
Abstract
The world has been facing a pandemic for the past 2 years. COVID-19 still leads to millions of deaths worldwide, while deteriorating the global economy. The need for therapeutic targets, thus, remains. Interestingly, red blood cells, apart from gas exchange, also serve as modulators of innate and adaptive immunity. This function is accommodated mainly by surface molecules (proteins, lipids, and carbohydrates) and increased antioxidant capacity. However, under the circumstances of a disease state, red blood cells can become proinflammatory cells. Recent evidence has shown that, in the context of COVID-19, erythrocytes present protein oxidation, decreased antioxidant capacity, increased glycolysis, altered membrane lipidome, increased binding of Cytosine-Guanine (CpG) DNA and complement proteins, and low CD47 levels. These changes lead to an erythrocyte-dependent inflammation, which possibly participates in the hyperinflammation status of COVID-19. The current knowledge for the dysfunction of red blood cells during COVID-19 implies that the BAND3 protein and toll-like receptor 9 are potential therapeutic targets for COVID-19.
Introduction
COVID-19 is responsible for millions of deaths (World Health Organization, 2022) and economic costs (Seghieri et al, 2021) worldwide. Patients with severe disease experience hypoxia that precedes hyperinflammation (Grange et al, 2021; Grieb et al, 2021). Despite the development of effective vaccines, a significant percentage of individuals remain unvaccinated (Statista, 2021), whereas a percentage of fully vaccinated people can still suffer from the disease (Grange et al, 2021). This indicates the need for the discovery of novel therapeutic targets (Assaf et al, 2021).
Red blood cells represent the most numerous cell type in mammals, including humans. During the past decades their role has been revisited. It is now accepted that apart from gas exchange, they also function as important modulators of innate and adaptive immunity (Anderson et al, 2018; Papadopoulos, 2022a; Papadopoulos, 2022b; Papadopoulos et al, 2021a; Papadopoulos et al, 2021b), systemic metabolism of lipids (Papadopoulos et al, 2021a), amino acids (Thorn et al, 2020), and hormones (Papadopoulos et al, 2021c). There is also evidence that red blood cells can serve as vehicles for the transport of viruses in the circulation (McCullough, 2014) or even as mediators of pathogenic mechanisms of infectious diseases (Neidlinger et al, 2006).
Various studies have implicated red blood cells in the pathogenesis of COVID-19. In this review, we summarize the current knowledge for the contribution of red blood cells in COVID-19 (Fig. 1) and try to unveil their underestimated role as a therapeutic target.
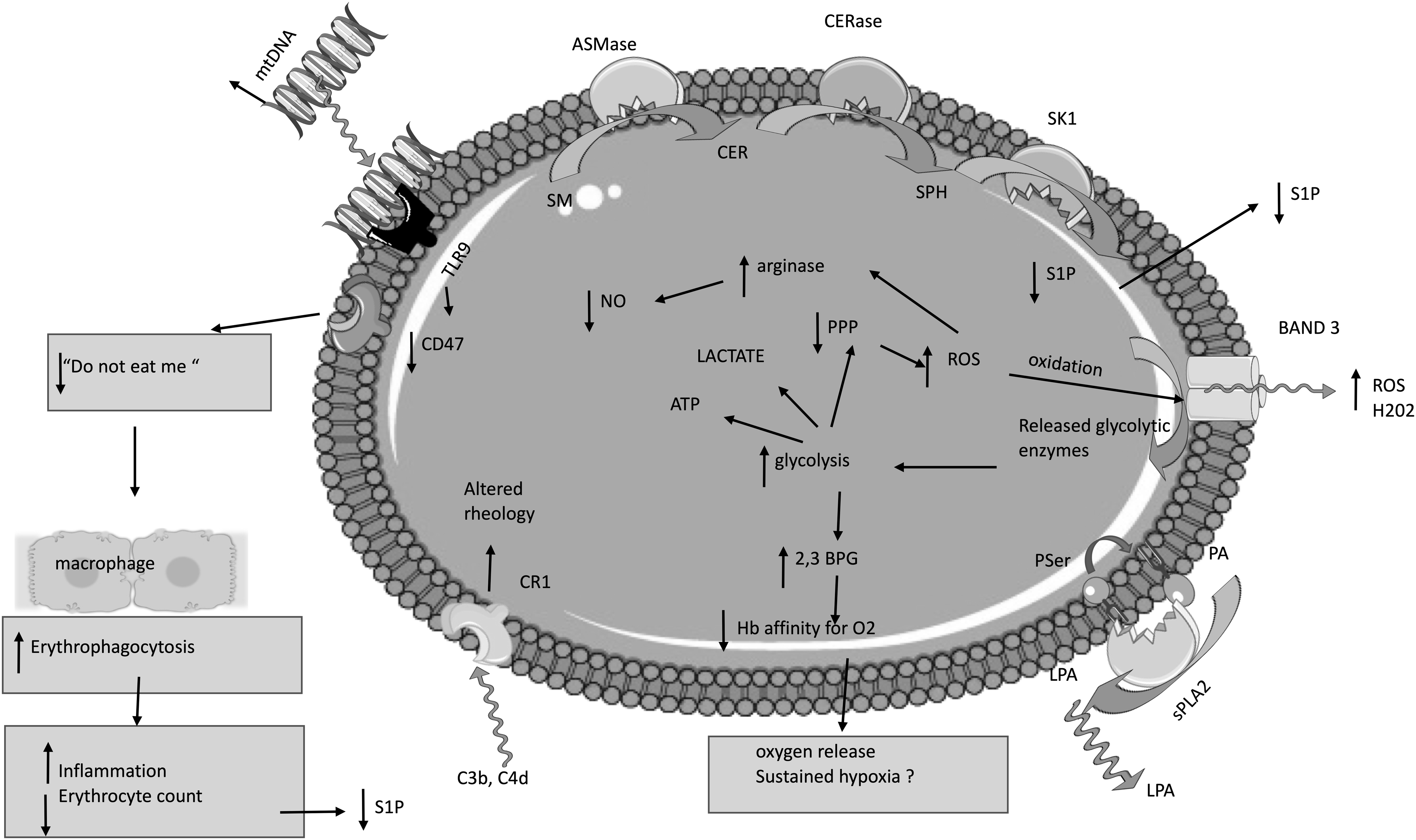
Molecular basis for erythrocyte dysfunction in COVID-19 patients. In erythrocytes of COVID-19 patients, oxidation of the BAND3 protein can lead to release of glycolytic enzymes. This step results in upregulation of glycolysis, leading to increased production of 2,3 BPG, promoting oxygen release. However, this step could lead to sustained hypoxia due to decreased oxygen affinity of hemoglobin. In addition, upregulation of glycolysis halts the pentose phosphate pathway, limiting in this way the reduction capacity of red blood cells. Thus, a positive feedback loop is created between the reduced antioxidant capacity and oxidized BAND3 protein. Next, ROS could upregulate arginase, limiting the substrate, arginine, for the production of NO. Damage of the lung epithelium leads to the release of mitochondrial DNA in the circulation that functions as a damage-associated molecular pattern. Mitochondrial DNA binds to the TLR9 of red blood cells, provoking conformational changes and loss of the CD47 protein, a “do not eat me” signal. Reduced levels of the CD47 protein result in increased erythrophagocytosis. This step enhances inflammation and leads to decline in the erythrocyte count. Since erythrocytes are the main source of S1P, erythrophagocytosis could also lead to reduced S1P in the circulation. However, this could also be the result of decreased S1P production by red blood cells. In addition, complement proteins bind to the CR1 of red blood cells, resulting in altered rheology. Finally, exposed phosphatidylserine on the erythrocyte surface can function as a platform for sPLA2, leading to the formation of LPA. 2,3-BPG, 2,3 biphosphoglycerate; ASMase, acid sphingomyelinase; BAND3, Band 3 anion transport protein; C3b, C4d, complement components 3b and 4d; CD47, cluster of differentiation 47; CER, ceramide; CERase, ceramidase; CR1, complement C3b/C4b receptor 1; LPA, lysophosphatidic acid; mtDNA, mitochondrial DNA; NO, nitric oxide; PA, phosphatidic acid; PPP, pentose phosphate pathway; PSer, phosphoserine; ROS, reactive oxygen species; S1P, sphingosine-1-phosphate; SK1, sphingosine kinase 1; SM, sphingomyelin; SPH, sphingosine; sPLA2, secretory phospholipase 2; TLR9, toll-like receptor 9.
COVID-19, Erythrocytes, and Glycolysis
The major evidence for erythrocyte malfunction during COVID-19 comes from the group of D'Alessandro (Thomas et al, 2020). In this study, the authors found increased glycolysis in erythrocytes of COVID-19 patients compared with healthy participants. Mechanistically, this can be associated with increased oxygen off-loading from hemoglobin, because of the 2,3-biphosphoglycerate synthesis through glycolysis, which lowers the hemoglobin affinity for oxygen (Sun et al, 2016). Augmented glycolysis can also be triggered by the oxidation of the protein BAND3 that was found in the same study (Thomas et al, 2020).
COVID-19, Erythrocytes, and Protein Oxidation
Indeed, oxidation of the protein BAND3 leads to the release of glycolytic enzymes in the cytosol of erythrocytes (Issaian et al, 2021). Increased glycolysis directs glucose toward adenosine triphosphate (ATP) and lactate production, while minimizing its use by the phosphate pentose pathway (Sun et al, 2016). This was, indeed, evident in the red blood cells of COVID-19 patients. However, the levels of reduced glutathione were unaltered. Nevertheless, increased levels of reactive oxygen species (ROS) in the red blood cells of COVID-19 patients could be provoked by the increased degradation of antioxidant enzymes, a finding observed in the study of Thomas et al (2020).
Consequently, increased ROS in red blood cells could lead to oxidation of membrane and cytoskeletal proteins of COVID-19 patients. Protein oxidation could also explain the findings of other studies, which noticed aberrant erythrocyte shape (Berzuini et al, 2021; Eta et al, 2021).
COVID-19, Erythrocyte, and Arginine Metabolism
In addition, previous studies have shown that increased ROS in red blood cells can upregulate the activity of arginase (Zhou et al, 2018). This could explain the increased arginase of COVID-19 erythrocytes (Thomas et al, 2020). Mortaz et al (2020) found increased intracellular nitric oxide (NO) in COVID-19 patients compared with healthy controls but did not find significant changes in the levels of NO in hypoxic non-COVID-19 patients compared with healthy controls. Conversely, Thomas et al (2020) did not find increased NO in erythrocytes of COVID-19 patients. However, we should consider that the study of Mortaz et al (2020) only included four healthy controls.
COVID-19, Erythrocytes, and Mitochondrial DNA
Reduced erythrocyte count in COVID-19 patients could be explained by the presence of toll-like receptor 9 (TLR9) in red blood cells, as Cytosine-Guanine (CpG) DNA binding to red blood cells provokes erythrophagocytosis. Lam et al (2021a) found increased CpG DNA in the circulation and bound on erythrocytes of COVID-19 patients. Mechanistically, mitochondrial DNA, which is secreted in the circulation as a signal of damage by the lung's epithelial cells, binds to the TLR9 of erythrocytes.
Consequently, this leads to reduced CD47 surface expression, as a result of alterations in the conformation of the protein CD47, due to physical interaction of CD47 with TLR9. Since CD47 functions as a “do not eat me” signal and inhibits erythrophagocytosis (Burger et al, 2012), reduced CD47 could augment erythrophagocytosis (Lam et al, 2021a). This step was found to promote inflammation and anemia.
COVID-19, Erythrocytes, and Complement Components
Lam et al (2021b) have also found increased disposition of C3, C3b, IC3b, and C3g+ on erythrocytes of COVID-19 patients. Binding of immune complexes to red blood cells could serve as a biomarker of tissue damage and organ injury.
COVID-19, Erythrocytes, and Lipid Metabolism
Thomas et al (2020) also evaluated the lipid profile of erythrocytes in COVID-19 and non-COVID-19 individuals. They found decreased phosphatidic acid (PA) and lysophosphatidic acid (LPA) levels on the membrane of red blood cells of COVID-19 positive patients. Since red blood cells exposing phosphatidylserine are targets of secretory phospholipase A2 (Neidlinger et al, 2006), and this enzyme is highly active in severe COVID-19 patients (Snider et al, 2021), we speculate that in the context of COVID-19, secretory phospholipase A2 leads to the synthesis and release of LPA. This could explain the lower levels of both PA and LPA that were found in COVID-19 patients.
Remarkable changes were also observed with regard to sphingolipids. Although sphingomyelin and ceramide levels remained unaltered, the total sphingolipid levels decreased in COVID-19 patients, meaning that the levels of sphingosine and sphingosine 1-phosphate (S1P) decrease. This hypothesis would be in line with the findings of Marfia et al (2021), who noticed decreased serum S1P levels in COVID-19 patients. However, Marfia et al (2021) correlated this change with reduced erythrocyte count, something that was not observed by Thomas et al (2020). In any case, since erythrocytes are the main source of S1P in the circulation, reduced erythrocyte-derived S1P could be related to compromised immune reaction.
Thomas et al (2020) noticed alterations in various lipids. It is of physiological relevance that they found increased levels of ω-3-derived local tissue-acting anti-inflammatory mediators, resolvins D1, D2, and E1. These lipid species could amplify erythrophagocytosis by macrophages.
COVID-19, Erythrocytes, and Vascular Inflammation in the Lung
In addition, increased ROS in erythrocytes have previously been implicated in hypoxic lung inflammation (Kiefmann et al, 2008). Specifically, hemoglobin binding to BAND3 has been shown to facilitate the release of ROS (mainly H2O2), provoking lung endothelial inflammation (Huertas et al, 2013). Indeed, Mahdi et al (2022) have found that erythrocytes from COVID-19 patients induce endothelial inflammation, through the production of ROS and compromised release of NO. Hence, we propose that a vicious cycle between hypoxia and erythrocyte dysfunction occurs, which augments lung inflammation.
Discussion
Based on the aforementioned studies, we speculate that molecules responsible for the red blood cell dysfunction could serve as therapeutic targets. Both BAND3 and TLR9 appear to regulate the cellular metabolism as well as the immune function of red blood cells. BAND3 oxidation could lead to inability of oxygen loading of erythrocytes, whereas CpG binding to TLR9 could result in anemia, reduced S1P levels, hypoxia, and inflammation.
It is of interest whether the changes in the erythrocyte function and structure during COVID-19 are directly triggered by the virus or comprise the result of the systemic alteration during COVID-19. Interestingly, Cosic et al (2020), through application of biophysical methods, hypothesized that BAND3 can also serve as an entry receptor for SARS-CoV-2 virus. However, because of the absence of experimental data, the question remains. We speculate that it is mainly the systemic alteration noticed in COVID-19 that impacts erythrocytes.
To sum up, erythrocyte dysfunction during COVID-19 could lead to aberrant immune function, through generation of ROS, reduced levels of S1P, and increased erythrophagocytosis. The current knowledge for the dysfunction of red blood cells during COVID-19 implies that the BAND3 protein and TLR9 are potential therapeutic targets for COVID-19.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.