
Abstract
Monocytes are one of the principal immune defense cells that encounter infectious agents. However, an essential role of monocytes has been shown in the spread of viruses throughout the human body. Considering this dilemma, this study aimed to evaluate monocyte subsets and Human Leukocyte Antigen–DR isotype (HLA-DR) expressions in clinical coronavirus disease 2019 (COVID-19) cases. This prospective, multicenter, case–control study was conducted with COVID-19 patients and healthy controls. The patient group was divided into two subgroups according to disease severity (severe and non-severe). Three monocyte subsets (classical, CL; intermediate, INT; non-classical, NC) were analyzed with flow cytometry upon the patients' hospital admission. A total of 42 patients with COVID-19 and 30 controls participated in this study. The patients' conditions were either severe (n = 23) or non-severe (n = 19). All patients' monocyte and HLA-DR expressions were decreased compared with the controls (p < 0.05). Per disease severity, all monocyte subsets were not significant with disease severity; however, the HLA-DR expressions of CL monocytes (p = 0.002) and INT monocytes (p = 0.025) were more decreased in the severe patient group. In patients with various clinical features, NC monocytes were more affected. Based on these results, NC monocytes were more decreased in acute COVID-19 cases, though related various clinics decreased all monocyte subsets in these patients. Decreased monocyte HLA expressions may be a sign of immune suppression in severe patients, even when the percentage of monocyte levels has not decreased yet.
Introduction
Monocytes are nonspecific phagocytic, nonlobulated white blood cells that make up the immune system's inflammatory arm (29,33). Monocytes can be both sessile and mobile in the human body (2). The sessile version forms the reticuloendothelial system, and the mobile version is composed of peripheric blood monocytes and macrophages, or mature monocytes (2). Their nonspecific phagocytic nature often makes monocytes superior against infection in the inflammation battlefield.
Research has identified three monocyte subsets according to functionality and phenotype via CD16 and CD14 expressions: classical (CL) monocytes (CD14highCD16−), non-classical (NC) monocytes (CD14lowCD16+), and intermediate (INT) monocytes (CD14highCD16+) (16). The CL and INT monocyte subsets, which account for 90–95% of the monocytes circulating in the blood, combat mostly bacterial infections, whereas the NC monocyte type, composing the remaining 5–10% of circulating monocytes, combats primarily viral infections (4,31). Monocytes or macrophages also present antigens to T lymphocytes during their phagocytosis process, thus potentiating the specific immunoglobulin secretion of activated T lymphocytes (25,31). The release of cytokines, such as gamma interferons secreted from activated T lymphocytes, then encourages macrophages to secrete other systemically effective cytokines (interleukin [IL]-1, tumor necrosis factor alpha, IL-6) (21,29).
However, having a large amount of cytokine released in the human body all at once can be harmful. In this context, a cytokine storm, also called hypercytokinemia, may be severe or life-threatening and lead to multiple organ failure. In addition, other secondary macrophage functions, such as complement component production, plasminogen activation, and collagen denaturation, provide significant support as the human body fights infection, though especially viruses (21).
However, despite these robust defense mechanisms, viruses use monocytes to spread throughout the body (21). For instance, adenovirus infections, such as severe acute respiratory syndrome coronavirus (SARS) and Middle East respiratory syndrome coronavirus (MERS-CoV), block monocytes from releasing interferons from virus-infected cells as their primary preferred maneuver (10,26). The resulting inflammation and immune cell infiltrations, which are more intense due to suppressed monocytes, lead to more immune and epithelial cell apoptosis; thus, acute respiratory distress syndrome (ARDS) in the lungs becomes inevitable (8,22).
Coronavirus disease 2019 (COVID-19), a modern-day pandemic agent featuring mutagenic types, uses monocyte interferon suppression methods like other adenoviruses (7,24). The disease progresses destructively in many organs, particularly the lungs (13). This damage is caused by direct epithelial and macrophage infiltration, and many triggered inflammatory mediators (22). All these cumulative effects lead to macrophage activation syndrome, and then cytokine storm may develop; consequently, the disease can be fatal.
The years 2020 and 2021 have been a period in which research has reached a climax about COVID-19, given its pandemic state. There are also ongoing studies about the virus itself, treatment, mortality, and morbidity. Meanwhile, the role of monocytes in antigen presentation against infections is of undeniable importance (29). This study thereby aimed to determine how monocyte subsets and Human Leukocyte Antigen–DR isotype (HLA-DR) expressions are affected by COVID-19 disease progression.
Materials and Methods
Patient selection
This article presents a prospective, multi-centered, case–control study conducted in 2021 after receiving approval from the Selcuk University Ethics Committee. An informed consent form was obtained from all participants. The study's control group included volunteers over 18 years of age without any immunosuppressive comorbidities, such as cancer, chronic kidney disease, rheumatic disorders, and antibiotic prophylaxis or treatment, and the patient group featured patients over 18 years of age who had a positive severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) [COVID-19] polymerase chain reaction (PCR) test result.
Patients with a low glomerular filtration rate, comorbidities that could interact with hemogram parameters, steroid treatment, or a COVID-19 diagnosis not determined through a PCR test were excluded from the study. The patients' demographic characteristics and most current laboratory tests determining COVID-19 disease stage were noted.
The patients' clinical presentation classifications such as asymptomatic or mild, moderate, severe, and critical illness were redefined according to their clinical condition and laboratory data during the study (5).
The patients were recruited from two university hospitals and further divided into two groups based on their disease severity. Accordingly, patients who had clinical complaints (fever, cough, headache, sore throat, anosmia, and diarrhea) with/without minimal abnormalities in a chest X-ray or computed tomography were rated as non-severe. In contrast, those with serious clinical complaints (dyspnea, tachypnea [respiratory frequency ≥30 breaths/min], arterial partial oxygen pressure to inspired oxygen ratio [PaO2/FiO2] <300 mmHg, oxygen saturation [SpO2] ≤93%, and pneumonic infiltrate >50% within 24–48 h) were rated as severe (32).
Ratio definitions
For neutrophil to lymphocyte ratio (NLR) and lymphocyte to monocyte ratio (LMR) calculations, absolute counts were used instead of percentages, and threshold values were determined for each parameter duo as means and standard deviations. Accordingly, all NLR and LMR values were classified as low or high and then expressed as percentages. Cases with absolute lymphocyte count (ALC) <1,000 × 106/L were considered lymphopenic (6) and absolute monocyte count (AMC) <0.24 × 103/μL were considered monocytopenic (28).
Flow cytometric analysis
The monocytes of the controls and patients with COVID-19 were analyzed using FACSAria III flow cytometry (Becton–Dickinson, CA) built in one of the university's immunology laboratories with the FACSDiva version 6.1.3 software package, which assessed ethylenediamine tetra-acetate from anticoagulated peripheral blood samples gathered upon admission and before the treatment from the patients and during the study from the control group. Peripheral blood mononuclear cells (PBMCs) were isolated using ficoll-histopaque density gradient centrifugation.
PBMC was stained with anti-CD14-PerCP-Cy5.5 (5 μL, clone HCD14; Biolegend, San Diego, CA), anti-CD16-PE (5 μL, clone 3G8; Biolegend), and anti-HLA-DR-APC/Cyanine7 (5 μL, clone L243; Biolegend) and incubated for 20 min at RT in the dark. After incubation, PBMCs washed with 2 mL of cell staining buffer (Biolegend). Flow cytometric analysis of these cells was performed at least 100,000 acquired events.
Monocyte subset analysis
For the monocyte analysis, monocyte ratios were determined per a PBMC gate, excluding lymphocytes. The three monocyte subsets (CL, INT, and NC monocytes) were defined in the participants' monocyte population according to CD14 and CD16 expression profiles (Fig. 1) (35). The HLA-DR expression level was detected as median fluorescence intensity (MFI) in the monocyte subsets (Fig. 2).
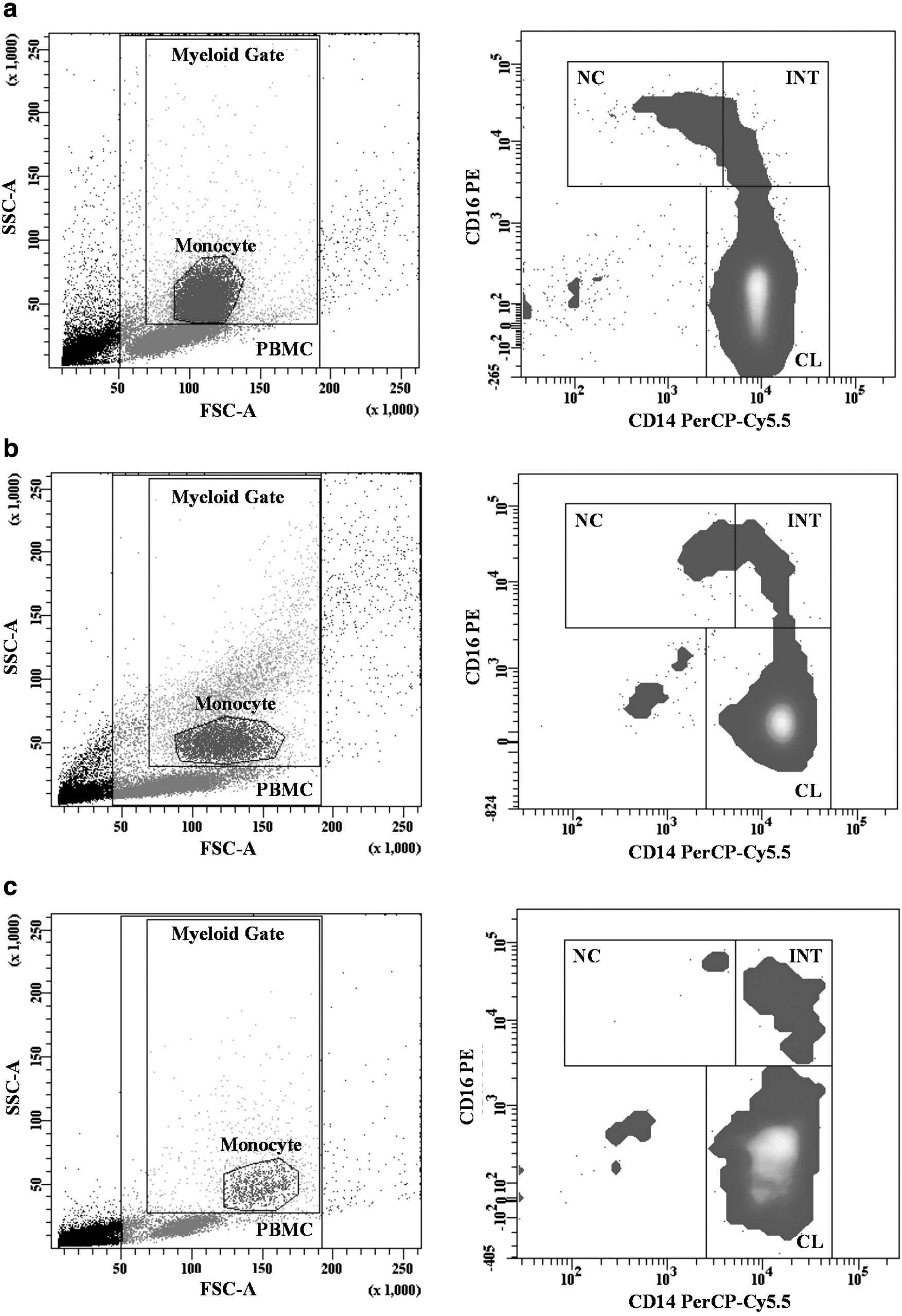
Gating strategy of monocytes subsets. Monocytes were gated in the myeloid gate of the PBMC population on the FSC-SSC dot plot graph, monocyte subsets were identified according to their CD14 and CD16 expressions.

HLA-DR expression of all events, monocyte and monocyte subsets. Surface HLA-DR expressions in all events, total monocytes and monocyte subsets were determined as MFI. The histogram plot of HLA-DR expression in all events, monocytes and, CL, NC, and INT monocyte subsets is given in figure
Statistical analysis
Statistical analyses were performed in SPSS version 21 (SPSS, Inc., Chicago, IL). The data distribution was determined with the Pearson correlation used to compare normally distributed data and the Spearman correlation for non-normally distributed data. To identify the monocyte subsets' significance between the patient and control groups, an independent t-test was applied (n > 30, normally distributed data). In contrast, a Mann–Whitney U-test was used for non-dispersed data or n < 30. Accordingly, nonparametric tests were preferred for the subgroup analyses due to the decrease in the group's patient numbers. The chi-square test was used to make the intergroup significance evaluations. For all statistical significances, p < 0.05 was considered significant.
Results
This study included a total of 42 patients (mean age 51.6 ± 14.2 years) and 30 controls (mean age 46.8 ± 16.6 years). The female and male populations were 20:22 in the patient group and 12:18 in the control group.
The average length of patient hospitalization was 10.2 ± 3.8 days. In the control group, four volunteers had diabetes mellitus (13.3%), seven were hypertensive (23.3%), and none of them had a history of COVID-19 disease. Pneumonia was prevalent in 30 patients (71.4%). One patient died after the study was conducted. The demographic and clinical characteristics of both patient subgroups are summarized in Figure 3. The monocyte subset percentages of the patients per-flow cytometry were as follows: 81.15 (76.4–86.25) for CL monocytes, 2.75 (0.97–6.5) for NC monocytes, and 11.85 (6.92–18.15) for INT monocytes.

Heatmap of the demographic and clinical characteristics of the patient subgroups, made with ClustVis. The patient features are displayed as colors ranging from red to violet, as shown in the key. Both rows and columns are clustered using directly associated patient counts. ALC, absolute lymphocyte count; AMC, absolute monocyte count; ANC, absolute neutrophil count; COPD, chronic obstructive pulmonary disease; CRP, C-reactive protein; LFT, liver functions tests; SpO2, blood oxygen level.
The patient and control groups were statistically well matched in terms of age (p > 0.05). There was a significant decrease in all percentage of monocytes (p = 0.043, F = 2.285, η 2 = 0.057) and HLA-DR expressions (p = 0.001, F = 7.817, η 2 = 0.148) in the patient group compared to the control group. In comparing the monocyte subsets between the two current groups, significant differences emerged in the evaluations of the NC and INT subsets. A decrease was stated in the percentage of CL monocytes and NC monocytes in the patient group; conversely, an increase was found in INT monocytes (Fig. 4a). However, the HLA-DR percentage levels decreased in the CL (p = 0.001, F = 10.89, η 2 = 0.160) and the NC (p = 0.001, F = 28.019, η 2 = 0.214) monocyte subsets were almost the same level in INT monocyte subsets (p = 0.423) (Fig. 4b).

In a comparison of groups classified by disease severity, even though there were no statistically significant decreases in all percentages of monocyte levels in the severe group, there were in the HLA-DR expressions. The decrease in overall HLA-DR MFI where the contribution of the CL (p = 0.002, η 2 = 0.229) and the INT (p = 0.025, η 2 = 0.121) monocyte subsets was more remarkable than the NC monocytes was statistically significant in the severe group (p = 0.005, η 2 = 0.193) (Fig. 5a). Moreover, there was a lower percentage of monocytes positive for HLA-DR in the severe group (96.30 ± 3.81) compared to the non-severe group (97.29 ± 2.96). On the contrary, the percentage of monocyte subsets levels did not differ in disease severity (p > 0.05).
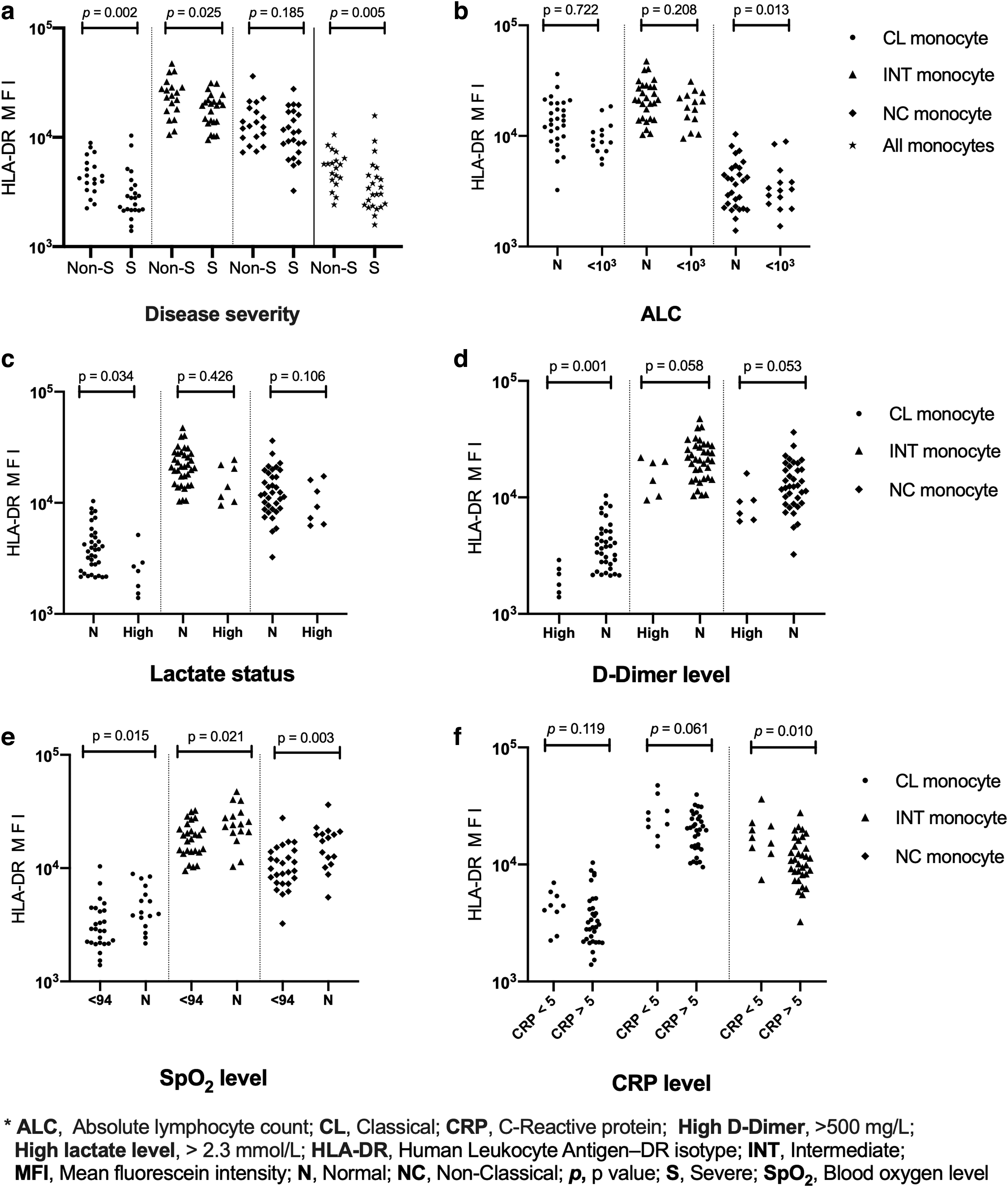
In comparing the clinical and laboratory parameters such as lymphopenia, hypoxemia, high C-reactive protein (CRP), lactate, or D-Dimer levels in disease severity, none of the monocyte subset percentage levels differed statistically. However, as shown in Figure 5b, lymphopenic patients (n = 14) have statistically significantly less HLA-DR expression of NC monocytes than non-lymphopenic patients (n = 28) (p = 0.013, η 2 = 0.147).
In addition, the HLA-DR expressions of CL monocytes at high lactate levels (>2.3 mmol/L) revealed a statistical significance (p = 0.034, η 2 = 0.109) (Fig. 5c) (30). Another statistical significance was found for the HLA-DR expression of CL monocytes (p = 0.001, η 2 = 0.227) in patients with a high D-dimer level (>500 mg/L; Fig. 5d). At low O2 saturation levels (SpO2 ≤93%), statistical significance emerged in all monocyte subsets, with the highest statistical significance for the HLA-DR expression of NC monocytes decrease (p = 0.003, η 2 = 0.212; Fig. 5e). In patients with high CRP values (>5 mg/L), statistical significance persisted in the HLA-DR expression of NC monocyte subset reduction (p = 0.010, η 2 = 0.156) (Fig. 5f).
In patients with low LMR results (<2.85), the HLA-DR expression of INT monocytes (p = 0.047, η 2 = 0.095) and the HLA-DR expression of NC monocytes (p = 0.035, η 2 = 0.108) were decreased, in contrast, in those with low NLR results (<4.23), the CL monocytes percentages (p = 0.049, η 2 = 0.095) and NC monocytes percentages (p = 0.002, η 2 = 0.217) were markedly decreased.
There were many correlations between the outcomes of the study. The prominent ones are summarized (Table 1).
Notable Correlations in The Study
Correlation coefficient.
O2 saturation level.
ALC, absolute lymphocyte count; ANC, absolute neutrophil count; CL, classical; CRP, C-reactive protein; HLA-DR, Human Leukocyte Antigen–DR isotype; INT, intermediate; LMR, lymphocyte to monocyte ratio; NC, non-classical; NLR, neutrophil to lymphocyte ratio.
Discussion
In this prospective, case–control study, all three monocyte subsets were affected in acute SARS-CoV-2 infection. Of the subsets, NC monocytes were most decreased under various clinical and laboratory conditions, such as lymphopenia, monocytopenia, septicemia, and hypoxemia. Moreover, monocyte HLA-DR expressions were reduced in severe patients even if cumulative monocyte counts did not decrease in number.
As research has already indicated, viruses cannot survive for long without a host (19). Accordingly, they mostly reprogram host cells to replicate until the cells die or have normal cells transform into malignant forms (18). Viruses and monocytes must intersect during these events, as viruses spread throughout the body via monocytes (21). It may be assumed that NC monocytes combat viruses more than other monocyte subsets (18); however, all three subsets are involved in acute and severe virus infections (20).
Among the immune system's marvel cellular networks, the monocyte response to control virus infections can sometimes result in more remarkable disease advancement, including organ damage, cytokine storms, or autoimmune diseases because of excessive mediator release (29). In this context, scholars have reported different disease clinics during the COVID-19 pandemic to confirm monocyte activity in cases of this disease (15).
The levels of HLA-DR expressions are considered necessary in response to the infections as the number of monocytes (11). This correlated dynamicity makes HLA-DR expressions more prominent in disease prognosis (34). In patients with COVID-19, monocytes were found to reduce CD expressions, correlated with the disease severity (9). In addition, a decrease in the percentage of monocytes subsets, chiefly NC monocytes, was also reported in patients with severe disease. We similarly found that the most decrease in the monocytes subsets was in the NC subset.
Intriguingly, we noticed that monocyte HLA-DR expression had already begun to decrease in severe cases, even when there were changes in the percentage of monocytes that were not statistically significant. However, HLA-DR expressions of all monocytes subsets in the patient group were increased compared to the control group at baseline. This might be a clue to the onset of immune suppression. It is notable for confirming that since HLA-DR expression is an activation marker of inflammation (11), in severe COVID-19 cases, both decreases in the number of expression-related monocyte subsets and virus-mediated inactivation become more pronounced, resulting in decreased antigen presentation and activation.
In a recent study that confirmed this process, NC monocytes of the severe patients with COVID-19 were lower than in the control group (3). Additionally, the moderate patient group had a higher INT monocyte percentage. The study also reported an exceptional case in which the INT monocyte levels of two patients who died in the severe group were minimal. Another notable finding concerned HLA-DR expression levels, which were downregulated in the severe group. These findings in two studies confirm that the NC monocyte population tends to be lower in patients with severe COVID-19.
In a study conducted on patients with different clinical presentations of COVID-19, the percentage of monocyte levels variations emerged in the monocyte subsets of patients whose AMCs were similar to the control group (27). Specifically, the study showed that the number of NC monocytes decreased with the severity of infection, whereas CL monocytes increased; these results were consistent with the patients' high levels of released calprotectin.
In the current study, INT monocytes increased in the non-severe but not the control or severe groups (15). However, NC monocytes did decrease in the severe group. The researchers also found variable HLA-DR expressions in the severe group's CL monocytes. Similar to that study, there was no statistical difference between the AMCs in the present study's patient subgroups. Although the study also revealed a more significant decrease in NC monocyte in the severe group, this decrease was not statistically significant.
In a review from the end of the first year of the COVID-19 pandemic, the researchers found that SARS-CoV-2 not only can infect both monocytes and macrophages and cause defects in the immune response against the virus, but it also can cause organ damage, such as ARDS, and cytokine storms due to dysregulated monocyte functions (14). In addition, the results illustrated that the decreased monocytes would migrate to almost all kinds of tissues due to weakened immunity and reduce immunopathological complications (14).
Therefore, a decrease in NC monocytes can be expected due to their nature in traveling to tissues and other infection sites; NC monocytes are exposed to more viral agents that reprogram them to spread. Meanwhile, a decrease in CL monocytes may be attributable to opportunistic bacterial infections accompanying viral infections or CL monocytes' migration to tissues for transformation into macrophages (17).
However, in patients with hypoxemia, which was one of the main criteria for inclusion in this study's severe patient group, all monocyte subsets significantly decreased compared to the non-severe group. This result may be explained by the fact that hypoxemia causes all monocyte subsets to enter combat in a viral disease's acute phase. In our comparisons with other identified prognostic factors (high lactate and d-dimer level, lymphopenia, hypoxemia) in COVID-19 disease, we found that the NC monocyte subset significantly reduces HLA-DR expressions with disease severity.
Remarkably, the percentage of monocytes did not reveal a significant discrepancy between the severe and non-serious groups; a significant decrease was observed in the severe group in HLA-DR expressions. This may mean a more significant reduction in monocytes capable of overexpressing HLA-DR due to disease severity. Therefore, lower monocyte resistance may facilitate the virus's spread in the human body, and dysregulated monocyte distribution may trigger cytokine storms sooner.
Since the beginning of the COVID-19 pandemic, many prognostic laboratory results, such as lymphopenia, have been evaluated (23). In this context, in a study evaluating various hemogram ratios in response to whether the decrease or increase of these antigen-presenting cells affects COVID-19 prognosis, lymphocyte-to-monocyte ratios were found lower in non-survivors compared to survivors (1). Although monocytosis has been attributed to low LMR in patients with COVID-19 (12), lymphopenia can cause the same result. The mean LMR results in the current study were 2.85 ± 1.57. There was also a reduction in the INT and NC monocyte subsets after evaluating them in patients with an LMR result of <2.85.
In addition, after comparing patients with and without monocytopenia, NC monocytes were depleted in the monocytopenic group. A similar result arose when the patients were grouped as lymphopenic or non-lymphopenic: the NC monocyte subset was again lower in the lymphopenic group. In contrast, no statistical significance emerged when evaluating patients with monocytosis. These findings may help interpret the low LMR results via lower ALCs rather than higher AMCs.
The major weakness of this study was its application of the terms monocytopenia and monocytosis. Unfortunately, there are numerous references in the literature to this issue with insufficiently current and precise monocyte reference ranges. Another point was that the NC monocytes count was relatively low in our study. However, NC monocytes, which generally constitute 5–10% of circulating monocytes, may also be depleted in number due to the COVID-19 severity. Furthermore, the reduction in number would make the amount of HLA-DR expressions quite broad. Indeed, our study detected reductions in the NC monocytes expressions in many prognostic criteria that are considered essential in the COVID-19 severity.
Moreover, since this study was not limited to “Interleukin-6 test” results, it was not wholly possible to assume that decreased monocyte subsets in patients with low O2 saturation were linked to cytokine storms.
Conclusions
This study evaluated monocyte subsets in patients with COVID-19 and controls. The study confirmed that all monocyte subsets were partially affected in acute COVID-19 infections, with NC monocytes most affected at all evaluations. Decreased HLA-DR expressions in patients with severe COVID-19 may be a precursor of immune suppression. Accordingly, more work is necessary to determine monocyte interactions on the COVID-19 battlefield.
Footnotes
Authors' Contributions
Conceptualization: H.A., A.C.; Methodology: H.A., A.E.; Laboratory procedure; H.A., A.E., H.T.D.; Formal analysis and investigation: A.C.; Sample collection; S.S., D.E., H.A.C.; Writing—original draft preparation: A.C., H.A.; Funding acquisition: H.A.; Supervision: H.A.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the Scientific Research Project Found of Selcuk University under Grant (no. 19611034).