
Abstract
Various vaccines are under development to prevent chikungunya (CHIKV) infection. For the assessment of the CHIKV vaccine-induced antibody response, it is extremely important to understand antibody response after the infection has occurred. Previously, we assessed IgG response in samples from healthy donors using I-CHIKV and found that IgG1 was the predominant subclass induced after CHIKV infection followed by IgG4. However, IgG3 subclass induction is reported in serum samples from patients with acute CHIKV infection. Therefore, in this study, we evaluated serum/plasma from samples of patients with acute CHIKV infection for the presence of IgG and IgG subclasses against I-CHIKV and recombinant E2 protein (rE2). Out of 44 samples that were positive against I-CHIKV, 43 were found reactive against rE2. The positivity of IgG1 either alone or together with other IgG subclasses using I-CHIKV was 89% samples, while 86% samples were positive using rE2. High titers of IgG1 are obtained with I-CHIKV (67%), while raised IgG4 levels are detected using rE2p (72%) in the samples that are positive for both these subclasses. Testing of 22 samples for neutralizing antibodies revealed 100% IgG1 positivity and neutralizing antibodies in 21, 1 sample negative for both. Overall, these data will be useful in assessing IgG subclass-specific CHIKV neutralization and response after CHIKV immunization.
Introduction
Chikungunya (CHIKV) is known to be associated with prolonged arthralgia (Chow et al, 2011; Ninla-Aesong et al, 2020). The acute febrile phase of the illness normally resolves within a few days, while the joint pain associated with CHIKV infection persists for weeks or months. Clinically, it has been demonstrated that most of the CHIKV patients develop severe arthralgia with higher circulatory proinflammatory cytokines, suggesting possible role of host immune response in pathogenesis (Chow et al, 2011).
IgG antibody and its subclasses, which serve as critical factors in virus neutralization, can also modulate immune response after antigen-antibody reaction (Bournazos et al, 2017; Yoon et al, 2015). Antibody-mediated aggravation of CHIKV infection and disease severity has been reported in mice (Lum et al, 2018). In such a scenario, it is crucial to understand the subclasses of antibody induced after infection or immunization as they are known to play differential roles in disease protection or exacerbation due to varied affinity and binding specificity of Fc region of IgG subclasses to Fc receptor (FcγR) (Bournazos et al, 2020).
Previously, using I-CHIKV, we showed IgG1 to be the predominant subclass in CHIKV-infected individuals (Patil et al, 2021), a typical response observed after viral infections in humans in convalescent (Hofmeister et al, 2011; Isa et al, 2002). The study was conducted employing convalescent samples. Surprisingly, IgG1 responses was followed by IgG4, an antibody subclass that is predominantly elicited against carbohydrate antigens and constitutes <5% of total IgG under normal conditions (Aalberse et al, 2009). Usually, in viral infections, other subclasses have been observed to be induced together with IgG1 (Hofmeister et al, 2011; Isa et al, 2002). Earlier study has reported induction of IgG1 and IgG2 in acute-phase plasma samples with negligible IgG3 and no IgG4 after CHIKV infection (Nayak et al, 2020).
This study and our previous study demonstrated that IgG1 was detected consistently, while IgG2 and IgG4 were not present in all samples (Nayak et al, 2020; Patil et al, 2021). On the contrary, other reports have shown IgG3 to be the only antibody induced after CHIKV infection (Kam et al, 2012a; Kam et al, 2012b; Kam et al, 2012c). These studies were mainly carried out using samples from patients in acute stage of infection.
In view of the reports of differential IgG subclass induction, this study was conducted using samples from acute-phase CHIKV patients to further understand the IgG subclass response toward I-CHIKV and E2.
Materials and Methods
Ethics
This study was approved by the human ethics committee of the Institutional Ethics Committee of Bharati Medical College (IEC/2017/04, renewed IEC/2018/11). No prospective sample collection was done.
Serum samples
The study was conducted using 44 blood samples collected between 2017 and 2019 (monsoon season), at Bharati Hospital (Pune, India). All samples had tested positive for anti-CHIKV-IgM and anti-CHIKV-IgG. The duration between exposure to CHIKV and sample collection is not known.
Enzyme-linked immunosorbent assay
Serum/plasma samples were evaluated for anti-CHIKV-IgM as per manufacturer's instructions (Cat. No. ab177835; Abcam). Same samples were analyzed for anti-CHIKV-IgG positivity and titers as per protocol described earlier (Patil et al, 2020). Anti-CHIKV-IgM and anti-CHIKV-IgG samples were further evaluated for IgG positivity and titers using recombinant E2 protein (rE2) protein. Briefly, 100 μL 0.75 μg/mL E2 protein was coated using carbonate-bicarbonate buffer, pH 9.2, in enzyme-linked immunosorbent assay (ELISA) plates (Greiner-BioOne). Coated wells were incubated overnight at 4°C and washed five times using 300 μL phosphate buffer saline (PBS) containing 0.05% tween 20 (Sisco). Plates were blocked for 1 h at 37°C using PBS containing 10% fetal bovine serum (FBS; Gibco) and washed five times with wash buffer. Washing protocol was kept constant for all further washing steps.
Diluted samples were double diluted in sample diluent (PBS containing 10% FBS, 1% Vero cell supernatant, and 0.1% tween 20), and 100 μL diluted sample was added to plates and incubated for 1 h at 37°C. Samples were initially diluted 1:10 followed by dilution to 1:100. After incubation, wells were washed and 100 μL conjugate diluted in PBS with 10% FBS and 0.1% tween 20 was added to wells. The conjugates were added individually to the wells based on the type of antibody to be detected. HRP-conjugated antibodies with different concentrations/dilutions were added to the wells. The dilution of anti-human-IgG-HRP (Sigma-Aldrich) was 1:20,000, while that of anti-human-IgG1-HRP, anti-human-IgG2-HRP, anti-human-IgG3-HRP, or anti-human-IgG4-HRP (all from Southern Biotech) was 1:5,000.
After incubation for 30 min at 37°C, 100 μL tetramethylbenzidine (TMB) substrate containing hydrogen peroxide (Clinical Science Products, Inc.) was added and reaction was stopped after 10 min using 50 μL 2 N H2SO4. Plates were read at 450 nm with background subtraction of 655 nm. Cutoff value was determined by multiplying optical density (O.D.) value of negative control by 3.
Plaque reduction neutralization assay
CHIKV-specific plaque reduction neutralization test 50 (PRNT50) was performed using n = 22 samples as described previously (Patil et al, 2020). Briefly, Vero cells were seeded in 24-well plate (Corning) using MEM with 10% FBS (Gibco) with penicillin and streptomycin. Twenty-four hours after seeding, heat-inactivated serum samples were diluted fourfold in MEM with 2% FBS. Equal volume of 500 PFU/mL CHIKV was mixed with diluted serum samples, incubated, and 100 μL was added to Vero cell monolayer. After incubation for 1 h, the mixture was removed and overlay medium was added to Vero cell monolayer. Plates were incubated for 48 h, overlay medium was discarded, and cells were stained using 1% crystal violet. Plaques were counted after thorough washing and drying of crystal violet-stained plates.
Statistical analysis
GraphPad Prism v8.1.1 (Graphpad Software) was used to perform statistical analysis. Two tailed Mann-Whitney U test was used to compare antibody titers between groups. Correlation coefficients between IgG and IgG subclasses were quantified by the Spearman rank correlation coefficient. **, ***, and **** represent p < 0.01, p < 0.001, and p < 0.0001, respectively.
Results and Discussion
Comparison of anti-CHIKV-IgG subclass antibody positivity in CHIKV patients by ELISAs using I-CHIKV and rE2p as coating antigens
We have compared IgG and IgG subclass seropositivity in 44 samples that are positive for anti-CHIKV-IgM and anti-CHIKV-IgG antibodies employing I-CHIKV or rE2p (Table 1). Out of 44 samples that are positive using I-CHIKV, 43 (98.7%) were found positive using rE2p-ELISA. IgG subclass analysis (Table 2) revealed maximum detection of IgG1 using both I-CHIKV (89%) and rE2p (86%) followed by IgG4 (55% and 65%, respectively, p > 0.1 for both). E2 protein-specific IgG2 was detected in 18% patients, while IgG3 was found in 7%. None of the samples were positive for all the subclasses, while three subclasses were found in 11% (n = 5) and 16% (n = 7) samples using I-CHIKV and rE2, respectively.
The Table Shows Distribution of the 44 Study Samples According to Their Reactivity Toward I-CHIKV and rE2p for Total IgG and the Four IgG Subclasses
Overall, 43 samples out of 44 were found positive for both I-CHIKV and rE2. The subclass reactivity of the samples varied depending on the coating antigen.
CHIKV, chikungunya; rE2, recombinant E2 protein.
Differences in IgG Subclass Positivity When I-CHIKV and rE2p Was Used as Coating Antigen
The values in bracket indicated percentage positivity.
Using rE2p, one sample each was found to be positive for only IgG3 and IgG4 with minimal titers of 100 and 200, respectively. For I-CHIKV-based ELISA, samples having O.D. <0.85 were tested negative for IgG subclass. Possibility of minimal reactivity of IgG3 to I-CHIKV and subclass shift over time was considered reasons for low positivity of IgG3.
However, our observations with rE2 confirmed that our results are indeed different from other studies (Kam et al, 2012a; Kam et al, 2012b; Kam et al, 2012c). Viral inoculums during infection or immunological factors may not be possible reason for these striking differences. Our results are in line with the results for other RNA viruses such as West Nile and Measles (Hofmeister et al, 2011; Isa et al, 2002), where IgG1 was shown to be the predominant antibody induced after infection. The unavailability of IgG3 reactive peptide epitopes on the coating antigens cannot be ruled out for these differences.
Antibody subclass response after CHIKV infection
IgG1 and IgG3 responses are typically elicited against viral proteins (Hofmeister et al, 2011; Isa et al, 2002; Wagner et al, 1987). To evaluate whether the antibody response in terms of titers differs for whole virion or individual proteins, we evaluated the IgG and its subclass responses using I-CHIKV and rE2p in the samples reactive for anti-CHIKV-IgM and anti-CHIKV-IgG antibodies (Fig. 1A). High IgG titers were obtained against I-CHIKV compared to rE2p (p < 0.0001, Fig. 1C). Similar results were obtained for IgG1 subclass (p < 0.0001, Fig. 1D).
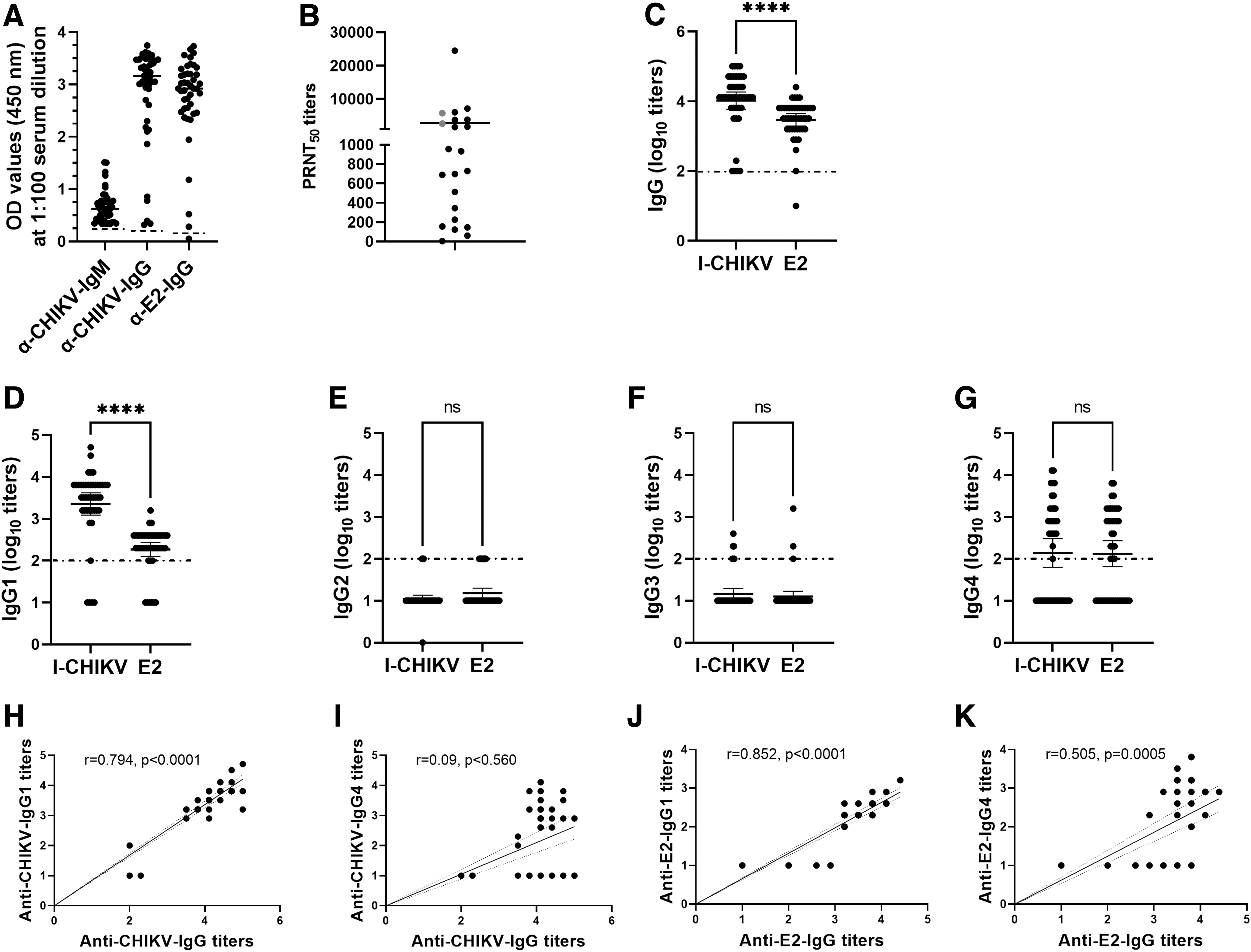
Antibody response to CHIKV. N = 44 samples that were positive for anti-CHIKV-IgM and anti-CHIKV-IgG antibodies were evaluated for the presence of
The titers (Fig. 1E–G) and the pattern of titers for IgG2, IgG3, or IgG4 antibody subclasses were similar for both the antigens (p = 0.127–0.759). A possible explanation for high IgG and IgG1 titers against I-CHIKV compared to rE2p could be binding of antibodies to multiple epitopes of E1, E2, and E3 present on CHIKV surface versus limited epitopes present on E2. Of note, a strong correlation was observed between IgG and IgG1 titers for I-CHIKV (r = 0.794, p < 0.0001) and rE2p (r = 0.852, p < 0.0001).
To understand the association of neutralizing antibodies with IgG subclasses, we tested 22 available samples by PRNT (Fig. 1B). Among these, one exhibited low OD values in both the ELISAs (0.39, 0.32), was negative for all IgG subclasses, and scored PRNT negative. Of the remaining 21 samples, all and 2 were reactive for IgG1 and IgG3 antibodies. These results suggest crucial role of IgG1 in CHIKV neutralization. Of note, two IgG3-positive samples were also IgG1 reactive. For the clarification of contribution of different IgG subclasses in CHIKV neutralization, a larger number of freshly collected samples need to be tested by using subclass-specific antibodies and immunostaining.
Although the reactivity of IgG4 was low compared to IgG1, a positive correlation with IgG titers was observed for rE2p (Fig. 1J, K, r = 0.505, p = 0.0005), but not with I-CHIKV (Fig. 1H, I, r = 0.09, p = 0.560). The differential reactivity indicates predominance of E2 antigen-specific IgG4. It may be noted that IgG4 is associated with autoimmune diseases (Anan et al, 2017; Fragoulis et al, 2017) and CHIKV infection has been associated with the induction of auto-antibodies (de Carvalho et al, 2021; Maek-a-nantawat and Silachamroom, 2009), and it would be important to explore possible role of IgG4 in CHIKV pathogenesis with special reference to the occurrence in a few patients.
IgG subclass ratio after CHIKV infection
IgG1 is the hallmark antibody for Th1 response in humans, while IgG4 is the outcome of Th2 response and is associated with diverse functions (Aalberse et al, 2009). Both these antibodies were predominantly produced after CHIKV infections and had reactivity to I-CHIKV and rE2p. The level of IgG subclass produced after infection might vary for different proteins. Therefore, we evaluated the ratio of IgG subclass titers obtained for either I-CHIKV or rE2p.
High IgG1 titers were observed when I-CHIKV was used (Fig. 2A) for coating. Out of 24 samples that were positive for both IgG1 and IgG4, 16 samples had 2- to 16-fold higher IgG1 titers and 4 samples had equal levels of both antibodies, while raised IgG4 levels were detected in the remaining 4 samples. On the contrary, the ratio of IgG subclasses obtained using titers toward rE2p highlighted IgG4 levels to be higher than IgG1 in 18/25 samples. IgG4 levels were 2–16 times more in 17 samples, while 1 sample had 32 times higher titers. Only seven samples had predominant IgG1 against rE2.
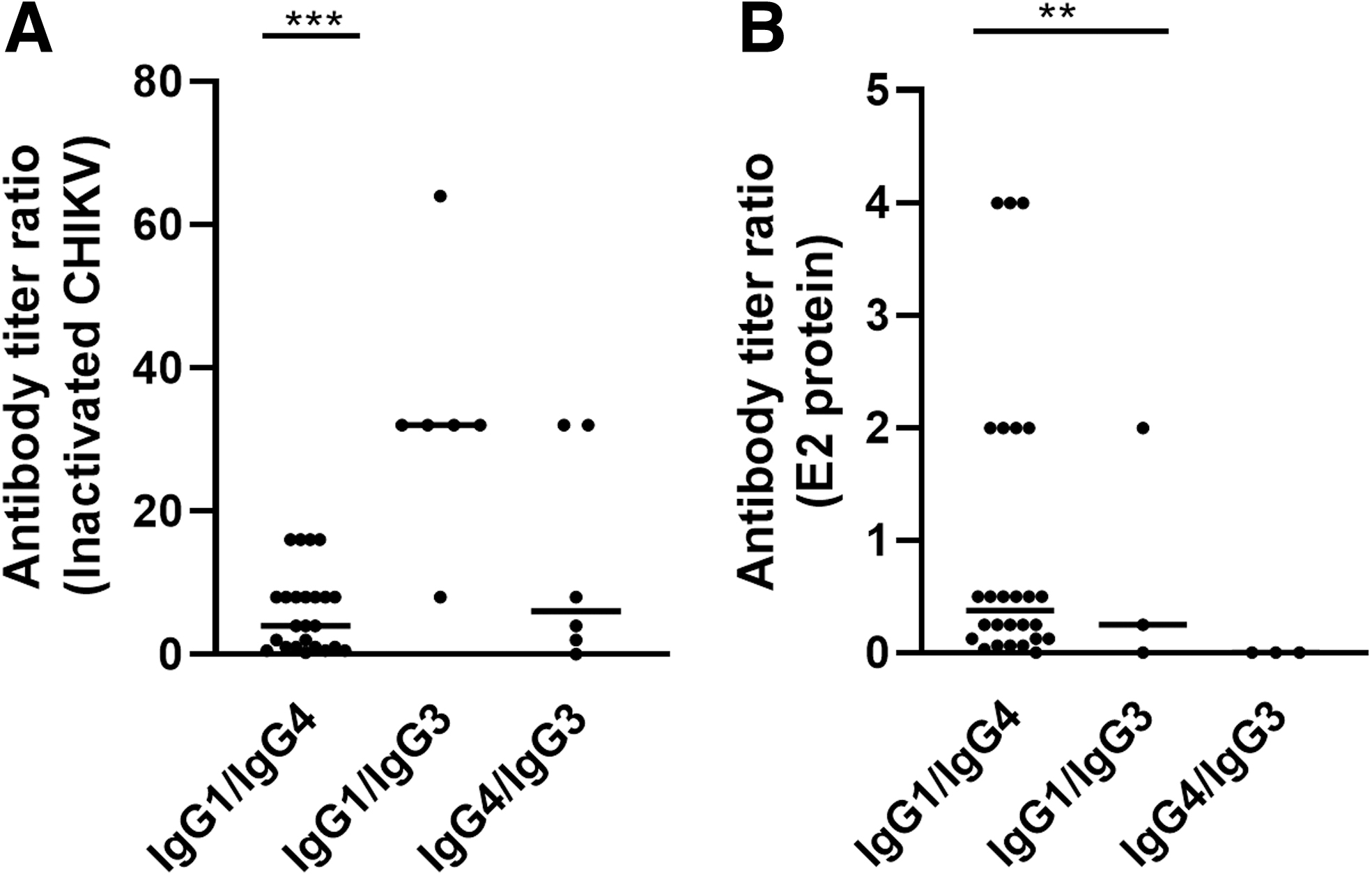
IgG subclass response ratio after CHIKV infection. Samples that were positive for both IgG subclasses were used to determine the ratio between IgG1/IgG4, IgG1/IgG3, or IgG4/IgG3. IgG and IgG subclass titers against CHIKV-E2 were determined. Ratio was determined for the samples detected using
These IgG1/IgG4 titer ratios revealed that the titers are coating antigen dependent. High titers of IgG1 are obtained with I-CHIKV, while raised IgG4 levels are detected for rE2p. These results indicate that the IgG4 subclass may predominantly be induced against E2. To determine if this is the case, further studies using other CHIKV structural proteins as coating antigens in ELISA need to be performed. Although E2 is available for antibody binding on I-CHIKV surface, the availability of epitopes that usually get hidden during interaction with E1, E3, and capsid after virus assembly might be a possible reason for higher titers of IgG4 (Sun et al, 2013; Yap et al, 2017).
While IgG1 mediates immunological effects like classical complement activation and FcγR-mediated phagocytosis, antibody-dependent cellular cytotoxicity, degranulation, and release of inflammatory mediators, the role of IgG4 in this disease is less well understood (Nimmerjahn and Ravetch, 2007). It is possible that IgG4 targeting rE2 may compete with IgG1, which could reduce the efficacy of the antibody response. Furthermore, IgG4 may contribute to or cause autoimmune-mediated disease.
Conclusion
In conclusion, the results of this study further our understanding of the IgG subclass responses elicited by CHIKV infections. We showed that in acute CHIKV patients, IgG1 is the predominant antibody in terms of positivity, titers, and excellent association with neutralizing antibodies. This subclass was followed by IgG4 with infrequent reactivity for IgG2 or IgG3. Comparison of IgG1/IgG4 titer ratios revealed that the titers were coating antigen dependent. Higher IgG1 titers were detected using I-CHIKV and IgG4 using rE2p. These data will be important for selection of the antigen for IgG subclass titer determination if induction of appropriate IgG subclass is considered one of the criteria for protection after infection/immunization. The contribution of different IgG subclasses in the neutralization of CHIKV needs to be determined.
Footnotes
Authors' Contribution Statement
H.P.P.: Conceptualization, data curation. Formal analysis, investigation, methodology, project administration, supervision, validation, visualization, writing – original draft. M.G.: Formal analysis, investigation, methodology, validation, visualization. R.K.: Formal analysis, investigation, methodology, validation. A.C.M.: conceptualization, funding acquisition, project administration, resources. V.A.A.: Conceptualization, data curation, methodology, project administration, resources, supervision, validation, visualization, writing – review and editing.
Author Disclosure Statement
None of the authors have any conflict of interest. Samples from the study “Establishment of a novel Electronic Surveillance System for DENV in Pune: an initiative for Smart Cities Mission” funded by Indian Council of Medical Research, Government of India, New Delhi, were used in this study.
Funding Information
We thank Interactive Research School for Health Affairs, Bharati Vidyapeeth (Deemed to be University), Pune, for funding the study.