
Abstract
Neutralizing antibody level is used to predict immune protection against SARS-CoV-2 infection. Spike protein of SARS-CoV-2 is a major target for virus-neutralizing antibody. A number of neutralizing epitopes were mapped on receptor binding domain (RBD) and N-terminal domain (NTD) of S1 subunit of the spike. Anti-SARS-CoV-2 antibody usually decreases over time after recovery. Level of neutralizing antibody and binding antibody to several domains from COVID-19 recovered patients was observed longitudinally in this study. Sequentially collected serum samples from 35 patients demonstrated both similar and different trends of neutralizing antibodies versus binding antibodies to each domain. Twenty-three individuals showed similarly decreasing pattern of neutralizing titer, binding antibodies to RBD, NTD, fusion protein (S2), and nucleocapsid (NP). Interestingly, eight individuals had stably high neutralizing titer (≥320) for 3–12 months, whereas their binding antibodies to RBD, NTD, and NP rapidly decreased. Moreover, their binding antibodies to S2 were stable over time similar to the persistence of neutralizing antibody levels. The long-lasting antibody to S2 suggested an anamnestic response to cross-reactive epitopes from previous infections with other related coronaviruses. These data indicate a difference in kinetics and longevity of antibodies to various domains and epitopes of the SARS-CoV-2 proteins. A better understanding in this difference may help improve vaccine design to induce long-lasting immunity to COVID-19.
Introduction
The humoral immune response to SARS-CoV-2 has been widely studied since neutralizing antibody is a hope for antiviral protection and treatment. A number of neutralizing epitopes were characterized. Potent neutralizing antibodies mainly target epitopes on spike glycoprotein (S) of SARS-CoV-2 (1,11,14,17,22). Each monomer of trimeric S-protein consists of S1 and S2 subunits. Receptor binding domain (RBD) located in the S1 subunit specifically interacts with human angiotensin converting enzyme 2 (ACE2) to mediate cell entry. RBD is considered the main target for efficient neutralization (11,14,20,24).
N-terminal domain (NTD) is also located on the S1 subunit and interacts with sialic acids or coreceptors. The neutralizing antibodies targeting NTD is usually less prevalent than those targeting RBD. S2 subunit mediates fusion of the viral and cell membrane. It is highly conserved among corona viruses and S2-targeting neutralizing antibodies may inhibit formation of six-helix bundle structure and consequently block membrane fusion (2,25,27). Nucleocapsid (NP), located inside the virion, is responsible for RNA packaging and virus replication. It might be important in modulating antiviral immunity and inhibiting interferon production (6).
Neutralizing efficacy relies on binding position and affinity of antibodies (25). SARS-CoV-2 variants with mutations in RBD and NTD exhibited partial resistance to neutralizing antibodies generated by natural infection or vaccination (15). Delta variant (B.1.617.2), containing diverse mutations in the RBD and NTD, was resistant to neutralization by some anti-NTD and anti-RBD monoclonal antibodies and convalescent sera from COVID-19 patients (16).
The duration of protective immunity to SARS-CoV-2 plays a pivotal role in rates of reinfection and vaccine breakthrough and hence the overall trajectory of the pandemic. Neutralizing antibody dropped rapidly in the first 2–3 months after infection and slower thereafter (8,13). Although NP and S-specific immunoglobulin G (IgG) titers declined, the potent neutralizing activity sustained for up to 12 months after natural infection in ∼20% of patients (23,28). Further investigations in the long-lasting antibody of COVID-19 patients might provide important insights leading to novel approaches in vaccine design.
In this study, we characterized longitudinal trend of binding antibodies to several domains of SARS-CoV-2 that may be responsible for the neutralizing activities in COVID-19 recovered patients. Microneutralization of SARS-CoV-2 and enzyme-linked immunosorbent assay (ELISA) analysis of binding antibody were tested in 307 convalescent sera that were sequentially collected from 35 patients from 2 to 12 months after recovery. Binding levels of the sera to SARS-CoV-2 RBD, NTD, S2, and NP antigens were determined and compared with neutralizing titers.
Materials and Methods
Serum samples
The study included 307 sera from 35 COVID-19 recovered patients during February 2020 to April 2021 provided by Thai Red Cross Society. The blood collection was part of the activity to prepare convalescent hyperimmuned plasma for COVID-19 treatment. For negative control, sera collected prior the SARS-CoV-2 pandemic (in 2010) were used. The study was approved by the ethic committees of the Thai Red Cross and the Faculty of Medicine Siriraj Hospital.
Live-virus microneutralization assay
Live-virus microneutralization of SARS-CoV-2 using a cell-based indirect ELISA detecting NP of virus was performed in a certified Biosafety Level 3 facility at the department of microbiology, faculty of science, Mahidol University. The experimental protocols were performed following standard protocols approved by the biosafety committee of Mahidol University.
SARS-CoV-2 viruses (SARS-CoV-2/01/human/Jan2020/Thailand) representing the original Wuhan strain isolated from a confirmed COVID-19 patient at Bamrasnaradura Infectious Diseases Institute, Nonthaburi, Thailand, and the B.1.617.2 delta variant (SARS-CoV-2/human/THA/VTM1_P2/2021) were used for in vitro experiment. In brief, heat-inactivated serum was twofold serially diluted in culture medium starting at 1:10 and 60 μL and mixed with 60 μL (100 the median tissue culture infectious dose [TCID50]) SARS-CoV-2 for 1 h. The mixture was then added to Vero-E6 cells and incubated for 1 h. The cells were washed and further incubated in medium for 48 h.
Cytopathic effect was observed under microscope and cells were fixed with 1:1 cold methanol–acetone for 20 min. The fixed cells were stained using SARS-CoV-2 NP monoclonal antibody (Sino Biological) and a secondary peroxidase-labeled goat antirabbit IgG (Dako). Signal was developed using 3,3′,5,5′-tetramethylbenzidine (TMB) substrate (Supplier of TMB reagent [KPL]) and color was halted by adding 1 N HCl. The plate was read by an ELISA reader at wavelength of 450 and 620 nm. The average optical density (OD) 450/620 is determined for quadruplicate wells of 100 TCID50 and negative control wells (CC), and a neutralizing endpoint is determined by using a 50% specific signal calculation. The endpoint titer was expressed as the reciprocal of the highest dilution of serum with average OD450/620 value (duplicate wells) less than X. X = [(average OD of 100TCID50) – (average OD of CC)]/2 + (average OD of CC).
ELISA of antibody binding to SARS-CoV-2 spike and NP antigens
SARS-CoV-2 RBD, NTD, S2, or NP protein (Sino Biological) at 0.5 μg/mL was coated on a MaxiSorp Nunc-immuno 96-well plate overnight at 4°C. Wells were blocked with blocking buffer (5% bovine serum albumin and 0.1% Tween-20 in phosphate-buffered saline [PBS]) for 1 h at room temperature, followed by incubation with 1:2,000 diluted sera in blocking buffer for 1 h at 37°C. Wells were washed four times with washing buffer (0.1% Tween-20 in PBS). A 1:5,000 dilution of horseradish peroxidase-conjugated goat antihuman IgG antibody (Thermo) was added for 1 h at 37°C. Wells were washed four times with washing buffer and developed using KPL TMB microwell peroxidase substrate (Seracare) for 10 min at room temperature. The reaction was stopped by adding 1 M H2SO4. Absorbance was read at 450 and 630 nm using microplate reader.
Results
We followed 35 individuals with confirmed history of COVID-19 who were willing to donate blood regularly and had neutralizing antibody titers over 160 at the first blood collection. Demographic data, onset duration before blood donation, symptoms, and severity of individuals are given in Table 1.
Demographic Data, Onset Duration Before Blood Donation, Symptoms, and Severity of 35 Donors with Confirmed History of COVID-19 Who Had Neutralizing Antibody Titers to SARS-CoV-2 Over 160 at the First Blood Collection
F, female; ICU, intensive care unit; M, male; NT, neutralization assay.
Blood samples were collected every 2–3 weeks and tested for neutralizing antibody titers, and the sequential blood collection was terminated once the neutralizing antibody titer declined to <160 since the sera could not be used for treatment. Figure 1 shows patterns of neutralizing antibody titers and binding antibody levels of RBD, NTD, S2, and NP of the individuals. Sera from donors shown in Figure 1A exhibited similar trend between neutralizing titer and binding level of RBD, NTD, S2, and NP that mostly decreased over time. The neutralizing antibody titers decreased to <160 within 3 months after the first blood collection in nine donors. Another three donors Nos. 4, 10, and 27 had neutralizing antibody titers <160 after 23, 13, and 33 weeks, respectively.
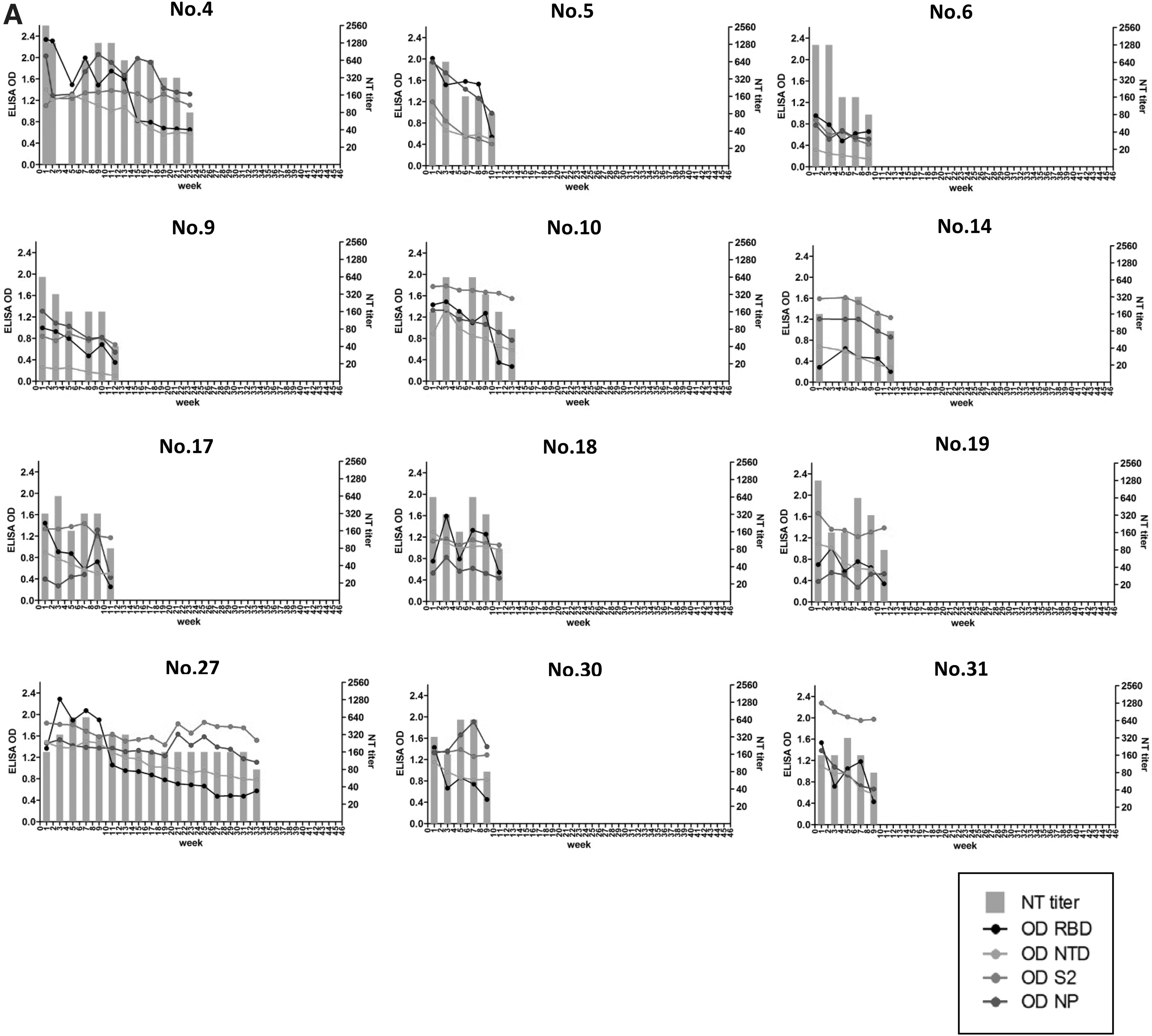
Comparison of neutralizing antibody titers and binding antibody levels of RBD, NTD, S2, and NP of sequentially collected sera from 35 donors.
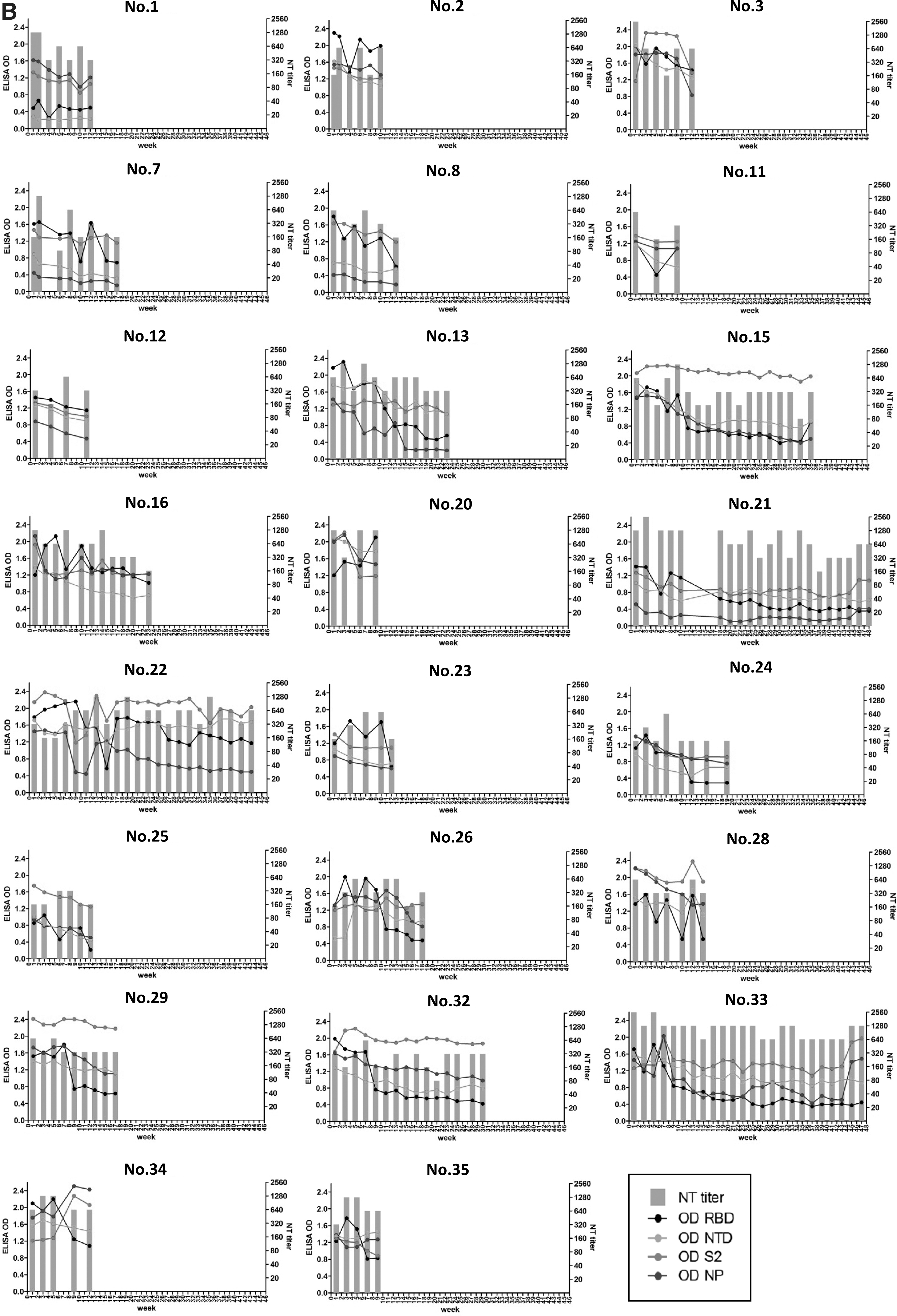
There were 10 donors with high neutralizing antibody titers shown in Figure 1B but they were lost to follow-up after 2 months, leaving 13 donors with sequential antibody titer data for 3–12 months. These 13 donors showed persistently high neutralizing titer for >3 months. Interestingly, donor Nos. 21 and 33 still had high neutralizing titer at 640 and 1,280, respectively, at 1 year after the first collection.
As RBD is generally a major target of SARS-CoV-2-neutralizing antibodies, similar trends between neutralizing titers and binding levels of RBD were observed in donor Nos. 1, 2, 7, 8, 11, 22, 23, 28, 34, and 35. However, donor Nos. 13, 15, 21, 24, 26, 29, 32, and 33 had persistently high neutralizing titer (≥320), but their binding levels to RBD rapidly decreased. This decline of the RBD binding antibody levels in these subjects was faster than the decline of antibody to the other domains. As time passed, the dramatically decreasing of binding antibody to RBD continued while neutralizing titers were still maintained. This suggested that other epitopes were also crucial.
Binding antibody level of NTD mostly decreased over time except sera of donor Nos. 22, 26, 28, 29, and 34, which corresponded to the persistently high neutralizing titer. However, donor Nos. 1 and 9 always had low binding level of NTD since the first collection. Their neutralizing antibodies may target NTD much lesser than other epitopes. Although binding antibody level of NP usually dropped over time, donor Nos. 27, 33, and 34 demonstrated similar trends between NP and S2, which also corresponded to their neutralizing titers.
Surprisingly, binding antibody specific to S2 was always stable or slowly decreased in most of donors in this study. Donor Nos. 15, 22, and 33 clearly demonstrated high and long-lasting neutralizing titer >640. These donors also showed high and persistent levels of binding antibody to S2. We also tested neutralizing titers against a delta variant in selected samples from subjects with rapidly declining and persisting neutralizing antibody at early and the latest time point with high neutralizing titers to see whether persistent S2 antibody provided any cross-neutralization against the variant.
We found that these sera with high neutralizing titers and high S2-binding antibody did not show significantly higher cross-neutralization to the delta variant than sera with lower S2-binding antibody (Table 2). This suggested that the persistent neutralizing activity mainly targeted variable S1 epitopes despite the low binding antibody to the NTD and RBD.
Neutralizing Titer and Binding Antibody Level of Receptor Binding Domain, S2, Nucleocapsid, and N-Terminal Domain Epitope
ELISA, enzyme-linked immunosorbent assay; NP, nucleocapsid; NTD, N-terminal domain; RBD, receptor binding domain.
Overall, the antibody response to each epitope can be differently maintained over time after infection. The persistent neutralizing titer despite the low level of binding antibody to RBD was probably from the specific binding antibodies to non-RBD epitopes such as S2, NP, and even quaternary epitopes. For natural infection, RBD seems to be mainly responsible in earlier periods, and other epitopes may play roles for neutralizing activities in later periods.
Discussion
In general, immune protection against COVID-19 can be implied by the level of neutralizing antibody against the virus. Among all of the SARS-CoV-2 proteins, RBD of S1 is the major target for the development of potent neutralizing antibody (5). Escape variants carrying mutations and deletions in RBD and NTD could reduce sensitivity to antibody neutralization. For instance, highly concerned mutations in RBD including E484K and K417N/T in B.1.351 (Beta) and P.1 (Gamma) variants were reported that they partially impair neutralization generated by previous infection or vaccination (15,26). In addition, neutralizing antibody titer of most of the convalescent sera correlated with level of specific binding antibody to RBD. Thus, many anti-RBD assays were developed to use as a predictor of the neutralization capability (7,9,12).
Nevertheless, SARS-CoV-2 spike protein contains other important antigenic sites. Monoclonal antibodies isolated from COVID-19 patients were potent and diverse, and there were monoclonal antibodies targeting RBD, NTD, and quaternary epitope on top of the spike (10). Besides RBD region, NTD and S2 might contribute to the neutralization as well (5,27). S1-targeting monoclonal antibody 4A8 isolated from COVID-19 recovered patients demonstrated that it did not block the interaction between ACE2 and S protein, but exhibits high levels of neutralization against both authentic and pseudotyped SARS-CoV-2. This monoclonal antibody was proved to bind NTD (3). Binding antibodies to NTD, S2, and NP also corresponded to the neutralizing titers.
In contrast, some donors who still had high neutralizing titer for the longer period at 3–12 months showed a discrepancy between the stable neutralizing titers and the declining levels of RBD, NTD, and NP-binding antibodies. Only capability. However, a study showed that immunoglobulin M at days 21 and 42 preferentially recognized epitopes on S2, followed by NTD and limited binding to epitopes in RBD (18).
Antibodies to spike reached a peak by 5–8 weeks and then declined, whereas those to S2 of seasonal beta coronaviruses continued to rise and correlated significantly with neutralizing antibodies (4,21). In fact, S2 is more conserved among coronaviruses than S1. The high and stable level of anti-S2 is probably from anamnestic boost of cross-reactive neutralizing antibodies against S2 that pre-exist in populations since previous exposures to human coronaviruses (19). This cross-reactive antibody epitope in S2 was also suggested to reduce the severity of COVID-19 (22).
Conclusion
Antibody responses to SARS-CoV-2 recognize several epitopes of the virus. Antibodies to different epitopes could have different rate of decline over time, and their contribution to neutralizing activity could change. Although RBD is such a dominant epitope that can instantly elicit potent neutralizing antibodies, anti-RBD might not be long lasting. The mechanism governing the differential longevity of different antibodies is not well understood and further studies into the longevity of specific antibody may help us improve vaccine design to induce long-lasting immunity to COVID-19.
Footnotes
Authors' Contributions
S.P. contributed to formal analysis (lead), investigation (lead), methodology (lead), visualization (equal), and writing—original draft (lead). S.M. was involved in data curation (lead), investigation (equal), methodology (equal), and resources (equal). C.B. was in charge of project administration (lead), validation (lead), data curation (equal), investigation (equal), and methodology (equal). P.K. carried out data curation (equal), investigation (equal), and resources (equal). D.C. was in charge of resources (lead) and supervision (equal). T.R. was in charge of resources (equal) and validation. P.A. took care of conceptualization (lead), funding acquisition (lead), supervision (lead), and writing—review and editing (lead). A.T. was involved in visualization (lead), conceptualization (equal), resources (equal), supervision (equal), and writing—review and editing (supporting).
Compliance with Ethical Standards
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by National Science and Technology Development Agency (P-20-50640), Thailand, and partly supported by Faculty of Medicine Siriraj Hospital, Mahidol University, Thailand.