
Abstract
Several studies have reported that hepatitis B virus (HBV) infection is mediated by macrophages and that the B7x (B7-H4, VTCN-1) protein plays an important role in immune regulation in HBV-associated hepatocellular carcinoma (HBV-HCC). However, the relationship among HBV, macrophages, and B7x has not been studied. In this study, HBV-infected mouse model and coculture of HBV cell lines and macrophages were used to observe the changes in macrophages and the role of B7x after HBV infection. The expression of HBV markers (HBeAg, HBsAg), negative regulator of immunity (B7x), T-helper 17 (Th17)/T-regulatory (Treg)-related cytokines, and macrophage markers, as well as changes in the apoptosis and cell cycle of macrophages were analyzed through reverse transcription quantitative polymerase chain reaction, enzyme-linked immunosorbent assay, western blot, and flow cytometry. The expression of HBsAg, HBeAg, and B7x increased and the levels of macrophage surface marker and Treg cells secrete related cytokines (IL-10 and TGF-β) were altered after HBV infection both in vivo and in vitro.
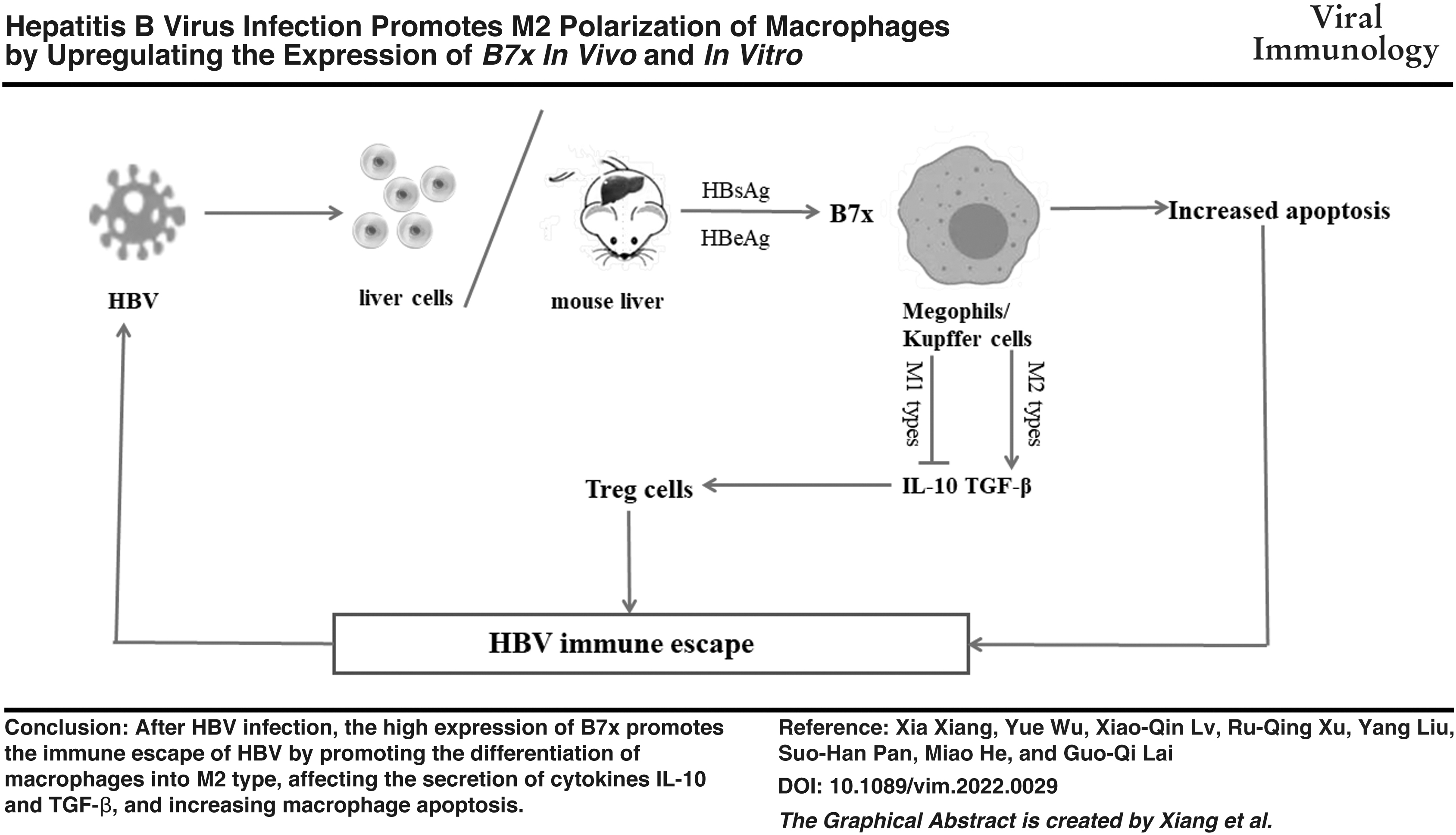
Introduction
Hepatitis B virus (HBV) infection is a major disease threatening human health all over the world. Despite the widespread use of hepatitis B vaccines, an estimated 250 million people are chronically infected with HBV (Khanam et al, 2021). The interaction between HBV and the immune mediators of the body leads to varying degrees of liver damage, such as acute and chronic hepatitis, liver fibrosis, cirrhosis, and even liver cancer. Therefore, it is important to study the efficacy of immunotherapy in treating HBV.
Macrophages of the liver, composed of tissue-resident Kupffer cells (KCs) and monocyte-derived macrophages (MDMs), play an indispensable role in the immune response induced by HBV infection (De Simone et al, 2021). It is reported when the body is infected with HBV, M1-activated MDMs secrete less proinflammatory cytokines (IL-6, IL-1β), whereas M2-activated MDMs secrete more interleukin-10 (IL-10) (Faure-Dupuy et al, 2019). It has been reported that HBsAg, an HBV antigen marker, can inhibit the activity of KCs by regulating the balance between T-helper 17 cells and T-regulatory lymphocytes (Th17/Treg balance) (Liu et al, 2018). Although it is known that macrophages regulate HBV infection through T cell dysfunction and the TLR4/NF-κB signaling pathways (The classical signaling pathway of immune response) (Zhang et al, 2021), the exact mechanisms underlying this regulation are still not clear.
B7x (B7-H4, B7S1, V-set domain-containing T cell activation inhibitor 1), a member of the B7 family, is a cosuppressor molecule expressed by tumor cells and tumor-associated macrophages that plays a negative immunoregulatory role in cancers by inhibiting T cell-mediated inflammation (Suh et al, 2006; Zang et al, 2003). Studies have found that B7x is highly expressed in hepatocellular carcinoma (HCC) cell lines as well as in CD3+ T cells and CD68+ macrophages from HCC patient tissues, but not in normal hepatocytes. B7x expression is closely related to the occurrence and development of HBV-associated HCC (HBV-HCC) and acute-on-chronic liver failure (Abadi et al, 2013; Guo et al 2012; Yuan et al, 2016). Therefore, we speculated that B7x was involved in the interaction between HBV and KCs/MDMs.
In this study, we evaluated the joint expression of B7x, macrophage marker molecules, and Th17/Treg-related cytokines. We also explored the roles of HBV-B7x-macrophages in vivo and in vitro. These results might provide new theories and insights into the immune mechanism of HBV infection, with implications in the development of immunotherapy strategies.
Materials and Methods
Preparation of HBV covalently closed circular deoxyribonucleic acid
The preparation of HBV covalently closed circular deoxyribonucleic acid (cccDNA) has been performed as described before (Lv et al, 2021). In short, the HBV-pEASY plasmid previously constructed by our laboratory (Lv et al, 2021) was used as a template, the HBV gene fragment was amplified with a high-fidelity polymerase, cut by a BspQI restriction enzyme, and ligated by T4 DNA ligase. Finally, HBV cccDNA was purified.
Establishment of a CBA/CaJ mouse model
The HBV CBA/CaJ mouse infection model was constructed following our group's previous modeling (Cao et al, 2018). The method is described as follows: CBA/CaJ mice with an average body weight of 18–22 g and 8–10 weeks of age were selected. Eight mice were used in each of the two groups: experimental and control group. The control group was injected with 2 mL normal saline, and the experimental group was injected with 2 μg/2 mL HBV cccDNA (prepared as described previously) (Lv et al, 2021) through the high-pressure hydrodynamic tail vein injection method. Mice were infected with HBV for 4 weeks, after which peripheral blood and liver tissue were collected and preserved (Wang et al, 2017).
Institutional Review Board Statement
The animal experiments in this study followed the guidelines of the state and related institutions for the use of experimental animals and were approved by the Animal Ethics Committee of Chongqing Medical University (No. 2021052).
Cell lines
Various cell lines (L02, Huh7, and Thp-1 cells) were obtained from American Type Culture Collection (ATCC, Manassas, VA, USA) and cultured in high-glucose DMEM (Thermo Fisher Scientific, USA) containing 10% fetal bovine serum (Biological Industries, Beit HaEmek, Israel) and 1% antibiotics (penicillin and streptomycin) in a 5% CO2 incubator at 37°C.
Construction of HBV cell models and B7x interference cell models
Both L02 and Huh-7 cell lines were cultured separately in six-well plates. When the cell shape is full and the density reaches 70–80%, HBV cccDNA was transfected using Lipofectamine 8000 (Beyotime, Shanghai, China) (transfected cells were named L02+HBV or Huh7+HBV cells). Alternatively, HBV cccDNA was cotransfected with the si-B7x fragment (Origen, USA) (transfected cells were named L02+HBV+si-B7x or Huh7+HBV+si-B7x cells).
Coculture cell construction
Thp-1 macrophages were prepared using the abovementioned method, and 1 mL (1 × 106 cells/mL) was placed into the upper chamber of a transwell chamber system (Corning, USA). After 48 h of Thp-1 cell stimulation with 100 ng/mL Phorbol 12-myristate 13-acetate (PMA) (Sigma, USA), the cells were attached to the wall. They were then cocultured with L02 +HBV cells, Huh7+ HBV cells, L02+HBV+si-B7x or Huh7+HBV+si-B7x cells according to the Lipofectamine 8000 operation instructions, and cells were harvested after 48 h of culture.
Enzyme-linked immunosorbent assay
The expression levels of HBsAg and HBeAg in the cell supernatant and mouse serum were detected using enzyme-linked immunosorbent assay (ELISA) according to the manufacturer's instructions for the HBV Surface Marker Diagnostic Kit (Kehua, Shanghai, China). The expression levels of B7x in mouse serum was detected by the RuiXin Mouse B7H4 ELISA Kit (Code: RX203263M) and the expression of cytokines (IL-10, transforming growth factor-β [TGF-β]) in cell supernatants were detected using the Human IL-10 ELISA Kit (Code: 3520-1HP-1; MABTECH) and the Human TGF-β ELISA Kit (Code: 3410-1HP-1; MABTECH).
Reverse transcription quantitative polymerase chain reaction
RNA was extracted from cells and tissues using TRIzol (Invitrogen, Carlsbad, CA, USA) and 2 μg RNA was reverse transcribed into cDNA using the RT Master Mix for qPCR (gDNA Digester Plus) Kit (Med Chem Express, Monmouth Junction, NJ, USA). The expression of B7x and macrophage markers (including CD86, INOS, CD206, and Arg1) in cells and tissues and the cytokine expression in liver tissue were detected by reverse transcription quantitative polymerase chain reaction (RT-qPCR). A Bio-Rad CFX Connect Real-Time PCR Detection System (Bio-Rad, Hercules, CA, USA) was used as per the manufacturer's instructions for RT-qPCR experiments. The primer sequences are listed in Table 1 (Liu et al, 2020; Sadun et al, 2007; Wang et al, 2020; Yan et al, 2021; Zhang et al, 2019; Zheng et al, 2020).
Reverse Transcription Quantitative Polymerase Chain Reaction Primer Sequences
Western blotting
Total cellular protein was extracted using Radio Immunoprecipitation Assay (RIPA) lysate containing phosphatase and protease inhibitors (Beyotime, Shanghai, China). Total protein was quantified using the BCA Protein Concentration Test Kit (Solarbio, Beijing, China) and stored at −80°C. The obtained extract (containing about 40 μg protein) was added to 5 × loading buffer and denatured at 98°C for 10 min. First, sodium dodecyl sulfate–polyacrylamide gel electrophoresis (100 V, 120 min) was performed. Second, the protein in the gel was transferred to a 0.22 μm PVDF membrane (250 mA, 60 min). The membrane was then blocked with Quick Block™ Western Blocking Solution (Beyotime, Shanghai, China) for 15 min. After washing three times (15 min each), the membrane was incubated with anti-B7-H4 (D1M8I) rabbit mAb (1:1,000; CST, USA) and anti-GAPDH (D16H11) rabbit mAb (1:1,000; CST, USA) primary antibodies overnight at 4°C, followed by incubation with the secondary antibody, rabbit anti-Mouse IgG (H+L), on the next day.
Horseradish peroxidase (1:5,000; Invitrogen, USA) was added to the membrane and incubated at 37°C for 1 h. After washing three times with Tris buffered saline with Tween 20 (TBST), proteins were visualized using an Ultra-High-Sensitivity Enhanced Chemiluminescence Kit (HY-K1005; Med Chem Express, USA). Each experiment was repeated three times.
Flow cytometry
Apoptosis and cell cycle stages of Thp-1, Th17, and Treg cells were detected by flow cytometry. First, Thp-1 cells in each group were harvested and washed twice with phosphate-buffered saline (PBS) (1,000 × g for 5 min). Cells (1 × 106) were resuspended in 50 μL PBS with 5 μL Annexin V-FITC and Propidium Iodide (PI) (CA1510-50T; Solarbio, Beijing, China); cells were then incubated at room temperature (25°C) in the dark for 15 min. Apoptosis was detected based on Annexin V and/or PI fluorescence. Then, Thp-1 cells in each group were harvested and washed twice with PBS; 1 × 106 cells were resuspended in precooled 75% ethanol and fixed overnight. Apoptosis and cell cycle stages were detected by flow cytometry.
In addition, 200 μL per tube of anticoagulated blood was collected from mice, red blood cells were lysed using red blood cell lysate (BioLegend, USA), and CD4 and CD25 antibodies (BioLegend, USA) were added, respectively. The nuclear membrane was fragmented using the Fixed Membrane Breaking Kit (BioLegend, USA), and IL-17A and Foxp3 antibodies (BioLegend, USA) were added and detected by flow cytometry.
Statistical analysis
All data were analyzed using GraphPad Prism 8.0 and Student's t-test (two-tailed and with equal variance). All experiments were independently repeated at least three times. Statistical significance was set at p < 0.05.
Results
Alterations in B7x, macrophage types, Th17, and Treg after in vivo HBV infection
The HBV CBA/CaJ mouse model was established, and serum and liver tissue samples were collected at 4 weeks to detect relevant indicators. HBsAg and HBeAg were found to be positive in the serum of infected animals (p < 0.001) (Fig. 1A, B). Similarly, B7x levels were significantly increased in serum and liver tissue (p < 0.001) (Fig. 1C, D). Next, we examined the expression of related indicators in liver tissue; the expression of M2 macrophage markers, CD206 and Arg1, showed significantly increased expression (p < 0.01). The expression of IL-10 and TGF-β significantly increased (p < 0.01) (Fig. 1E–G). Meanwhile, the proportion of Th17 cells decreased while the number of Treg cells increased (p < 0.05) (Fig. 2A–D). These results suggest that an HBV infection can increase B7x expression, promote M2 polarization, and disturb the Th17/Treg balance.
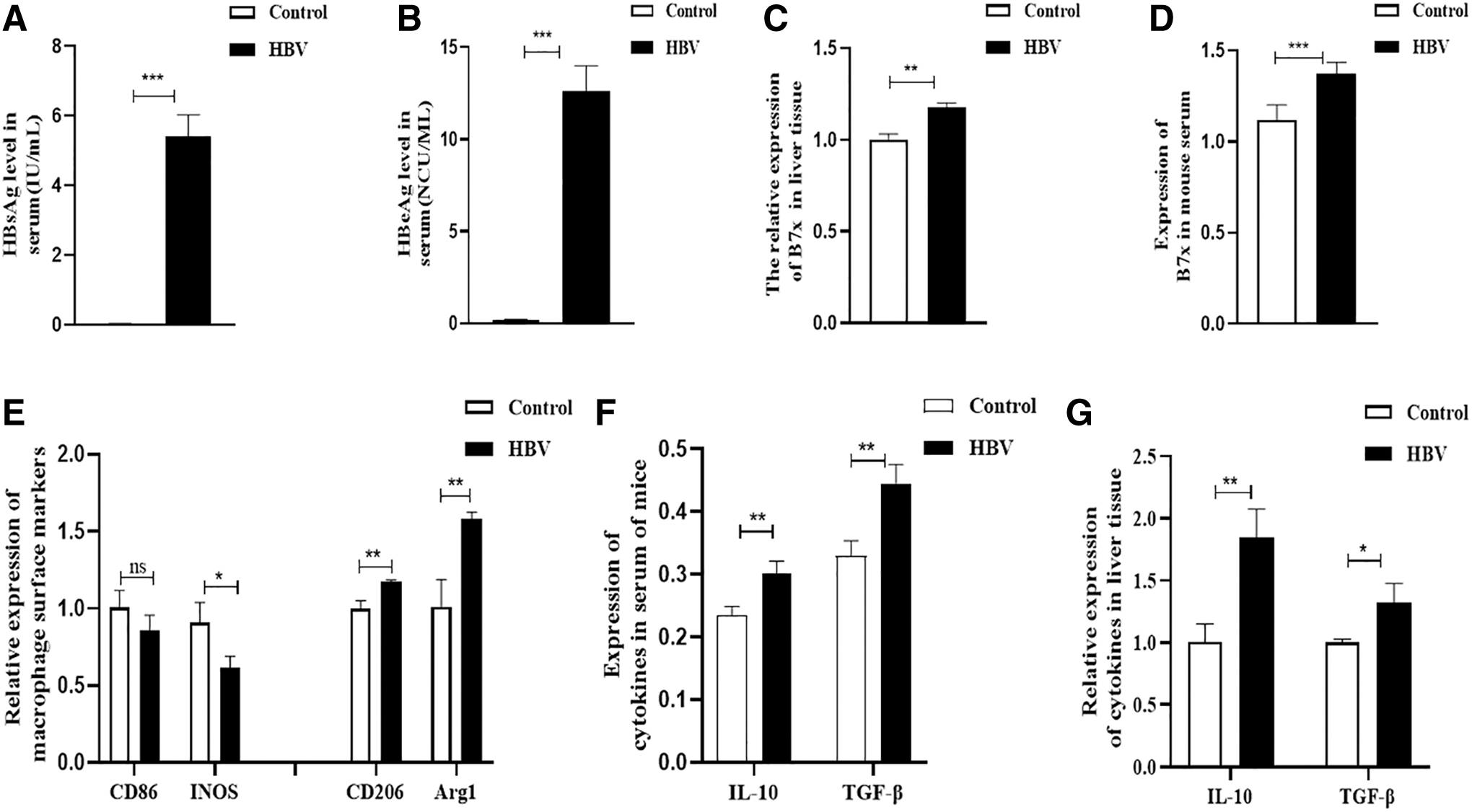
Expression of HBV antigen markers, B7x and macrophage types.
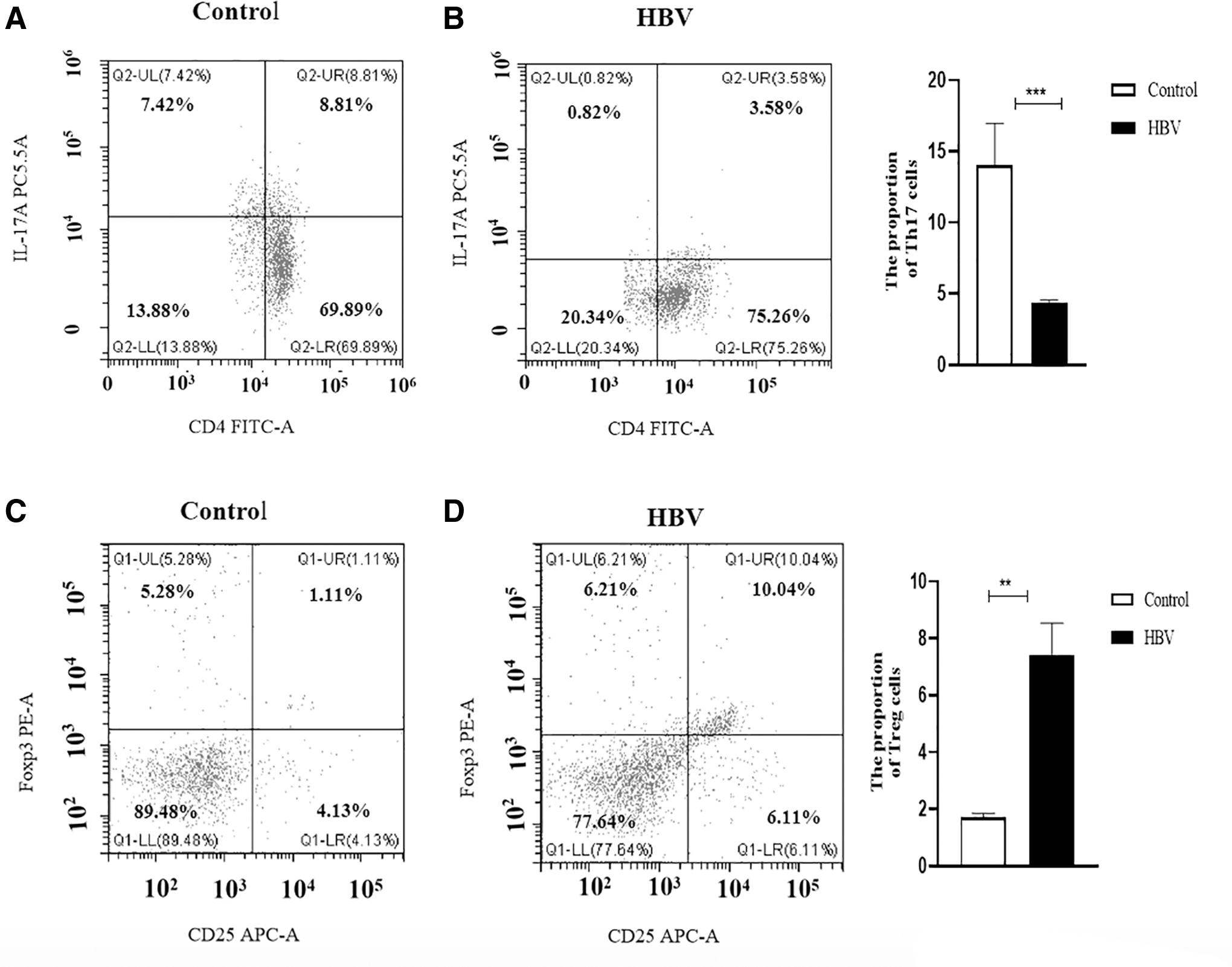
The secretion of Th17 cells and Treg cells was affected by HBV infection in mice.
HBV promotes the expression of B7x in vitro
To observe the relationship between HBV and B7x, the L02/Huh7+HBV cell line models were used. The expression of HBV antigen markers (HBsAg and HBeAg) in the cell supernatant was detected by ELISA. The expression of B7x messenger ribonucleic acid was detected by RT-qPCR and B7x protein by western blotting. The results showed that the expression of HBV surface markers was increased after transfection (p < 0.001) (Fig. 3A–D), and the expression of B7x in cells was also significantly increased (p < 0.001) (Fig. 3E–H). These results indicate that HBV infection promotes B7x expression.

Increased expression of HBV marker B7x after infection of L02 and Huh7 cells with HBV.
B7x can regulate the polarization of Thp-1 cells and affect their cycle and apoptosis
To further observe the effect of B7x on macrophages and its potential mechanism of action, Thp-1 was stimulated with PMA for 48 h and then cocultured with L02/Huh7+HBV cells. The expression of HBeAg and HBsAg in the cell supernatant significantly increased after transfection with HBV (p < 0.001) (Fig. 4A–D), whereas the expression of B7x was high in L02 and Huh7 cells after infection with HBV (p < 0.001) (Fig. 4E, F). The changes in surface markers of M1 macrophages (CD86, INOS) were negligible (p > 0.05), but the levels of M2-type surface markers (CD206, Arg1) significantly increased (p < 0.05) (Fig. 4G, H). Meanwhile, in the supernatant of L02 and Huh7 cells co-cultured with Thp-1 cells, the expression of IL-10 and TGF-β were significantly increased after infection with HBV (p < 0.05, p < 0.001, p < 0.01, respectively) (Fig. 4I, J). Interestingly, after HBV infection, the proportion of apoptotic Thp-1 cells increased significantly (p < 0.05) (Fig. 5A, B), and the cell cycle of Thp-1 cells was arrested at the G1/M phase (p < 0.01) (Fig. 5C, D).
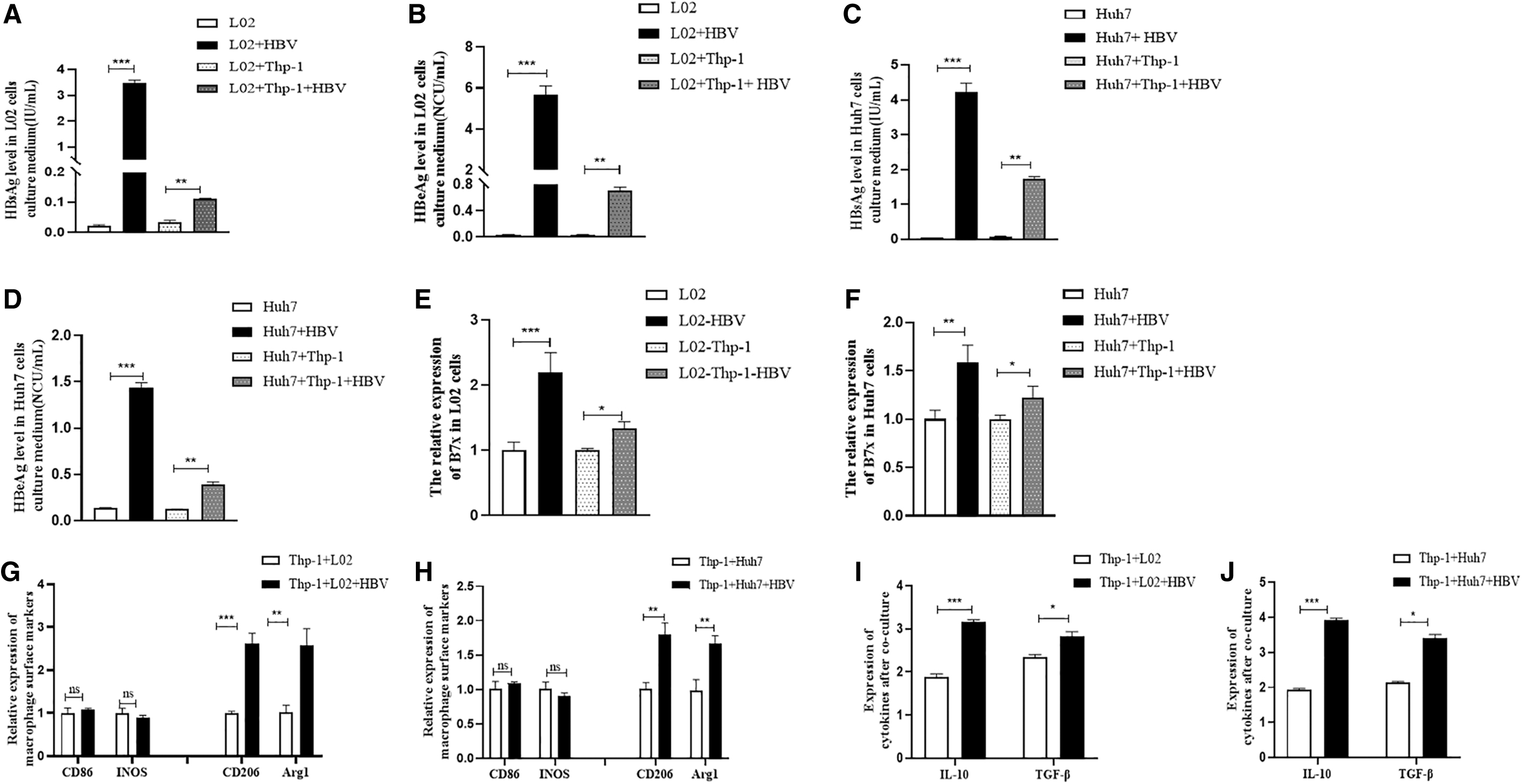
HBV infection promotes the expression of B7x and Changes the polarization of Thp-1 cells.
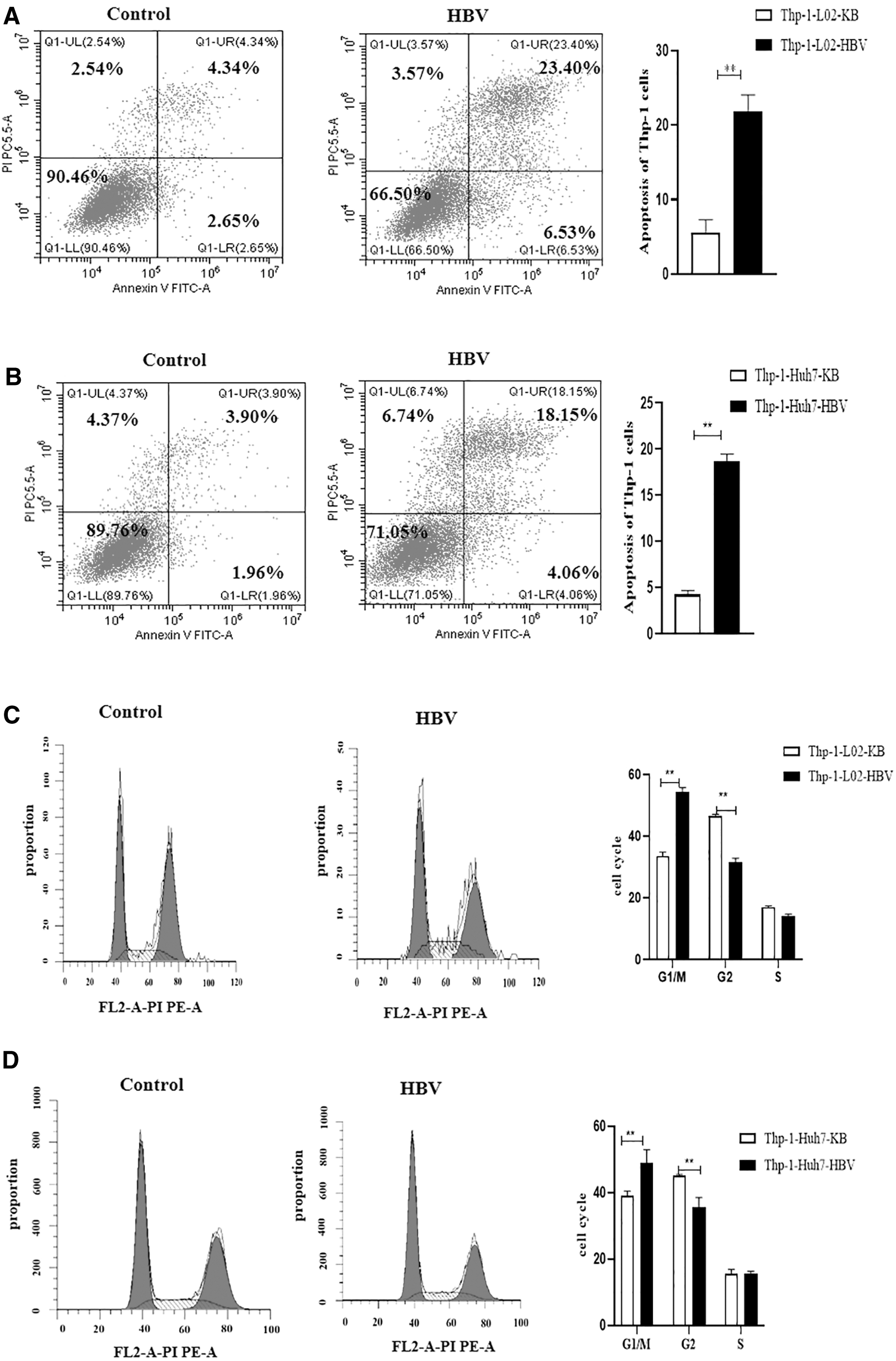
The high expression of B7x changed the apoptosis and cycle of Thp-1 cells.
B7x knockdown can promote HBV immune escape by changing macrophage state
To analyze the role of B7x in macrophage polarization and cytokine secretion after HBV transfection, si-B7x was transfected into liver/HCC cell lines (L02/Huh7) that had been previously transfected with HBV in a coculture system with Thp-1. The expression of HBsAg and HBeAg was also significantly decreased as B7x decreased (Fig. 6A–F). Both CD86 and INOS levels in L02+HBV+si-B7x+Thp-1 cell model, and CD86 levels in Huh7+HBV+si-B7x+Thp-1 cell model were unchanged, although INOS levels increased in Huh7+HBV+si-B7x+Thp-1 cell model, whereas the expression levels of CD206 and Arg1 decreased after cotransfection of HBV and si-B7x (Fig. 6G, H). Interestingly, IL-10 and TGF-β levels were decreased in the supernatant of the coculture cell system (p < 0.05) (Fig. 6I, J). Thp-1 cell apoptosis rates were decreased (p < 0.05) (Fig. 7A, B), but the cell cycle showed no changes (Fig. 7C, D).

Interference with B7x inhibits the effects of HBV on the polarization of Thp-1 cells.

After HBV infection interferes with B7x, Thp-1 cell apoptosis can be reduced.
Discussion
Numerous clinical studies have shown that chronic HBV persistent infection can lead to dysfunction of innate and adaptive immune responses (Hong et al, 2016). Therefore, immunotherapy, a method for curing chronic HBV infection by enhancing HBV-specific immunity, may serve as an effective antiviral treatment strategy. B7x inhibits T cell-mediated inflammatory responses (Chen et al, 2014), and when B7x binds to the receptor, T cell proliferation, cytokine production, and cell cycle progression are all negatively regulated, thereby reducing T cell immune responses. Moreover, it is worth mentioning that B7-H4 has been reported to be significantly upregulated in HepG2.2.15, HCC, and HBV-HCC patients (Chen et al, 2014; Prakoso et al, 2006; Zang et al, 2003), which suggests that HBV and B7x may have some connection, providing a basis for HBV immunotherapy potential avenues. In this study, an in vivo HBV infection model in CBA/CaJ mice was established by tail vein injection of HBV cccDNA, and an HBV-infected hepatoma cell model was obtained in vitro. In in vitro and in vivo experiments, B7x was well demonstrated to be significantly elevated in HBV infection models.
Polarization of macrophages is a key pathway for regulating immunity, and studies have reported that it is closely related to HBV infection. M2 macrophages regulate the secretion of various cytokines, such as IL-4, IL-6, and IL-10, and in previous studies, we have found that these cytokines are significantly increased in HBV-infected mice (Zhao et al, 2020). In the present study, we found that mouse KCs differentiated into M2 type after HBV infection, which is consistent with the results of Li (Abadi et al, 2013; Li et al, 2020). In addition, the secretion of IL-10 and TGF-β is increased after HBV infection, and both cytokines have been shown to be involved in Treg-mediated immunosuppression and affect Th17 differentiation (Rinaldi et al, 2021); at the same time, we also found that Th17 cells decreased and Treg cells increased in anticoagulation of HBV mice, which is consistent with the findings of Chen (Li et al, 2013), suggesting that HBV infection changes the Th17/Treg balance.
More interestingly, when we further explored the relationship between B7x and macrophage Thp-1, we found that the high expression of B7x may promote the increase of macrophage apoptosis and inhibit its cell cycle in G1 phase. These results demonstrate that B7x is closely related to macrophage polarization and Th17/Treg balance after HBV infection.
B7x plays an important role in various cancers and inflammation because it has an immune function (Bility et al, 2014) to inhibit T cell-mediated inflammatory responses. The expression of B7x can also be used as a prognostic marker of gastric cancer (Li et al, 2019; Sica et al, 2003). Therefore, we proposed that the observed phenomenon is caused by the high expression of B7x. To test this hypothesis, we cocultured Thp-1 cells with HBV-infected hepatoma cells in vitro. We were surprised to find that, in addition to the significantly reduced expression of HBsAg and HBeAg, macrophage Thp-1 showed a M1-type differentiation trend, the secretion of IL-10 and TGF-β was significantly reduced, and more importantly, the apoptosis rate of macrophages (Abadi et al, 2013) was significantly reduced, although their cell cycle was not significantly different. Taken together, our findings fully demonstrate that B7x can serve as an important potential way of immunotherapy for HBV-infected patients, even HBV-HCC patients, and it is of great significance to further study the potential mechanism of B7x and HBV infection.
Conclusions
Overall, the results suggest that B7x expression increases after HBV infection. At the same time, B7x can promote the differentiation of macrophages into M2 and induce secretion of IL-10 and TGF-β that stimulate Treg cells. B7x can promote apoptosis of immune cells. However, this study has some limitations; it is not confirmed whether HBV infection regulates macrophage phenotypic changes and Th17/Treg balance, based on experiments performed in the B7x knockout model mouse. However, the present study is the first to investigate the correlation between HBV and the negative immunoregulatory factor B7x in immune-healthy HBV-infected mice; the results of the current study revealed for the first time that B7x may lead to immune escape and immune cell incompetence after HBV infection by changing macrophage polarization and apoptosis to regulate the Th17/Treg balance. This new discovery suggests that B7x may be a new target for HBV immunotherapy and provides a new research direction for the clinical treatment of HBV infection.
Footnotes
Acknowledgments
The authors would like to thank the Key Laboratory of Molecular Oncology and Epigenetics of the First Affiliated Hospital of Chongqing Medical University for providing cell lines, thank Sanxiu He for providing technical guidance on cell coculture, and thank Zhihao Feng for providing technical guidance on western blot.
Authors' Contributions
G.-Q.L., X.X., and Y.W. provided experimental operation ideas and wrote articles together. X.X. and Y.W. participated in the whole experiment and data analysis; X.-Q.L. provides cell resources for experiments; Y.L. provided animal model construction for experiments; R.-Q.X., S.-H.P., and M.H. participated in the experiments of mouse model. All authors have read and agreed to the published version of the article.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Natural Science Foundation of China [Grant No. 81570541], and the Min-Sheng Project of Chongqing Science and Technology (Grant No. cstc2017shmsA130097).