
Abstract
Several inactivated SARS-CoV-2 vaccines have been approved for human use, but are not highly potent. In this study, different formulations of the inactivated SARS-CoV-2 virus were developed in Alum, Montanide 51VG, and Montanide ISA720VG adjuvants, followed by assessment of immune responses. The SARS-CoV-2 virus was inactivated with formalin and formulated in the adjuvants. BALB/c mice were immunized subcutaneously with 4 μg of vaccines on days 0 and 14; (IL-4) and (IFN-g), cytotoxic T lymphocyte (CTL) activity, and specific immunoglobulin G (IgG) titer and IgG1, IgG2a, and IgG2a/IgG1 ratio, and anti-receptor-binding domain (RBD) IgG response were assessed 2 weeks after the final immunization. Immunization with SARS-CoV-2-Montanide ISA51VG showed a significant increase in the IFN-γ cytokine versus SARS-CoV-2-Alum, SARS-CoV-2-Montanide ISA720VG, and control groups (p < 0.0033). Cytokine IL-4 response in SARS-CoV-2-Alum group showed a significant increase compared with SARS-CoV-2-Montanide ISA51VG, SARS-CoV-2-Montanide ISA720VG, and control groups (p < 0.0206). In addition, SARS-CoV-2-Montanide ISA51VG vaccine induced the highest IFN-γ/IL-4 cytokine ratio versus other groups (p < 0.0004). CTL activity in SARS-CoV-2-Montanide ISA51VG and SARS-CoV-2-Montanide ISA720VG groups showed a significant increase compared with SARS-CoV-2-Alum and control groups (p < 0.0075). Specific IgG titer in SARS-CoV-2-Montanide ISA51 VG and SARS-CoV-2-Montanide ISA720VG showed a significant increase compared with SARS-CoV-2-Alum and control groups (p < 0.0143). Results from specific IgG1and IgG2a in SARS-CoV-2-Alum, SARS-CoV-2-Montanide ISA51VG, and SARS-CoV-2-Montanide ISA720VG vaccine showed a significant increase compared with phosphate buffer saline (PBS) group (p < 0.0001), but SARS-CoV-2-Montanide ISA51VG and SARS-CoV-2-Montanide ISA 720VG groups showed the highest IgG2a/IgG1 ratio and a significant increase compared with SARS-CoV-2-Alum group (p < 0.0379). Moreover, inactivated SARS-CoV-2+Alum and SARS-CoV-2-Montanide ISA 720VG groups demonstrated a significant increase in anti-RBD IgG response versus the SARS-CoV-2-Montanide ISA51VG group. It seems that the type of vaccine formulation is a critical parameter, influencing the immunologic pattern and vaccine potency and human-compatible oil-based adjuvants were more potent than Alum adjuvant in the vaccine formulation.
Introduction
The COVID-19 pandemic started in December 2019 in Wuhan, China. The new SARS-CoV-2 is a positive-strand RNA virus with a genome of ∼30 kb. This virus is a member of Betacoronavirus genus, Nidovirales order, Coronaviridae Family, and Coronavirinae subfamily, which infects humans and mammal cells. The World Health Organization named the epidemic a Public Health Emergency of Global Importance in January 2020 and a pandemic in March 2020 (Jeyanathan et al, 2020). Vaccination is the most effective way to prevent viral infections and it seems that it is important to have a global vaccination to prevent the pandemic (Mahdavi et al, 2011).
The virus has relatively higher rate of transmission and infectivity, predominantly along the respiratory pathway. The most known infection route is droplet delivery, while aerosols can constitute another significant route (Ciotti et al, 2020). Therefore, the virus spread should be stopped as soon as possible (Pfefferbaum and North, 2020). In this light, different countries and governments have paid a high price to prevent the spread and treatment of this disease and develop an efficient vaccine (Wang et al, 2006). Genome analysis and comparison with previously released data showed that SARS-CoV-2 has distinct characteristics different from the other coronaviruses (Andersen et al, 2020; Nao et al, 2017).
It is clear that both humoral and cellular immune responses are required to combat and clear the viral infections. Therefore, these properties should be considered for successful vaccine development in both vaccine design and formulation (Jamali et al, 2007; Zhu et al, 2020). Different platforms of SARS-CoV-2 vaccines, including mRNA vaccine, DNA vaccine, viral vectors, recombinant vaccine, and inactivated vaccines, have been developed, some of which were commercialized in different countries (Frederiksen et al, 2020; Jafari et al, 2022). Formulation of a vaccine in an adjuvant is important and influences vaccine potency.
In addition, formulation of a vaccine influences the vaccine stability and the type of immune responses triggered after immunization (Amini et al, 2017; Fakharzadeh et al, 2013; Mojarab et al, 2020; Najminejad et al, 2019; Rostami et al, 2017). One of the commercialized vaccines used for COVID-19 is inactivated viral vaccines, which is being now applied in different countries. The immunogenicity and potency of inactivated virus are low, especially for induction of T cell response, and making a change in the vaccine formulation may result in achievement to a more potent vaccine (Hotez et al, 2020; Logunov et al, 2020; Wang et al, 2020).
In this study, we developed different formulations of inactivated SARS-CoV-2 vaccines with different classes of adjuvants, followed by assessment of cellular and humoral immune responses in experimental mice. The results showed that different formulations of vaccine triggered different levels and platforms of immune responses.
Materials and Methods
Virus isolation and propagation
SARS-CoV-2 virus was isolated from throat/nasal swab sample derived from a patient (positive in real-time polymerase chain reaction [PCR]) from Karaj, Iran (SARS-CoV-2/human/IRN/Alborz-IR/2020 surface glycoprotein (S) gene, complete cds 3,822 bp linear RNA MW709393.1 GI: 1998262910). The virus replication was performed on Vero cell line cultured in Dulbecco's modified Eagle's medium supplemented with 10% heat-inactivated fetal bovine serum (FBS) and then confirmed by real-time PCR and gene sequencing (MW709393). For efficient growth in Vero cells, the virus strain was purified by the plaque assay.
The first purified clone was passaged three times on Vero cells to obtain an efficient virus stock. Large-scale virus production was performed on Vero cells cultured in 175 cm2 flasks with a density of more than 90% and MOI 0.1. After 48 h, when 85–90% Cytopathic effect was observed, all flasks were frozen at −20°C and then harvested. The harvested viruses were first clarified by centrifugation and then passed through a 0.2 filter. Finally, the obtained titration was found to be equal to 7 TCID50/mL (Kaffashi et al, 2021). The SARS-CoV-2 propagation was carried out in biosafety level-III facilities.
Virus inactivation and quantification
The SARS-CoV-2 strain was cultivated in the Vero cell line for propagation, and the supernatant of infected cells was inactivated with formalin 0.04% vol/vol at RT for 30 h (finally 4:10,000). Following clarification of cell debris, concentration was performed by ultrafiltration and 8% PEG-6000. Purification was performed using a column chromatography (Kaffashi et al, 2021). The purified and inactivated viruses were dialyzed versus phosphate buffer saline (PBS), passed through 0.22 filter, and finally quantified using the Bradford method. The quantified virus aliquots were stored at −70°C until use.
Vaccine formulation
Inactivated SARS-CoV-2 viruses were formulated in Montanide ISA 720VG, Montanide ISA51VG (SEPPIC, France), and/or Alum hydroxide (Pasteur Institute of Iran) adjuvants. To formulate in oil-based adjuvants, inactivated SARS-CoV-2 virus in PBS buffer was mixed with Montanide ISA 720VG (at a ratio of 30/70) and Montanide ISA 51VG (at a ratio of 50/50) by vigorously shaking to develop a milky white suspension, and homogenized using homogenizer to develop a homogen suspension.
To formulate the Alum-based vaccine, 4 μg of the inactivated SARS-CoV-2 virus in PBS buffer was mixed with 200 μg of Alum hydroxide adjuvant (for each dose of vaccine formulation) and shaken at 100 RPM for 60 min at room temperature and allowed the adsorption of viral particles on the surface of Alum gel (Savoji et al, 2021). All vaccine formulations were performed in the clean room of Department of FmD Vaccine formulation of Razi Vaccine and Serum Institute of Iran (Karaj, Iran). After final vaccine formulation, each 200 μL of all vaccines contained 4 μg of inactivated SARS-CoV-2 virus used for immunization.
Mice and immunization
Six-to 8-week-old male BALB/c mice (n = 40) were purchased from Royan Institute of Iran (Tehran, Iran). The mice were housed for 1 week before the experiments, given the food and water ad libitum, and maintained in a 12-h light/12-h dark cycles. The experimental mice were handled by an expert technician and in accordance with the Animal Care and Use Protocol of Razi Vaccine and Serum Research Institute of Iran. Experimental mice were randomly divided into four groups, each consisting of 10 mice as below:
The mice were immunized subcutaneously two times with 14-day intervals, and immunological parameters were assessed 2 weeks after the final immunization.
In vitro antigen recall of spleen cells
Two weeks after the second immunization, the spleen of immunized mice was removed under sterile conditions and mechanically dissected and suspended in sterile cold PBS containing penicillin and streptomycin. The cell suspension was centrifuged at 300 g/5 min and RBCs were lysed using lysis buffer, and single-cell suspension was washed three times and adjusted to 3 × 106 cells/mL in RPMI-1640 (Gibco, Germany) supplemented with 10% FBS, 4 mM L-glutamine, 1 mM sodium pyruvate, 100 μg/mL streptomycin, and 100 IU/mL penicillin.
One milliliter of cell suspension containing a total number of 3 × 106 spleen cells was seeded in each well of 24-well plates. The spleen cells were then stimulated in vitro by adding 1 μg/mL of inactivated SARS-CoV-2 virus and the plates incubated at 37°C in 5% CO2. After 48 h postantigen recall, the culture supernatant was centrifuged at 5,000 RPM/10 min and the supernatants were harvested and stored at −70°C for cytokine assay (Savoji et al, 2021).
Interferon-γ and interleukin-4 enzyme-linked immunosorbent assay
The quantity of interferon (IFN)-γ and interleukin (IL)-4 cytokines of supernatants, derived from in vitro antigen recalled culture, was estimated by quantitative mouse IFN-γ and IL-4 cytokine enzyme-linked immunosorbent assay (ELISA) Kits (Mabtech, Stockholm, Sweden) according to the manufacturer's instruction (Fathi et al, 2021). ELISA for IFN-γ and IL-4 cytokines was performed in the presence of cytokine standards and using the standard curve of each cytokine. The pg/mL of each sample was reported and for calculating the IFN-γ/IL-4 ratio, the quantity of IFN-γ of each mouse was divided into that one of IL-4 cytokine.
Cytotoxic T lymphocyte assay
The cytotoxic T lymphocyte (CTL) activity of experimental groups was assessed by Granzyme B (Gr-B) release. Briefly, 1.5 × 106 spleen cells of experimental mice were cultured in a 96-well plate and stimulated with 1 μg/mL of inactivated SARS-CoV-2 virus. As a negative control for each mouse, some wells were cultured without antigen stimulation. The plates were incubated at 37°C in 5% CO2, and culture supernatants were harvested after 48 h of antigen stimulation and used for Gr-B assay by ELISA according to the manufacturer's instruction (eBioscience). The quantity of Gr-B of stimulated wells was subtracted with unstimulated one of each mouse and used as net Gr-B release as a criterion of CTL activity (Mahdavi et al, 2017b).
ELISA of specific Immunoglobulin G antibody titer and their isotypes
Specific total Immunoglobulin G (IgG) antibodies were determined by an optimized indirect ELISA. Briefly, 100 μL of inactivated SARS-CoV-2 in PBS (5 μg/mL) was added into 96-well ELISA Maxisorp plates (Greiner, Germany) and incubated overnight at 4°C. The wells washed with PBS containing 0.1% Tween 20 (washing buffer) and blocked 1 h at 37°C with 2% skimmed milk in PBS +0.1% Tween 20 (blocking buffer). The plates were washed with washing buffer and 100 μL of 1/25 diluted sera up to 16 serial dilutions was added into each well and incubated at 37°C for 90 min.
The wells were washed five times with washing buffer and incubated for 2 h with 100 μL of 1/8,000 dilution of anti-mouse conjugated to horseradish peroxidase (HRP) (Razirad, Iran). The wells were washed five times and incubated 30 min with 100 μL of TMB substrate in the dark, the reaction was stopped with 100 μL of 2N H2SO4, and color density was measured at 450 nm with ELISA plate reader. The cutoff of ELISA for each titer was calculated on 10 serum samples of sham group by the following formula: mean + 3standard deviation (SD). Then, the optical density (OD) value of the last serum dilution which was above than cutoff value was considered as the endpoint titer of each individual mouse.
The final results of antibody titer for each mouse presented as Log10 of antibody titer (Khorasani et al, 2016). In addition, detection of specific IgG1 and IgG2a subclasses was carried out on the 1/25 serum dilution of experimental serums using goat anti-mouse IgG1 and IgG2a secondary antibodies (Sigma) according to the manufacturer's instruction.
Assessment of specific IgG anti- receptor-binding domain protein
The specific IgG antibody response against receptor-binding domain (RBD) protein was determined by a laboratory-optimized indirect ELISA. At the first for antigen coating, 100 μL of 2 μg/mL of RBD protein (The Native Antigen Company, UK) in carbonate-bicarbonate buffer with pH 9.6 was added into 96-well ELISA Maxisorp plates (Greiner, Germany) and incubated overnight at 4°C. The wells were washed with PBS containing 0.1% tween 20 (washing buffer) and then blocked 1 h at 37°C with 1.5% BSA in PBS +0.05% tween 20 (blocking buffer). The plates were washed five times with washing buffer and 100 μL of 1/50 diluted sera of experimental vaccinated and sham group of mice was added into each well and incubated at 37°C for 90 min.
The wells were washed five times with washing buffer and incubated for 90 min with 100 μL of 1/8,000 dilution of anti-mouse HRP conjugate (Razirad, Iran). The wells were washed five times and incubated for 10 min with 100 μL of TMB substrate in the dark, the reaction was stopped by adding 100 μL of 2N HCL, and color density was measured at 450 nm with ELISA plate reader. The cutoff of RBD-ELISA was calculated on the serum samples of the sham group by the following formula: mean + 3SD, and then IgG response to RBD for individual mouse was reported by the formula, OD of RBD ELISA of individual mouse/cutoff.
Statistical analysis
The data are presented as mean ± SD of each experiment. The statistical significance of differences was analyzed using two-way analysis of variance (ANOVA) followed by Tukey test (Graph Pad Prism 6.01 software, Graph Pad Software, La Jolla, CA). Statistical analysis of antibody titters and IgG isotypes was carried out by the Mann-Whitney U test. In all the cases, p-values <0.05 were considered significantly different.
Results
IFN-γ cytokine
Results from the IFN-γ cytokine in the experimental groups showed that only the injection of SARS-CoV-2 vaccine formulated in Montanide ISA51VG adjuvant resulted in a significant difference, compared with the control group (p = 0.0003). In addition, SARS-CoV-2-Montanide ISA51VG showed a significant increase in the IFN-γ response, compared with SARS-CoV-2-Alum and SARS-CoV-2-Montanide ISA 720VG groups (p = 0.0004 and p = 0.0033, respectively) (Fig. 1).
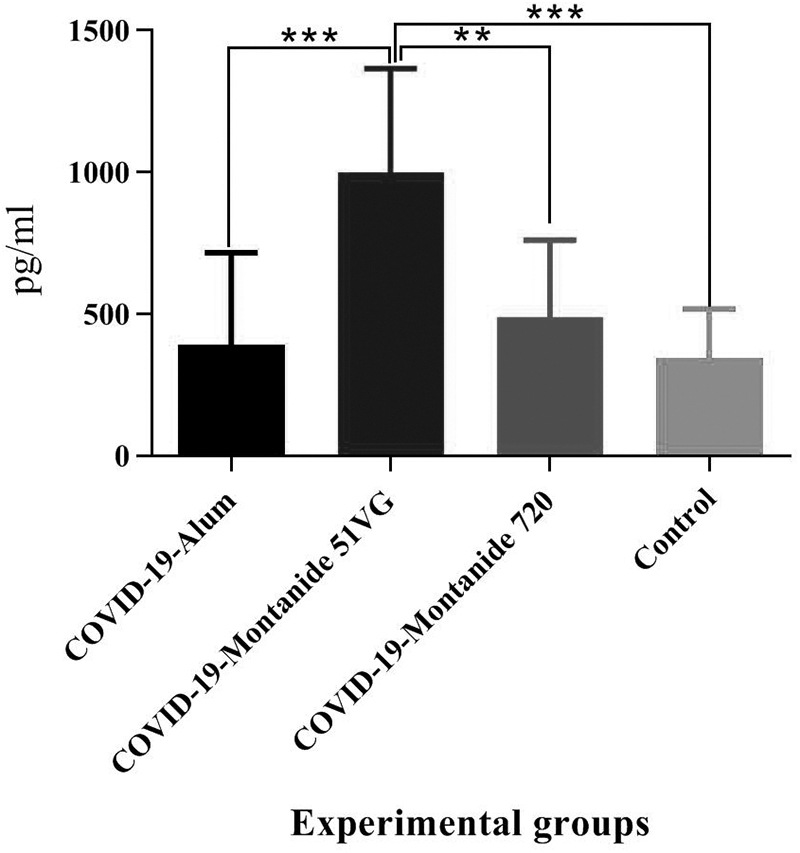
Results from the IFN-γ cytokine in the experimental groups. Injection of SARS-CoV-2-Montanide ISA51VG resulted in a significant difference, versus the PBS group (***p = 0.0003). In addition, SARS-CoV-2-Montanide ISA51VG showed a significant increase in the IFN-γ response, as compared with SARS-CoV-2-Alum and SARS-CoV-2-Montanide ISA 720VG groups (***p = 0.0004 and **p = 0.0033, respectively). IFN-γ, interferon-γ; PBS, phosphate buffer saline.
IL-4 cytokine
Immunization with SARS-CoV-2-Alum and SARS-CoV-2-Montanide ISA 51VG showed a significant increase in IL-4 cytokine secretion, compared with the control group (p = 0.0001 and p = 0.0288, respectively). In addition, mice immunized with SARS-CoV-2-Alum vaccine showed a significant increase compared with those immunized with SARS-CoV-2-Montanide ISA 51VG and SARS-CoV-2-Montanide ISA 720VG vaccines (p = 0.0206 and p = 0.0005, respectively) (Fig. 2).
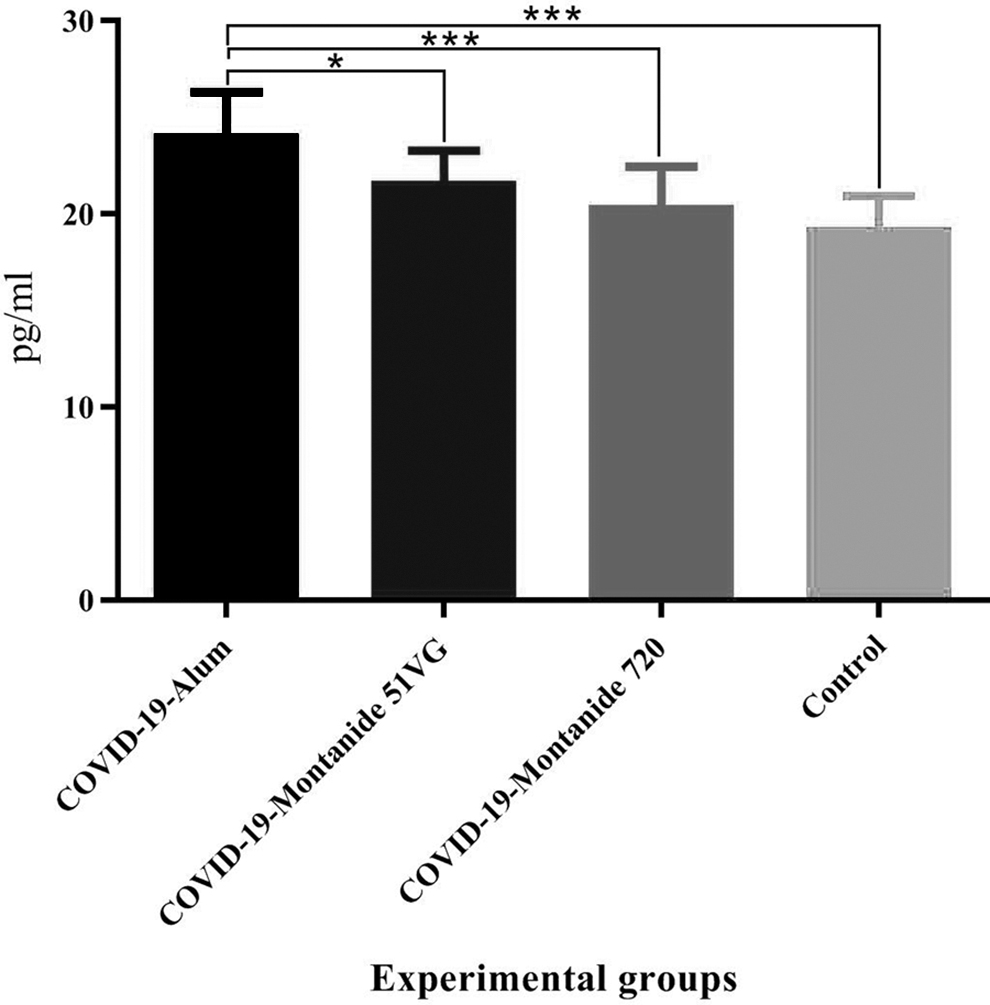
Results from IL-4 cytokine in the vaccinated mice. Immunization with SARS-CoV-2-Alum and SARS-CoV-2-Montanide ISA-51VG revealed a significant increase in the IL-4 cytokine response versus the control group (***p = 0.0001 and *p = 0.0288, respectively). Furthermore, the SARS-CoV-2-Alum vaccine showed a significant increase versus SARS-CoV-2-Montanide ISA-51VG and SARS-CoV-2-Montanide ISA-720VG vaccines (*p < 0.0206). IL-4, interleukin-4.
IFN-γ/IL-4 cytokine ratio
Results from the IFN-γ/IL-4 cytokine ratio in SARS-CoV-2-Montanide ISA51VG group revealed the highest IFN-γ/IL-4 ratio, which showed significant differences compared with the SARS-CoV-2-Alum, SARS-CoV-2-Montanide ISA 720VG, and PBS group (p = 0.0001, p = 0.0004, and p = 0.0001, respectively) (Fig. 3).

IFN-γ/IL-4 cytokine ratio in the mice groups. Immunization with SARS-CoV-2-Montanide ISA-51VG vaccine showed a significant increase in IFN-γ/IL-4 ratio versus SARS-CoV-2-Alum, SARS-CoV-2-Montanide ISA-720VG, and control group (***p < 0.0004).
CTL response
Results from CTL activity based on Gr-B secretion in mice immunized with SARS-CoV-2-Montanide ISA51VG, SARS-CoV-2-Alum, and SARS-CoV-2-Montanide ISA 720VG vaccines showed a significant increase compared with the control group (p = 0.0001, p = 0.0017, and p = 0.0001, respectively). In addition, SARS-CoV-2-Montanide ISA51VG and SARS-CoV-2-Montanide ISA 720VG vaccines showed a significant increase compared with the SARS-CoV-2-Alum group (p = 0.0001 and p = 0.0075, respectively). No significant difference was found between SARS-CoV-2 Montanide ISA51VG and SARS-CoV-2 Montanide ISA720VG group (p = 0.2628) (Fig. 4).

Results from CTL activity in the study groups. Immunization with SARS-CoV-2-Montanide ISA-51VG, SARS-CoV-2-Alum, and SARS-CoV-2-Montanide ISA-720VG vaccines revealed a significant increase versus control group (**p < 0.0017). In addition, SARS-CoV-2-Montanide ISA51VG and SARS-CoV-2-Montanide ISA 720VG vaccines showed a significant increase versus the SARS-CoV-2-Alum group (***p < 0.0075). CTL, cytotoxic T lymphocyte.
Specific IgG titer
Results from specific IgG titer in SARS-CoV-2-Montanide ISA51VG, SARS-CoV-2-Alum, and SARS-CoV-2-Montanide ISA 720VG groups showed a significant increase compared with the control group (p < 0.0001). Furthermore, SARS-CoV-2-Montanide ISA51VG and SARS-CoV-2-Montanide ISA 720VG vaccines revealed a significant increase in the IgG titer when compared with the SARS-CoV-2-Alum group (p = 0.0143 and p = 0.0048, respectively). There was no significant difference between SARS-CoV-2-Montanide ISA51VG and SARS-CoV-2-Montanide ISA720VG groups (p = 0.6867) (Fig. 5).
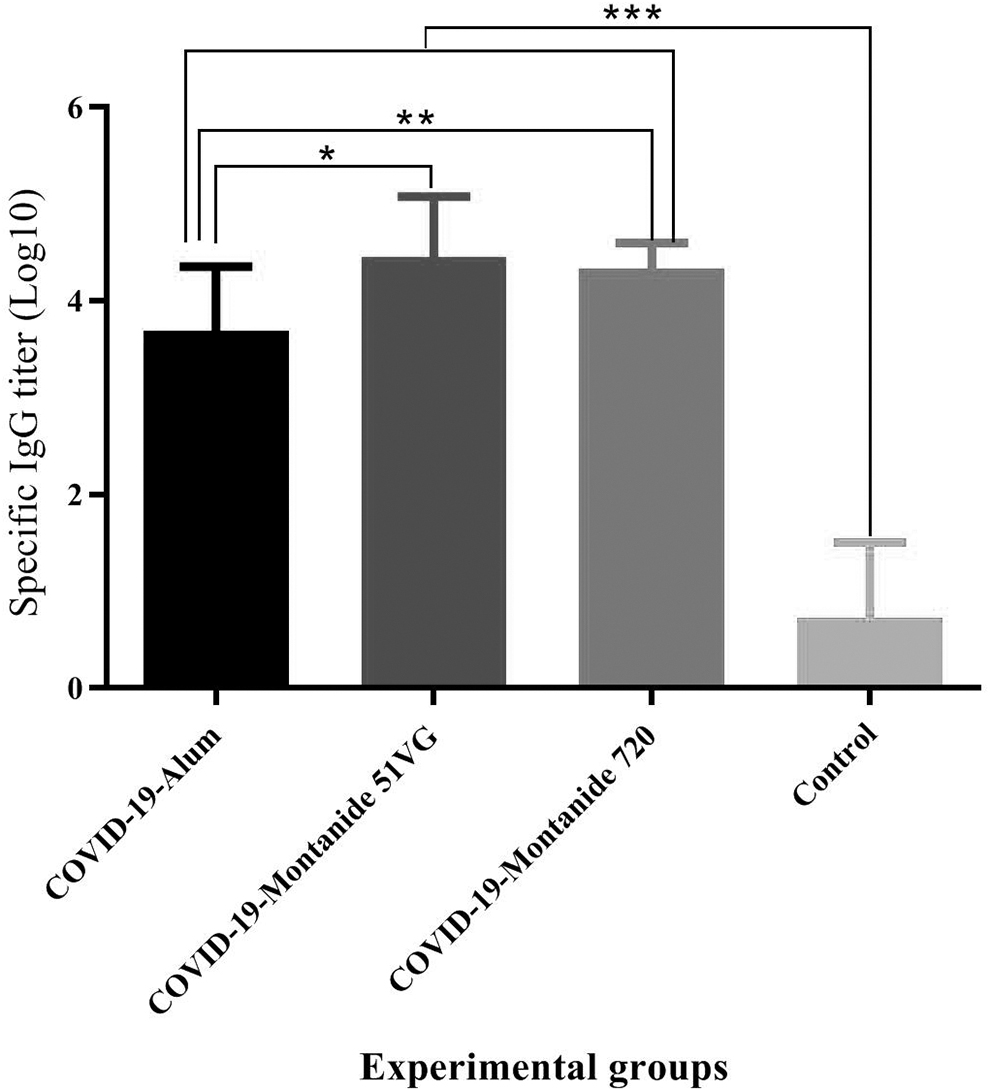
Results from specific IgG titer in the mice groups. Immunization with SARS-CoV-2-Montanide ISA51VG, SARS-CoV-2-Alum and SARS-CoV-2-Montanide ISA 720VG groups showed a significant increase versus the PBS group (***p < 0.0001). Furthermore, SARS-CoV-2-Montanide ISA51VG and SARS-CoV-2-Montanide ISA 720VG vaccines revealed a significant increase as compared with the SARS-CoV-2-Alum group (*p = 0.0143 and **p = 0.0048, respectively). There was no significant difference between SARS-CoV-2-Montanide ISA51VG and SARS-CoV-2-Montanide ISA720VG groups (p = 0.6867).
Specific IgG1 antibody
Specific IgG1 isotype in SARS-CoV-2-Montanide ISA51VG, SARS-CoV-2-Alum, and SARS-CoV-2-Montanide ISA 720VG vaccines showed a significant increase compared with the control group (p < 0.0001). Furthermore, SARS-CoV-2-Alum group revealed a significant increase in the IgG1 isotype compared with the SARS-CoV-2-Montanide ISA51VG and SARS-CoV-2-Montanide ISA 720VG vaccines (p = 0.0043 and p = 0.0041, respectively) (Fig. 6).

Specific IgG1 isotype antibody in the experimental mice. Immunization with SARS-CoV-2-Montanide ISA-51VG, SARS-CoV-2-Alum, and SARS-CoV-2-Montanide ISA-720VG vaccines showed a significant increase versus the control group (***p < 0.0001). In addition, SARS-CoV-2-Alum group revealed a significant increase in IgG1 isotype response versus SARS-CoV-2-Montanide ISA-51VG and SARS-CoV-2-Montanide ISA-720VG vaccines (**p < 0.0043).
Specific IgG2a antibody
Results from specific IgG2a isotype antibody in SARS-CoV-2-Montanide ISA51VG, SARS-CoV-2-Alum, and SARS-CoV-2-Montanide ISA 720VG vaccines showed a slight increase compared with the control group and no significant difference among the experimental groups (p > 0.1550) (Fig. 7).
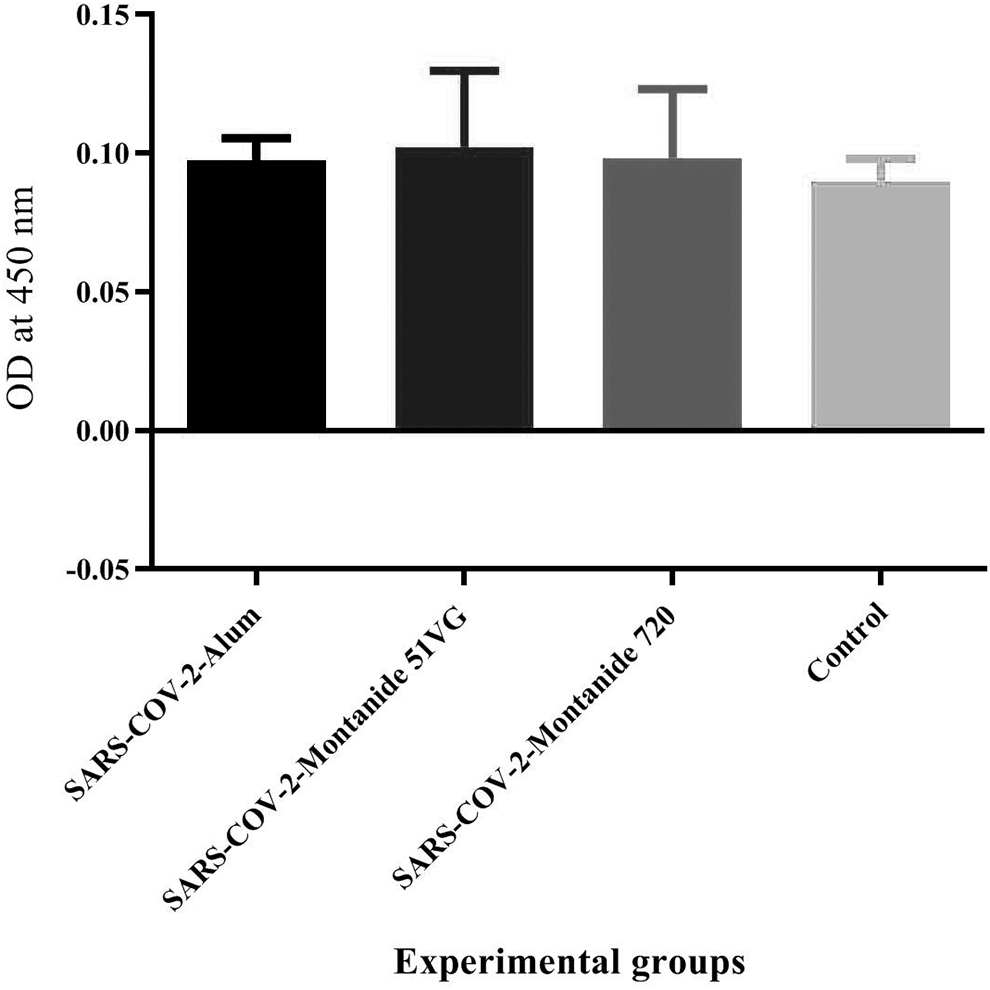
Results from specific IgG2a isotype antibody in the vaccinated groups. Immunization with SARS-CoV-2-Montanide ISA51VG, SARS-CoV-2-Alum, and SARS-CoV-2-Montanide ISA 720VG vaccines showed a few increases versus the control group (p > 0.1550).
IgG2a/IgG1 ratio
Results from the IgG2a/IgG1 ratio in SARS-CoV-2-Montanide ISA51VG and SARS-CoV-2-Montanide ISA 720VG vaccines resulted in a significant increase versus the SARS-CoV-2-Alum group (p = 0.0379 and p = 0.0175, respectively). However, there was no significant difference between SARS-CoV-2-Montanide ISA51VG and SARS-CoV-2-Montanide ISA720VG groups (p = 0.4458) (Fig. 8).
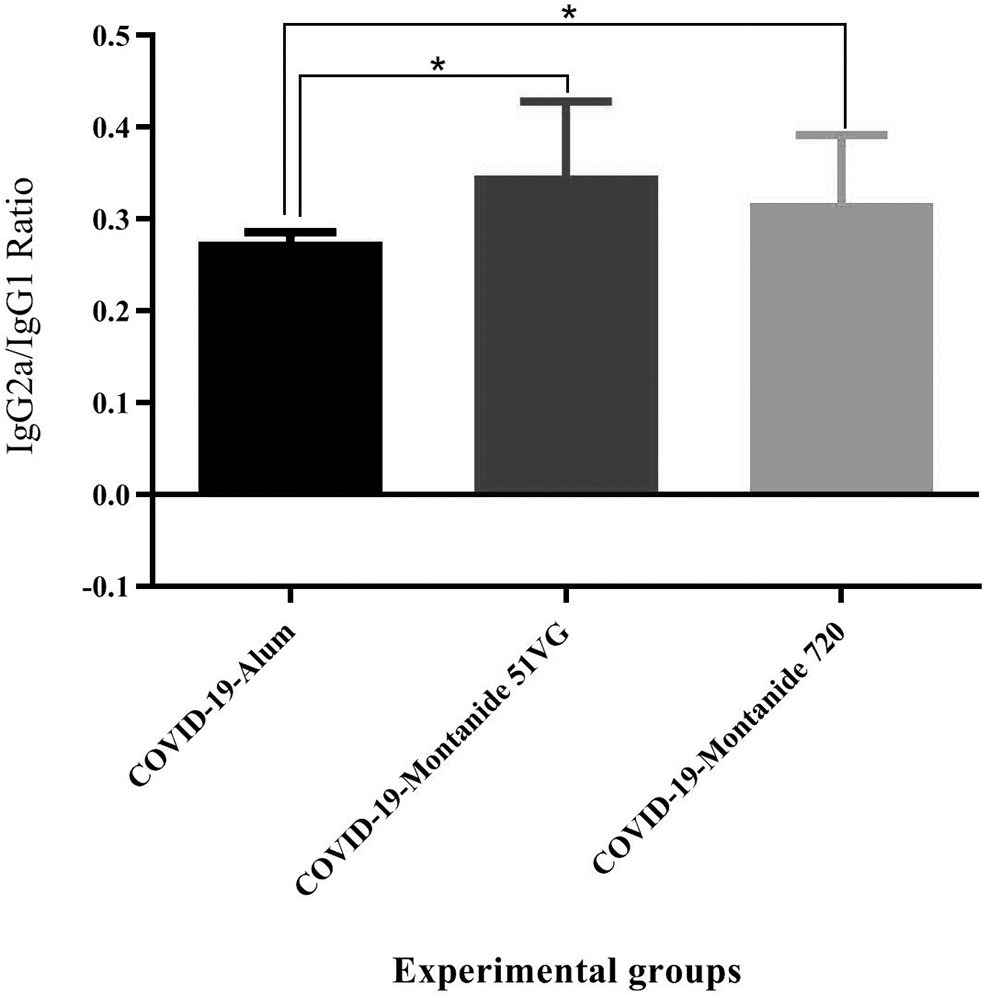
Results from the IgG2a/IgG1 ratio in the experimental groups. Immunization with SARS-CoV-2-Montanide ISA-51VG and SARS-CoV-2-Montanide ISA-720VG vaccines resulted in a significant increase in the IgG2a/IgG1 ratio versus the SARS-CoV-2-Alum group (*p < 0.0379). No significant difference in IgG2a/IgG1 ratio was found between SARS-CoV-2 Montanide ISA51VG and SARS-CoV-2 Montanide ISA720VG group (p = 0.4458).
Specific IgG against RBD protein
Results from anti-RBD IgG response in SARS-CoV-2-Montanide ISA51VG, SARS-CoV-2-Alum, and SARS-CoV-2-Montanide ISA 720VG vaccines revealed a significant increase versus the control group (p < 0.0185). In addition, inactivated SARS-CoV-2+Alum and SARS-CoV-2-Montanide ISA 720VG vaccines showed a significant increase versus the SARS-CoV-2-Montanide ISA51VG group (p < 0.0001), while SARS-CoV-2-Alum and SARS-CoV-2-Montanide ISA 720VG groups showed a comparable IgG response versus RBD protein and did not show a significant difference versus each other (p = 0.9999) (Fig. 9).

Specific IgG against RBD protein reported as OD of RBD ELISA/cutoff for each individual mouse. Immunization with SARS-CoV-2-Montanide ISA51VG, SARS-CoV-2-Alum and SARS-CoV-2-Montanide ISA 720VG vaccines revealed a significant increase versus the PBS group (*p = 0.0185, ***p < 0.0001, and ***p < 0.0001, respectively). In addition, inactivated SARS-CoV-2+Alum and SARS-CoV-2-Montanide ISA 720VG vaccines showed a significant increase versus SARS-CoV-2-Montanide ISA51VG group (***p < 0.0001 and ***p < 0.0001, respectively). But, immunization with SARS-CoV-2-Alum did not show a significant difference versus SARS-CoV-2-Montanide ISA 720VG group (p = 0.9999). ELISA, enzyme-linked immunosorbent assay; OD, optical density; RBD, receptor-binding domain.
Discussion
With the emerging pandemic of COVID-19 and high mortality and fast dissemination, much effort was put into the development an efficient vaccine (Bos et al, 2020). Finally, several vaccines for COVID-19 have been developed by a variety of organizations and academic centers, ranging from the inactivated virus and protein-based vaccines to DNA and mRNA-based vaccines with high efficacy in human clinical trials. One important issue regarding the vaccine potency is the type of adjuvant in the vaccine formulation and its relation to the immunologic patterns and vaccine potency (Crommelin et al, 2021; Peng et al, 2020; Wang et al, 2020).
Herein, we have developed three different inactivated SARS-CoV-2 vaccine formulations in human compatible adjuvants and focused on the nature of immune responses, which was triggered by each formulation of vaccine to show the critical role of adjuvant and also vaccine formulation on the immune platforms of SARS-CoV-2 vaccine. Result from the IFN-γ cytokine in the inactivated SARS-CoV-2-Montanide ISA-51VG group revealed the highest response compared with the Aslum- and Montanide ISA-720VG-based vaccines.
Several studies showed the potency of Montanide ISA-51VG adjuvant on the polarization toward a strong IFN-γ and Th1 response (Atsmon et al, 2012; Savoji et al, 2021; Soema et al, 2015), as we achieved in SARS-CoV-2 vaccine model. The role of IFN-γ cytokine in controlling viral infections is well known (Noh, 2020). Infected patients with SARS-CoV-2 had a complex trend of IFN-α, medium levels of IFN-γ, and undetectable IFN-β and λ cytokines (Trouillet-Assant et al, 2020). Several studies demonstrated that vaccines capable of inducing higher levels of the IFN-γ cytokine were more protective (Bos et al, 2020; Ramasamy et al, 2021; Rasmussen et al, 2021).
Cytokine IL-4 response showed a strong and dramatic response in Alum-based vaccine and even higher than the oil-based vaccines. The potency of Alum-based vaccines in the induction of IL-4 cytokine response is well known and confirmed the Th2 response pattern (Bungener et al, 2008; Serre et al, 2010). On the other hand, formulation of inactivated SARS-CoV-2 virus in oil-based adjuvants suppressed IL-4 cytokine response, confirming the Th1 pattern in Montanide adjuvanted groups. In addition, the IFN-γ/IL-4 cytokine ratio representing the Th1 cytokine network (de la Cruz et al, 2019; Maspi et al, 2017) has increased significantly in Montanide ISA51VG adjuvanted group versus Alum and Montanide ISA720VG-formulated vaccines. This finding is another conformance for Th1 polarization in Montanide ISA51VG adjuvanted vaccine group.
Results from Gr-B release as a criterion of the CTL activity in inactivated SARS-CoV-2-Montanide ISA-51VG group showed the highest Gr-B release, highlighting the highest CTL activity among the experimental groups. Gr-B triggers a cascade of events within the infected target cells that lead to induction of apoptosis and cell death (Lord et al, 2003), as well as clearance of viral infection from the body (Doherty et al, 1997). Evidences showed that early decrease in TCD8+ population is an important factor in the pathogenesis of COVID-19 infection, and improvement of TCD8+ lymphocyte response in the vaccination would be an important mechanism for resistance against the infection (Elsaesser et al, 2009; Ganji et al, 2020; Zheng et al, 2020). In this study, oil-based adjuvants were more successful than Alum-based vaccine in the induction of CTL response against SARS-CoV-2 antigens.
Results from IgG responses showed that SARS-CoV-2-Montanide ISA-51VG and SARS-CoV-2-Montanide ISA-720VG vaccines were more potent than SARS-CoV-2-Alum group. This property of oil-based vaccines is so critical because the protection in COVID-19 infection relies mainly on the specific antibodies; however, T cell response is important, too (Carrillo et al, 2021; Juno et al, 2020). Various studies have shown that oil-based adjuvanted vaccines are more potent in the induction of antibody responses and even T cell responses than Alum-based vaccines (Khorasani et al, 2016; Savoji et al, 2019, 2021).
More robust humoral immune responses in oil-based adjuvanted SARS-CoV-2 vaccines may be due to the nature of oil-based adjuvants, because Montanide family adjuvants are able to protect the antigen from denaturation by protease enzymes and free radicals at the injection site. These agents, through denaturation the structure of protein, decrease the vaccine potency. In addition, several studies have shown the potency of Montanide adjuvant family in the induction of follicular helper T cell that has a critical role in the differentiation of plasma cells and boosting humoral immune responses (Aucouturier et al, 2002; Riteau et al, 2016; Savoji et al, 2021).
In addition, IgG isotype analysis showed that inactivated SARS-CoV-2-Alum triggered highest IgG1 response, while immunization with SARS-CoV-2-Montanide ISA-51VG and SARS-CoV-2-Montanide ISA-720VG vaccines showed a tiny increase in IgG2a response and a significant increase in IgG2a/IgG1 ratio, compared with the Alum-based vaccine. Considering IgG1, as a symbol of Th2 response, and IgG2a and IgG2a/IgG1 ratio, as a symbol of Th1 response (Mahdavi et al, 2017a, 2017b), this finding showed the potency of oil-based vaccines in the polarization toward Th1 responses.
Furthermore, due to the critical role of the RBD protein in the attachment to ACE2 receptor of host cells and anti-RBD IgG antibody as a potentially neutralizing factor against SARS-CoV-2 virus, IgG response against the RBD protein was assessed, which may potentially show the neutralization activity (Khoury et al, 2021; Starr et al, 2021). Inactivated SARS-CoV-2+Alum and SARS-CoV-2-Montanide ISA 720VG vaccine groups showed a comparable IgG response versus each other against RBD protein and an increase versus SARS-CoV-2-Montanide ISA51VG group. These findings showed that Montanide-based vaccine is able to trigger the anti-RBD response similar to Alum-based vaccine. However, oil-based vaccines were superior to Alum-based vaccines in the induction of Th1 cytokine and CTL activity, which is critical in controlling SARS-CoV-2 infection.
Conclusion
In conclusion, results from this study provide evidence that the type of adjuvant in the vaccine formulation can modify the type of immune responses as well as potency of humoral immune responses. This finding encourages the SARS-CoV-2 vaccinologists to focus on different formulations of vaccine to achieve a more potent and safe vaccine, especially for inactivated SARS-CoV-2-based vaccine, which has lower efficacy than mRNA-based vaccines.
Footnotes
Acknowledgments
The authors are grateful to Borna Zist Pazhohan Knowledge Company and their staff for funding and technical support of this project. We also thank Dr. Morteza Taghizadeh and the staff of Vaccine Formulation Laboratory from Department of FMD vaccine production of Razi Vaccine and Serum Research Institute of Iran for technical support.
Authors' Contributions
M.H., A.K., and R.K. are from the Agricultural Research, Education and Extension Organization (Karaj, Iran); P.K. is from Islamic Azad University, Tehran Branch (Tehran, Iran); and M.M. is from the Academic Center for Education, Culture and Research and Tehran University of Medical Sciences (both in Tehran, Iran), all where education and research are primary functions.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was partially supported by a grant from Razi Vaccine and Serum Research Institute of Iran (grant no. 2297/250).