and washing with PBS again. Then 100 μL of 3,3′,5,5′-tetramethylbenzidine (TMB) was added and the plate was incubated in the dark for 10 min before stopping the reaction with 100 μL of 1N sulfuric acid. The optical density was measured with a microplate photometer Multiscan FC (Thermo Scientific) at 450 and 620 nm.
Extraction of viral nucleic acids and reverse transcription
The nucleic acids were extracted using QIAamp One-For-All Nucleic Acid Kit (QIAGEN) according to the manufacturer's instructions. Reverse transcription was performed in 20 μL containing 10 mM Tris-HCl (pH 8.6), 5 mM MgCl2, 0.05 M KAc, 10 mM dithiothreitol, 0.25 mM deoxyribonucleotide triphosphate (dNTP), 1 U of M-MuLV reverse transcriptase (Biosan, Novosibirsk, Russia), 50 mM random N9 primers, and extracted nucleic acids. The reaction was carried out in the T100 (Bio-Rad) according to the following program: 3 min at 75°C and 30 s at 5°C followed by 12 min at 35°C, 12 min at 42°C, 12 min at 48°C, 6 min at 52°C, 3 min at 95°C, and 30 s at 5°C.
Library preparation and sequencing
Primers for two conservative regions of common coronaviruses were constructed using the NGS-PrimerPlex software (4). Two-step polymerase chain reaction (PCR) was performed in 20 μL, containing 16 nM (NH4)2SO4, 65 nM Tris-HCl (pH 8.9), 0.2 mM dNTP, 3 mM MgCl2, 0.1% Tween-20, 1U of Taq-polymerase (Biosan), 0.3 μM primers, and cDNA. The PCR cycling conditions were as follows: 15 min at 96°C followed by 26 cycles of 10 s at 96°C, 1 min at 60°C, and 40 s at 72°C.
Afterward 2 μL of 5 μM index sequences was added to the mixture followed by another round of PCR. The second PCR step was carried out according to the following program: four cycles of 1 min at 96°C, 1 min at 60°C, and 1 min at 72°C. The PCR products were purified with AmPure magnetic beads and quantified. Sequencing was performed on the Illumina MiniSeq platform according to the manufacturer's instructions.
Analysis of NGS data
Reads were “demultiplexed” using bcl2fastq conversion software. Qiime2 (
These sequences were mapped to a preconstructed database containing reference genomes and microorganism taxonomy. Sequences with coverage >90% were given a taxonomic classification. Unmapped sequences were analyzed against the Nucleotide database (NCBI) using Blast (
Results
The titer of IgG antibodies against SARS-CoV-2 before and after the Sputnik V vaccination was determined dynamically in three participants during ∼14 weeks. Before vaccination, participants did not have IgG antibodies against SARS-CoV-2 (Fig. 1). After 2 weeks, there was still no significant changes in antibody titer. One week after the second vaccination, antibodies were detected in all three patients.
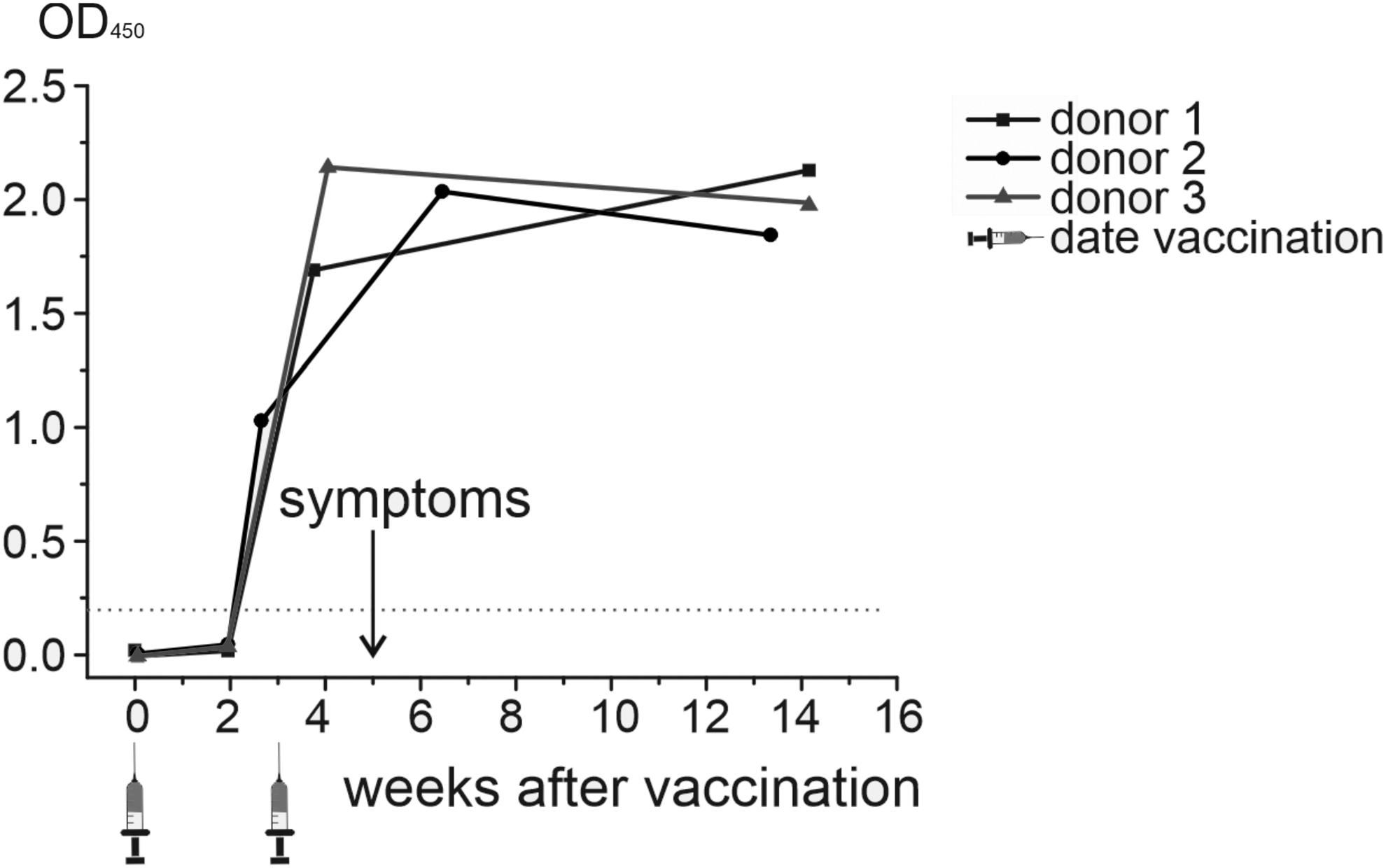
Antibody titer after vaccination.
Five weeks after vaccination, all participants developed symptoms like those caused by SARS-CoV-2—sore throat, nasal congestion, and myalgia. To determine the etiology, we used an in-house NGS panel targeted to all known coronavirus genomes. The average number of sequences generated per sample was 1481 (range 766–2175). Among these reads, 90.2% were filtered out (primer-dimers, nontarget amplicons formed on the unknown human genome fragments), leaving 434 reads to be analyzed (Table 2).
Sequencing Results
Read statistics from Qiime2 software.
Most of them (87.7%) were mapped to HCoV-OC43 genomes (Fig. 2). For other reads, taxonomy was unassigned, and no significant matches were found with Blast NCBI.
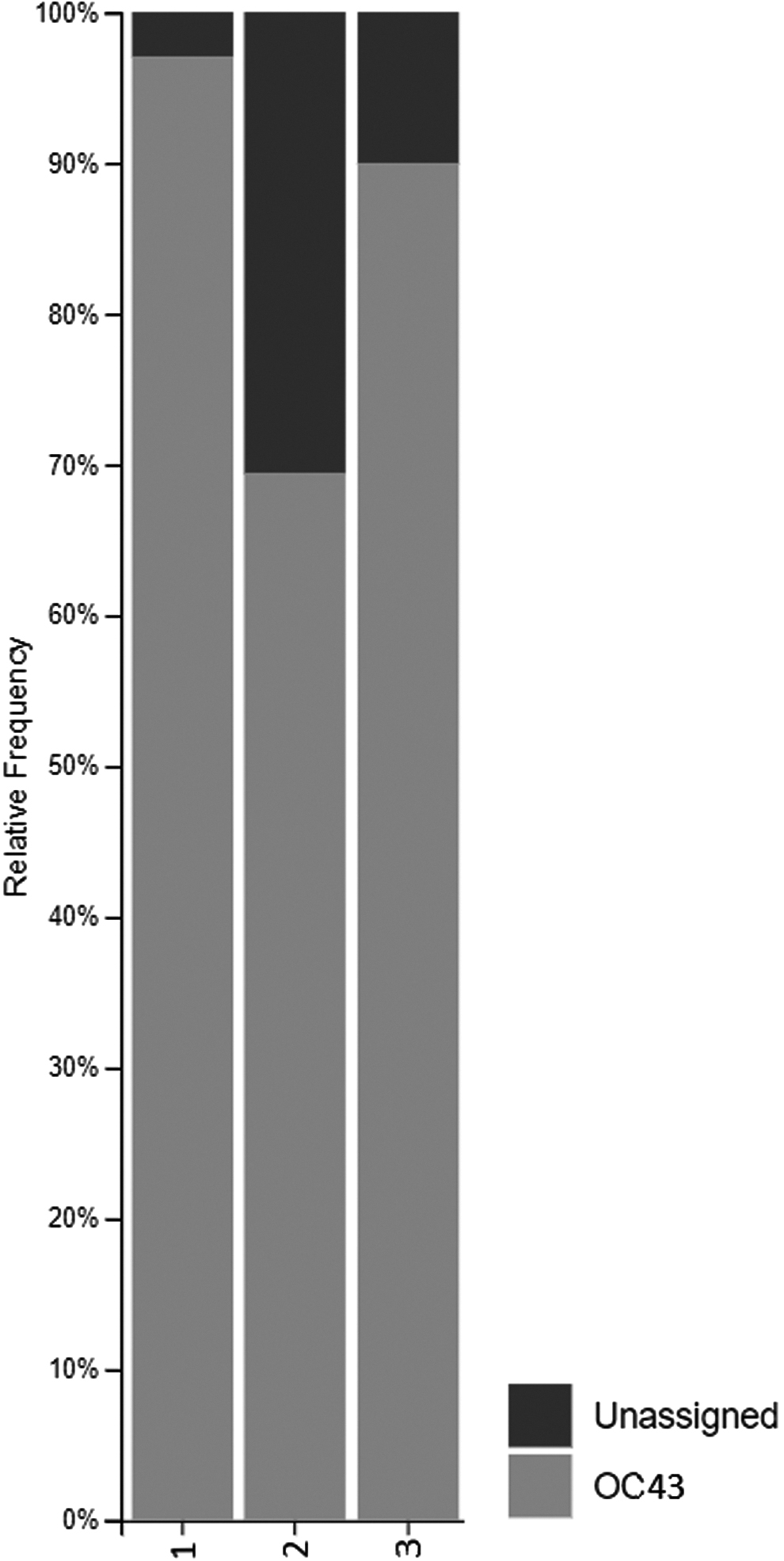
Sequencing results. Bar plot.
Discussion
Seasonal coronaviruses are common causative agents of human respiratory tract infections. Here we suggested that cross-reactivity for IgG antibodies against S protein of SARS-CoV-2 and HCoV-OC43 viruses might not be enough for effective disease prevention. Some studies showed that previous exposure to seasonal HCoVs may play a protective role in an immune response against SARS-CoV-2, others suggested that individuals who tested negative for anti-OC43 antibodies may benefit from vaccination against SARS-CoV-2 (2).
Although potential cross-reactivity of antibodies against these viruses has been reported in numerous studies, the real protective effect is yet to be understood. Cross-reactivity of anti-SARS-CoV-2 antibodies with SARS-CoV was detected in plasma samples as well as in mice studies. These cross-reactive antibodies mainly target S2 subunits of spike protein due to high homology between coronaviruses. It appears that cross-reactive antibodies are relatively common, but the cross-neutralizing response is rare. Only 1 out of 15 patients with COVID-19 was able to generate a cross-neutralizing response to SARS-CoV.
It is also unknown whether these non-neutralizing antibodies can implement a protective role in vivo, despite the lack of in vitro neutralization. Recent studies compared spike protein sequence homology among different coronaviruses. Results showed that the greatest homology was between SARS-CoV-2 and SARS-CoV, whereas HCoV-OC43 and HCoV-HKU1 was the lowest. The higher homology was toward the C-terminus of the protein, which is responsible for the insertion of the fusion protein in the host cell, whereas in the N-terminal regions, homology was significantly lower.
Results suggest that the majority of the S protein-specific cross-reactive responses were directed toward the S2 region of the C-terminus, which displays a relatively high homology between SARS-CoV-2 and other HCoVs (1). This indicates that the S protein carries common epitope(s) between SARS-CoV-2 and HCoVs. Most studies have demonstrated the cross-reactivity of antibodies against HCoVs, but to what extent this phenomenon can influence the course of the infection is not completely understood.
Thus, our results extend the knowledge about the cross-neutralizing effect of Sputnik V vaccine-induced anti-SARS-CoV-2 antibodies against seasonal HCoV-OC43 and suggest that they do not prevent OC43 infection.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The study was supported under the Russian State-funded budget project 1750-20/1755-21.