
Abstract
The effectiveness of inactivated H9N2 influenza vaccines is doubtful due to changes in antigenic regions of the virus hemagglutinin (HA) protein. One strategy for the development of the efficacious vaccine is the use of nanoparticles that display more immunogenic regions of the influenza virus. In this study, chitosan (CS)-based nanoparticles were developed as a delivery system for intranasal immunization using recombinant H9N2 virus HA1 and nucleoprotein (NP), for the induction of humoral and cellular responses. CS-HA1 and CS-NP nanoparticles were prepared by the ionic gelation method and characterized for their physicochemical properties and shape. The immunogenicity and the protective efficacy were evaluated by measuring antibody titers, T cell proliferation response, CD4+/CD8+ ratio, and quantitative real-time RT-PCR following intranasal administration of the prepared nanoparticles alone or in combination in chickens compared to an inactivated H9N2 vaccine. The average size, surface charge, and spherical structure of the synthesized nanoparticles showed high quality. Serologic analysis revealed that the immunization of inactivated vaccine groups resulted in strong influenza antibodies, which were significantly (p < 0.05) higher compared to the other groups. The vaccinated chickens with CS-HA1+CS-NP developed higher specific anti-influenza antibodies than in those vaccinated with each of rHA1 and rNP. Administration of a combination of the protein-based nanoparticles has stimulated the activation of both CD4+ and CD8+T cells and induced a significantly higher T cell proliferation. The viral shedding was significantly lower in CS-HA1+CS-NP and inactivated vaccine groups compared with other challenged groups. The data demonstrate the potential of CS-HA1+CS-NP nanoparticles for eliciting specific influenza antibodies and conferring protection in chickens.
Introduction
Various avian influenza virus subtypes cause respiratory infections in bird species that range from mild to severe disease. Influenza H9N2 virus has been endemic in various geographical regions since the late 1990s (7,20,27). The virus infection shows high economic losses during coinfection with other avian pathogens. The immunosuppressive effect associated with H9N2 viruses by altering the differentiation of lymphocytes or apoptosis has been reported (24,28,34). Commercial whole-virus inactivated vaccines elicit neutralizing antibodies, which are directed to the globular head of hemagglutinin (HA) glycoprotein (18). This immunodominant region (HA1) plays a key role in virus binding to the host cell receptor through sialic acid (21). The antigenic variants and immune evasion mutants of H9N2 viruses were caused mainly by amino acid substitutions within the antigenic sites and 220-loop of the receptor-binding site (11,31).
Massive vaccination against influenza contributes substantially to the control of influenza virus outbreaks, however, frequent isolations of the H9N2 virus from vaccinated farms cast doubt on the effectiveness of the current vaccines. Differences in the estimated effectiveness of commercial vaccines are due to the antigenic drift of HA; hence, these vaccines provide limited immune responses to the circulating novel antigenic varieties. Consequently, influenza vaccines should be reformulated to include the dominant HA protein (2,35). Updating vaccine seeds to match the current circulating viruses requires a regional laboratory network for isolating viruses and sequencing their genomes. Full-scale vaccine manufacturing from a new antigenic strain would only begin when a suitable vaccine seed was prepared, meaning that production of an effective oil-adjuvanted vaccine will time-consuming and takes ∼3–6 months (26).
One of the challenges to developing an effective vaccine against influenza is mediating cross-protection within the new antigenic variants. To overcome the limitation, many investigations have focused on targeting the nucleoprotein (NP). The NP-based vaccines can promote a more efficient protective response against influenza infection. Although the anti-NP antibody does not neutralize viruses, it can contribute to protection by increasing T cell responses associated with enhanced dendritic cell (DC) function (6,8,29). Nanoparticles-loaded influenza peptides represent one of the most promising alternatives for the traditional inactivated vaccines. These particles as potent adjuvants and vaccine delivery systems induce protective and long-lasting immune responses. The strong and persistent protection against influenza infection is attributed to their ability to stimulate a comprehensive immune response, including mucosal, humoral, and cell-mediated immunities (14,23). This study is evaluated humoral and cell-mediated immune responses conferred by chitosan (CS)-based HA1 and NP nanoparticles against the H9N2 virus.
Materials and Methods
HA1 and NP proteins preparation
All of the full-length HA and NP nucleotide and amino acid sequences of H9N2 viruses isolated from Iran were downloaded from GenBank and aligned by multiple sequence alignment program ClustalW (BioEdit 7.2) with a number of bootstraps value 1,000 replicates. The fragment corresponding to the HA1 gene was detected. The linear B cell epitopes from the conserved regions were predicted using the ABCPred server (
The potential cytotoxic T lymphocyte (CTL) epitopes were predicted using the mapping algorithms at
The genomic RNA of a local H9N2 isolate (
The cells were sonicated on ice and centrifuged to remove the debris and insoluble proteins. The soluble fraction was filtered through 0.45 μm syringe filter and purified by His-tag affinity chromatography (ProBond; Invitrogen). Concentration of the purified protein was determined using the Bradford assay. Then, the proteins were separated by 12% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and subsequently confirmed by western blotting. The molecular weights of the fragments were calculated by ExPASy Compute pI/Mw tool available at
CS nanoparticles-based HA1 and NP formulation
CS nanoparticles were prepared using ionic gelation by dropwise addition of an ice-cold sodium tripolyphosphate (TPP; Merck) solution into the CS (Sigma-Aldrich) solution at ratios 1:1 to 5:1 under a magnetic stirrer at room temperature. Particle size, polydispersity index, and surface charge were determined using Dynamic light scattering (DLS) with Malvern Zetasizer Nano ZS (Worcestershire, United Kingdom). The morphology was determined using a transmission electron microscope (TEM; Zeiss-EM10C-100 KV, Germany). Fourier-transform infrared spectroscopy (FTIR) spectrum in the range of 400–450 cm−1 with FTIR-S 8400 (Shimdzu, Japan) was recorded. Then 0.5, 1.0, and 1.5 volumes of each HA1 and NP proteins (30 μg, pH 6.7) were mixed with an optimum ratio of CS/TPP solution.
The suspensions were centrifuged at 10,000 g for 10 min at 4°C. The pellets were resuspended in HEPES buffer to measure the physicochemical properties and also the morphology as discussed above. Loading efficiency of the nanoparticles were determined in the collected supernatants.
Cytotoxicity assay
Chicken hepatoma (LMH) cells were seeded on 12-well plate (1 × 106 cells/well) and cultured for 24 h at 37°C in a 5% CO2 incubator. The monolayers were washed and then treated with CS-HA1, CS-NP, and CS nanoparticles, and also HA1 and NP proteins alone. Cell viability was evaluated by MTT assay at 48 h postinoculation.
Nanoparticle immunogenicity trial in chickens
Specific pathogen-free (SPF) chickens were randomly assigned to six groups (n = 10): phosphate-buffered saline (PBS), inactivated H9N2 vaccine formulated with MONTANIDE™ ISA70, CS, CS-HA1, CS-NP, and premixed CS-HA1+CS-NP (at a 1:1 ratio). The immunization trial was illustrated in Figure 1. Except for inactivated vaccine group, chickens in other groups received intranasally the related samples with prime at 0-day of age and two boosters with a 1-week interval. At 21 days postvaccination, chickens were challenged via the oral-nasal route with 107 tissue culture infectious dose 50 (TCID50/mL) of A/chicken/Iran/SS7/2011 (

Immunization schedule of chickens with influenza protein-based nanoparticles. After the second boosting and challenge, peripheral blood mononuclear cells obtained from each group were used in lymphocyte proliferation assay and detection of CD4+ and CD8+ T cell populations. The specific antibody titers were determined by HI and ELISA with 2-week intervals up to the end of the experiment. Virus shedding was determined by targeting the viral matrix gene in real-time RT-PCR. HI, hemagglutination inhibition.
Blood samples were collected before prime and the second boost, as well as at 2-week intervals until day 70. HA-specific antibodies were assessed with hemagglutination inhibition (HI) (5). NP-specific antibodies were determined using competitive NP-ELISA (BioChek) according to the manufacturer's instruction.
The specific T cell proliferation response was determined on days 28 of age. The peripheral blood mononuclear cells (PBMCs) were washed in DMEM medium, and viable lymphocytes were detected using Trypan Blue. The cells were seeded in a 96-well plate at 2 × 106 cells/well in DMEM supplemented with 5% fetal bovine serum. The viral recombinant proteins were added in lymphocyte suspension and incubated for 72 h at 37°C and 5% CO2. Phytohemagglutinin (PHA) at 10 μg/mL concentration was used as a positive control. Proliferation was assessed by incubating with 10 μL of MTT (5 μg/mL final concentration in PBS) in dark for 4 h at 37°C. The absorbance was read at OD540 following another incubation with 100 μL of DMSO.
The proliferative activity was expressed as a stimulation index (SI), which calculated as the mean OD540 of stimulated cells–the mean OD540 of blank/the mean OD540 of unstimulated cells. The CD4+ and CD8+ T cell populations were obtained from the PBMCs prepared on the 14th day after the third vaccinations. Cells were washed three times with PBS and plated on 96-well microplate with 2 × 106 cells/mL and then incubated with PE-labeled anti-CD4 and FITC-labeled anti-CD8 monoclonal antibodies (BD Biosciences) for 30 min at 4°C. After washing and centrifugation at 400 g for 5 min, the cells were analyzed by flow cytometer (BD FACSAria).
Statistical analysis
Data are presented as mean ± SD from three experiments. Statistical analyses were performed using one-way ANOVA, and p-value <0.05 was considered significant.
Results
The predict HA1 and NP linear B cell epitopes result are summarized in Table 1. Of the 21 predicted B cell epitopes in HA (data not shown), 13 epitopes were located in HA1 indicating the more antigenic property of the fragment than HA2. The Kolaskar and Tongaonkar's algorithm estimated the average antigenicity 1.029 for HA1 and 0.998 for NP, where the threshold value >1.00 was potentiality antigenic. Both proteins were subjected to predict the MHC class I-binding epitopes through the proteasome cleavage and TAP transport. Based on a cutoff of IC50 value <500 nM for binders, 11 CTL epitopes in NP showed the binding affinity with MHC class I alleles (Table 2).
Predicted B Cell Epitopes of HA1 and NP Proteins of H9N2 Influenza Virus
Binding Affinity (IC50 < 500 nM) Value Calculated for Potential Cytotoxic T Lymphocyte Epitopes of H9N2 Influenza NP Protein
Three epitopes at positions 456, 256, and 187 with the lowest IC50 < 50 nM showed the strong binding affinity. All predicted CTL epitopes for HA1 have IC50 value greater than 1,000 nM revealed the weak binding with MHC class I alleles or no binding.
The oligonucleotides encoding HA1 and NP were successfully expressed into the E. coli competent cells. The purified protein bands corresponding to HA1 (∼31 kDa) and NP (∼52 kDa) were appeared in SDS-PAGE and confirmed by western blot. Based on the analysis (data not shown), the optimal nanoparticles were produced in a ratio of 1: 2 of CS/TPP. Then, the HA1 and NP proteins were encapsulated into CS nanoparticles by ionic gelation method. The physical characterization of the CS-based viral protein nanoparticles are shown in Table 2. Zeta potential as an indicator of the stability revealed that the CS-based viral recombinant antigen nanoparticles in the ratio 1:2:1 of CS:TPP:recombinant protein (either HA1 or NP) were electrically stabilized. Their sizes were in unison with the DLS size spectra.
The nanoparticles displayed spherical morphology and good dispersion. Tracking nanoparticle analysis as well as TEM monographs revealed the uniform particle size distribution and shape at the ratio (Table 3 and Fig. 2). Protein loading efficiencies in CS nanoparticles are inversely related to the protein molecular weight. It found that the encapsulation efficiency of NP protein with higher molecular weight was lower than HA1 at the same concentrations.
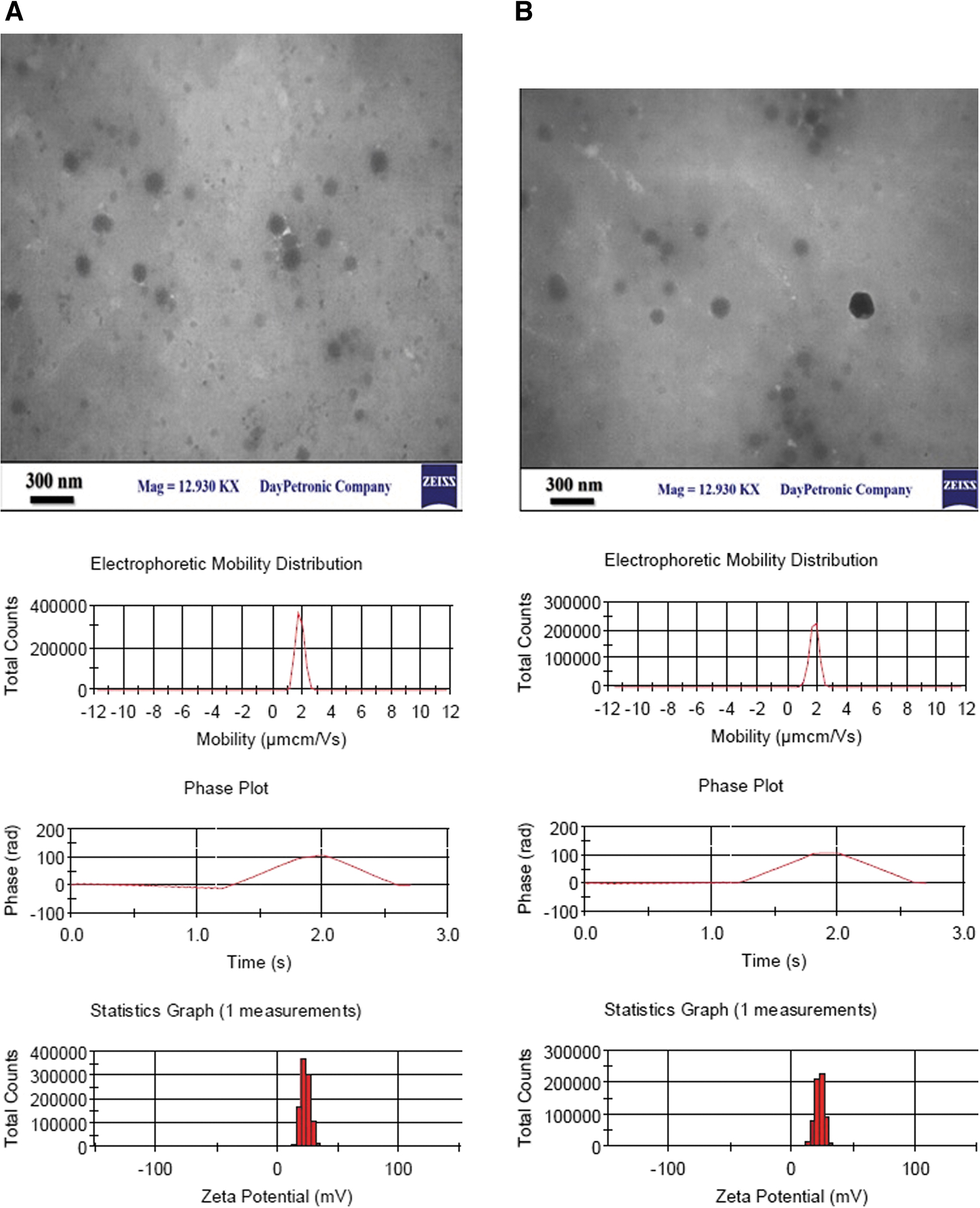
Electrophoretic mobility and transmission electron microscopy of influenza protein-based nanoparticles. Samples were negatively stained with 1% uranyl acetate. The conversion of electrophoretic mobility to zeta potential assumes that the particles are approximately spherical.
Properties of Chitosan and HA1- and Nucleoprotein-Loaded Nanoparticles at Different Ratios Generated by Ionic Gelation
The effect of viral proteins and the nanoparticle constructs on LMH cell viability was examined after 48 h of inoculation. Compared to the control cell, the cell viability was reduced in all treated cells, 90.4 ± 1.06 versus a range of 89.3% ± 5.7% to 88.4% ± 6.9%, which was not significant (p < 0.05). These results indicate that neither influenza virus proteins nor the prepared nanoparticles are toxic for LMH cells.
To validate the CS-based viral protein nanoparticles' immunogenicity, SPF chickens were primed at day 0 and boosted twice. The CS-HA1 induced HA-specific antibody titers, which remained low 2 weeks after prime then increased to 5 log 2. Chickens in this group had low HI antibody titers compared to the chickens vaccinated with CS-HA1+CS-NP that exhibited significantly (p < 0.05) a higher titer which reached to 6 log 2 at the end of experiment (Fig. 3). As expected, the inactivated vaccine produced higher titers of 8 log2. Sera from chickens inoculated with CS-NP failed to inhibit hemagglutination. The influenza NP-antibody levels remained low 2 weeks after prime in CS-NP-vaccinated group then increased to 4,123 (Fig. 4).
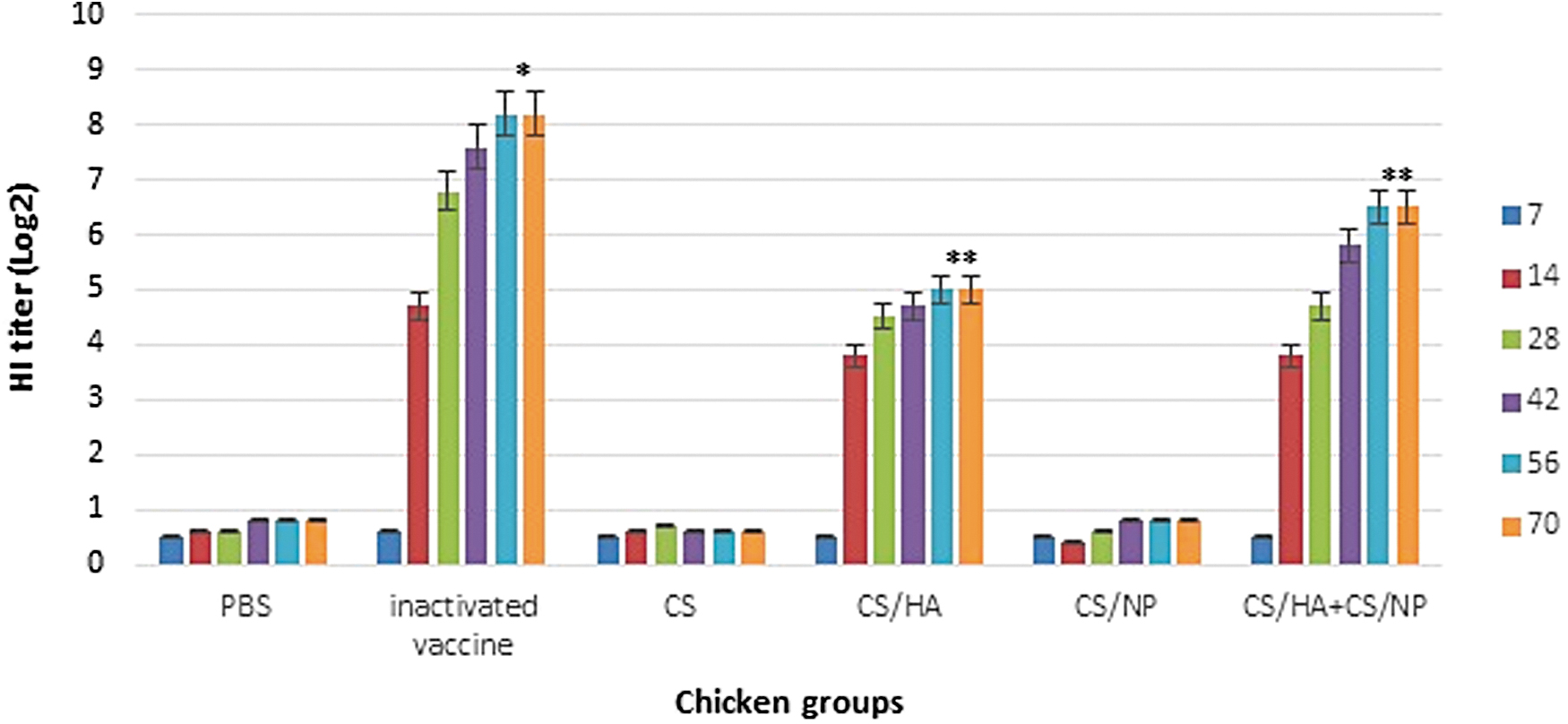
Induction of humoral immunity of influenza H9N2 protein-based nanoparticles in HI assay. *Significant difference p < 0.05 with other groups, **Significant difference with control and vaccinated groups.
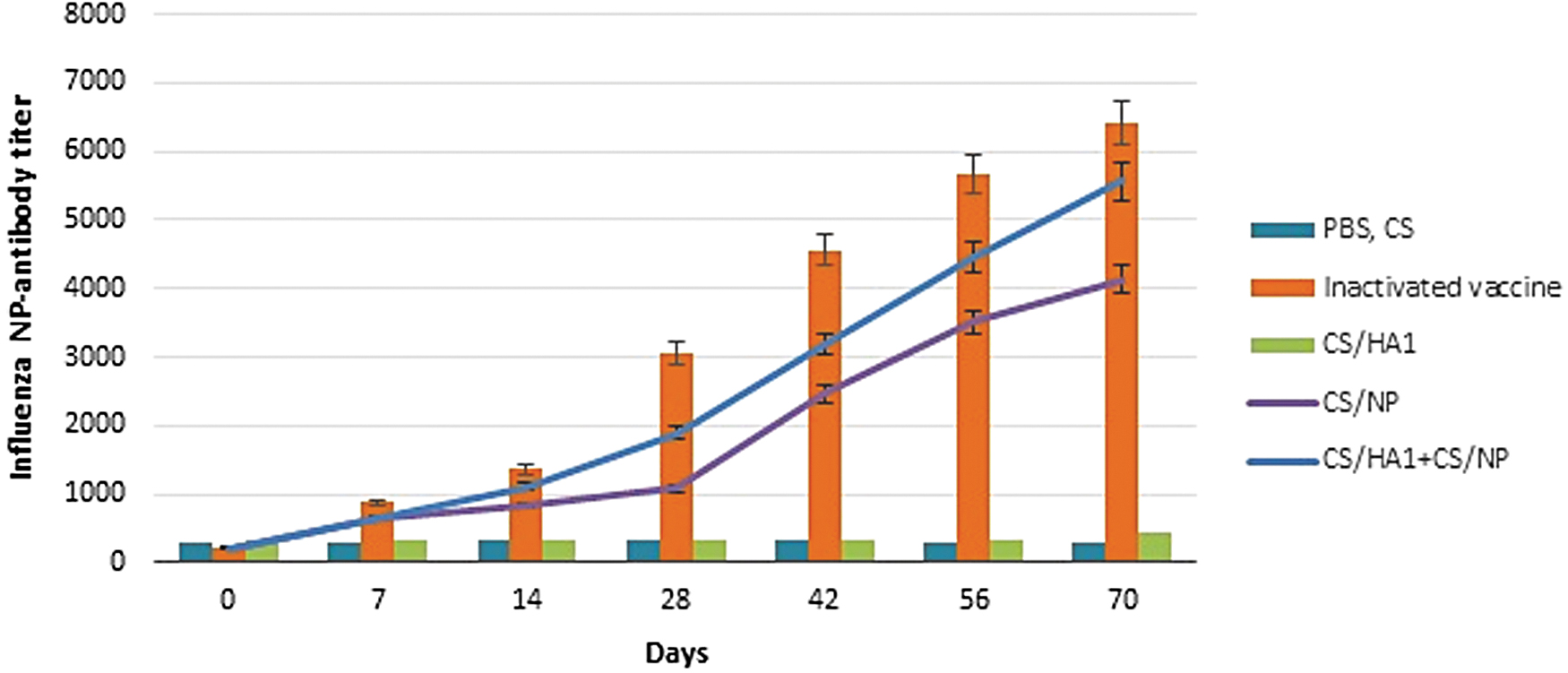
Induction of humoral immunity of influenza H9N2 protein-based nanoparticles in NP-ELISA assay. PBS and CS groups have similar antibody levels and plotted together on the chart. Bars indicate significant difference (p < 0.05). CS, chitosan; PBS, phosphate-buffered saline.
In contrast to the results obtained in CS-NP, no antibody responses were detected in CS-HA1 serum samples by the ELISA. Immunization with the CS-HA1+CS-NP resulted in a median titer of 5,753, which was higher (p < 0.05) compared to the CS-NP group. The combination of these nanoparticles has the superior capability in enhancing influenza-specific humoral responses compared to CS-NP alone. Mean serum IgG induced by the inactivated vaccine was significantly higher (p < 0.05) compared to the other groups. Changes in antibody titers could be observed after the first booster indicating that the inactivated vaccine is able to induce an earlier immune response compared to the CS-HA1+CS-NP.
The PBMCs obtained from chicken groups were examined for their ability to proliferate lymphocytes (Fig. 5). An increase in the SI value in CS-HA1, CS-NP, and CS-HA1+CS-NP groups were detected 2 weeks after the last vaccination. The lymphocytes of CS-HA1 and CS-NP groups were induced SI values of 2.38 and 3.64, respectively. A significantly higher (p < 0.05) proliferation was observed in CS-HA1+CS-NP group (SI = 3.76). The inactivated vaccine is formulated with MONTANIDE™ ISA70 and the oil adjuvant does not have the ability to induce a cellular immune response. Therefore, the SI value calculated for inactivated vaccine group was less than the cutoff (2.0). PHA-stimulated PBMCs showed robust proliferation in each of the chicken groups (4.23 on average). To get a better picture of T lymphocyte activation, PBMCs were analyzed for the generation of CD4+ and CD8+ T cells.
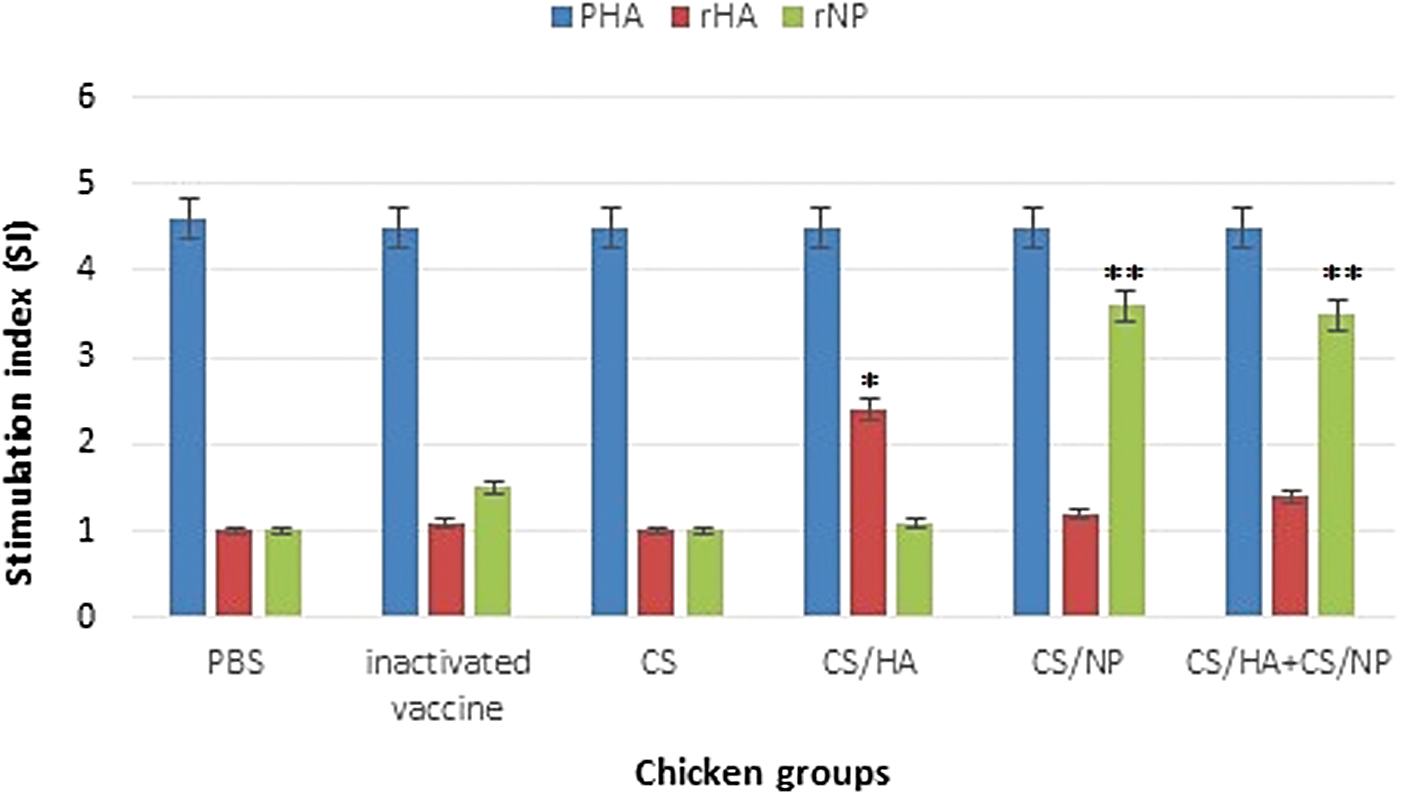
Specific T cell proliferation responses of peripheral blood mononuclear cells to influenza H9N2 protein-based nanoparticles. *Significant difference (p < 0.05) for rHA1, **Significant difference for rNP.
The results of this analysis, shown in Table 4, revealed that expansions of CD4+ and CD8+ cells were observed in all treated groups compared to chickens receiving PBS. No difference was observed in the percentages of CD4+ following vaccination with either protein-based nanoparticles or inactivated vaccine. However, a significant increase was observed in the expression of CD8+ with CS-NP compared to the CS-HA1 (20.48 ± 1.36 vs. 18.74 ± 1.89; p < 0.05). Vaccination with CS-HA1+CS-NP led to more increased abundance of CD4+ and CD8+ cells in chickens after the third vaccination. The CD4+/CD8+ ratios in the CS-HA1 alone and CS-HA1+CS-NP vaccinated groups were significantly higher (p < 0.05) than in the group inoculated with CS-NP. The differences in the percentage of CD8+T cells and the ratio of CD4+/CD8+ indicated that the inactivated vaccine induces only humoral responses.
T Lymphocyte Analysis from the Peripheral Blood Lymphocytes of Vaccinated Chickens with HA1- and Nucleoprotein-Based Nanoparticles
The data presented are means ± SD of results from three replicates.
Compared with chickens received CS-NP alone (p < 0.05).
Compared with chickens received CS-HA1 alone and inactivated vaccine (p < 0.05).
The most important feature of an effective influenza vaccine candidate is the capacity for protection against the infection. Challenge virus was selected on the basis of a comparison of the divergence of HA sequences from the cluster of latest circulating viruses thought to be diverse from the progenitor of Iranian H9N2 viruses. The virus shedding was detected in trachea of the challenged chickens by quantitative real-time PCR and samples with a Ct value ≤40 were considered positive for H9N2 virus. Due to the low pathogenicity feature of the H9N2 viruses, all groups showed no mortalities. The numbers of viral copies were reduced in the inactivated, CS-HA1, CS-NP, and CS-HA1+CS-NP-vaccinated groups compared to the negative controls. This number was significantly reduced (p < 0.05) in chickens immunized with CS-HA1+CS-NP and the inactivated vaccines.
As shown in Figure 6, vaccination with these vaccines led to a reduction in virus shedding at both time points compared to each of CS-HA1 and CS-NP. The virus shedding with Ct-values of 45.81 and 43.54 at 7 dpc revealed that CS-HA1+CS-NP and inactivated vaccines give an earlier viral clearance in the respiratory tract. For these groups, Ct-values at first and second sampling were significantly higher (p < 0.05) than in the CS-HA1 and CS-NP groups meaning that they conferred better protection against the virus challenge. Such differences in Ct-values signify the differences in virus load and its clearance rate between the challenged groups.

Means of viral Ct values in trachea of chickens vaccinated with influenza protein-based nanoparticles at 1st and 2nd weeks postchallenge. *Significant difference p < 0.05 with other groups, **Significant difference with PBS and CS groups. The red line indicates a positive Ct value for H9N2 influenza virus.
Both vaccines were highly protected against viral challenge in chickens, as shown by means of Ct values in the trachea. However, vaccination with CS-HA1+CS-NP induced higher reductions in viral load compared with the inactivated vaccine, especially at 7 dpc. Providing the best protection against the challenge virus by CS-HA1+CS-NP indicates that the conserved HA1 was antigenically matched closely enough with circulating viruses, which validate by the humoral response as a result of vaccination.
Discussion
The main goal in controlling influenza in the H9N2 endemic areas is the development of a more efficacious vaccine that could provide broad immunity to different isolates of the subtype. Most of influenza vaccines are produced from nonmatched strains, which may lead to diminished vaccine effectiveness due to positive selection pressure in HA. A key challenge for vaccine manufacturers is the genomic variability of influenza viruses. The persistence and continuous circulation of the viruses raise concern about the capacity of the marketed vaccines. The new strategies of influenza vaccine production have been focused on displaying one or more recombinant proteins in nanoparticles, reverse genetics, and mRNA platform to increase vaccine breadth and effectiveness (3,12,25,36).
Herein, we examined CS-based HA1 and NP nanoparticles' impact on immunity against H9N2 virus following mucosal vaccination. Each of the HA1 and NP constructs was loaded into the CS nanoparticles scaffold and used to evaluate the immunogenicity and protective capacity against H9N2 alone or in combination. We found that these nanoparticles were immunogenic in chickens and elicited HA- and NP-specific antibodies. Furthermore, the protein-based nanoparticles trigger cell-mediated immune responses in chickens.
The influenza viruses are highly variable due to genetic shift and drift, and therefore the vaccine formulation should be changed when new antigenic variants emerged. The influenza vaccine's effectiveness mainly depends on antigenic match between the vaccine strain and the circulated viruses (2,32). Among the vaccine production strategies, encapsulation of the viral antigens in nanoparticles to better stimulation of antibody production and T lymphocyte activation have been more studied. During influenza infection, the host immune system is stimulated to produce virus-specific neutralizing antibodies and also non-neutralizing antibodies that indirectly promote antibody-dependent cellular cytotoxicity (30). The neutralizing antibodies may become ineffective due to drifts and constant selective pressure on HA (2). Thus, the generation and expansion of CD8 T cell responses to provide long-term protection are urgent to vaccine design approaches.
It has been shown that NP antigen generates strong both antibody and cell-mediated immune responses. The anti-NP antibodies prolong antigen presentation to Fc receptor-bearing DCs and extend a CD8 response thereby a combination of anti-NP antibodies with CD8 T cells could be implicated in providing protection against influenza infection (29,33).
In light of the low immunogenicity of the recombinant antigens, the addition of adjuvants is required in increasing and directing immune responses against influenza viruses. The potential use of both natural and synthetic nanoparticles to promote the delivery and display of the targeted antigen to DCs and enhance its stability has been evaluated in influenza vaccine development. Zhao et al. (38) fused the spore coat protein of Bacillus subtilis with three copies of conserved influenza M2e protein for the development of a heat-stable and orally delivered vaccine. They reported that the viral protein was successfully displayed on the spore surface and M2e-specific IgG, as well as strong cellular immune responses, were induced in immunized mice.
In another similar study, oral immunization of mice with the recombinant spore carrying M2e led to producing relatively low antibody titers, which was not inadequate for protection (15). Virus-like particle (VLP)-based vaccine candidates could induce broad immune responses either as particulate carriers or immunopotentiators (12). Induction of potent humoral and cell-mediated immunity and promotion of cross-protection against different influenza subtypes were reported for VLP vaccine candidates. The biomimetic dual-antigen vaccines could elicit specific antibodies against these antigens, induce strong cell responses, and confer complete protection against the viruses (16,17,25).
Polysaccharides such as CS and its derivatives have immunomodulatory effects and able to activate the immune system and promote antigen-specific immune responses. The whole inactivated H9N2 virus and HA-based DNA vaccine encapsulated in CS nanoparticles induced efficient immune responses in immunized mice and chickens (4,9,10,13,19,37). According to the functions of HA and NP, we focused on producing recombinant proteins and nanoparticles to develop a more efficacious vaccine lead to broader protection. CS nanoparticles ionic cross-linked with TPP were used to deliver encapsulated HA1 and NP antigens. The average size, DPI, surface charge, and spherical structure of the synthesized nanoparticles showed high quality. These properties play roles in antigen cellular uptake.
Although the optimal size for mucosal vaccine delivery is still at issue, there is a consensus that nanoparticles sized 500 nm and below are more efficiently taken up by DCs than larger particles (1). In our study, the average sizes of CS-HA1 and CS-NP were estimated <400 nm. Nanoparticles with zeta potential in the range of greater than +25 mV or less than −25 mV are considered more stable due to the larger electrostatic repulsion interactions between the particles. The CS nanoparticles, and viral HA and NP exhibited a high positive surface charge. Interaction between the positive charge of nanoparticles and the negative charge of cell membranes enhances antigen uptaking and retention upon the mucosal surface. The better adhesion led to promote the presentation of the encapsulated antigen into DCs (1).
Consistent with the other studies, a reduction in zeta potential after encapsulation of the negatively charged antigens was seen compared with CS nanoparticles alone. Chicken groups were then used to assess the immunogenicity of the nanoparticles either alone or in combination. We found that each of HA1 and NP nanoparticles was immunogenic and elicited specific antibodies against H9N2 virus, but presence of NP is needed for induction of cell-mediated immune response. The CD4+ T cells have role in the generation and shaping of the immune response to influenza virus by providing B cell help for the production of high-affinity antibodies, and the expansion of CD8+ memory T cell subsets that contribute to boost the immune responses. CD8 T cell responses are frequently generated against more highly conserved proteins (30,33). Therefore, administration of NP-based vaccine could potentially provide long-term protection against heterologous strains.
The target set by the local Veterinary Organization in endemic areas is to produce at least one efficient vaccine providing immunological memory and broad-spectrum protection against circulating H9N2 variants. Most of the immunization studies following influenza vaccination are applied on mouse models, hence understanding the specificity of the adaptive immune responses in detail in poultry is limited. Adaptive responses include humoral and cell-mediated responses that are similar between birds and mammals in general terms; however, in many aspects, the avian immune system is simpler. This response is triggered following the presentation of antigen to T cells by DCs. As a result, CD4+ and CD+ cells become activated leading to the regulation of the effector cells (NK cells, heterophils, and T lymphocyte subsets) of humoral and cell-mediated immune responses and the establishment of immunological memory.
Cellular immunity plays an important role in limiting influenza virus replication and accelerating the clearance of virus-infected cells (19,37). To evaluate the potential effect of CS-HA1 and CS-NP nanoparticles on antigen-specific humoral responses to H9N2, serum antibodies in the vaccinated chicken were determined. The results suggested that a combination of these nanoparticles could increase and prolong specific antibody titers in chicken than the CS-HA1 or CS-NP alone. The finding is consistent with the antigen-stimulated lymphocyte proliferation results in the vaccinated chickens. The Challenge study indicates that the CS-HA1+CS-NP vaccine candidate can provide acceptable protection against H9N2 by reducing and shortening viral shedding, but complete prevention was not achieved.
The commercial H9N2 inactivated vaccines used in Iran are composed of the viruses isolated in 1998. The phylogenetic studies on H9N2 isolates have been revealed the genetic diversities in HA gene sequences, especially where the virus is endemic. Hence, the ability of such vaccines to inhibit viral replication and offer complete protection is questionable. Here, we have been addressed the immunogenic and protective effects of HA1- and NP-based nanoparticles. The particles accelerate the humoral immune response and elicit the cell-mediated immune response to H9N2. However, there is a concern about this approach that the CS-HA1+CS-NP cannot provide complete protection against influenza infection. Using other viral proteins such as M2e is one way to enhance broad protection. Antibodies against the conserved peptide are not neutralizing but contribute to the protection process by promoting effector functions based on their Fc region (22).
From the first favored protection against Marek's disease in chicken in 1973, efforts to develop more effective vaccines giving consideration to recombinant protein are continued. In comparison to the traditional vaccines, a recombinant protein generates overall lower immunogenicity. The vaccine candidate antigen needs adjuvants capable to trigger the cytokines signaling pathways and requires boosters to provide an adequate level of immune responses. The use of nanotechnology can overcome some of the limitations by triggering both Th1 and Th2 responses and produce quick and persistent immune responses to the influenza virus. However, difficulties in the codelivery of two or more antigens to the DCs need to be considered in the formulation vaccine.
Animal Welfare and Ethics Statement
The animal experiments were performed under protocols approved by the Animal Care and Use Committee at Razi Vaccine and Serum Research Institute (Approval
Footnotes
Authors' Contribution
S.S., M.M.E., and M.T. are from the Razi Vaccine and Serum Research Institute, Agricultural Research Education and Extension Organization (Karaj, Iran), and F.F. is from the Pasteur Institute of Iran (Tehran, Iran), all where the primary functions are education and research.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by Razi Vaccine and Serum Research Institute and Pasteur Institute of Iran under grant No. 3-18-1852-051-970773.