
Abstract
Dendritic cells (DCs), as the most powerful antigen-presenting cells, play a key role in the adaptive immune response, while the defective function of DC is an important factor in immune tolerance to hepatitis B virus (HBV) infection. Hepatitis B virus core antigen (HBcAg) is a highly antigenic protein that can induce a strong antigen-specific immune response against HBV. In this study, we first constructed the ubiquitinated HBcAg gene (UbHBcAg), and then utilized a recombinant lentiviral vector UbHBcAg (LV-UbHBcAg) to explore the role of them in DC autophagy and function. Meanwhile, the effects of autophagy on DC functional activation were further analyzed. Finally, we investigated the underlying mechanism of autophagy induced by LV-UbHBcAg. Results showed that LV-UbHBcAg could promote autophagic progression in DCs, and the upregulated autophagy can further enhance DC functional maturation. In addition, p62 may serve as an important role in autophagy degradation. More importantly, the PI3K/Akt/mTOR signaling pathway was involved in the process of autophagy induced by LV-UbHBcAg. These findings suggest that LV-UbHBcAg can activate DC function by inducing autophagy, which may represent a promising strategy to treat chronic HBV infection.
Introduction
Public health awareness of chronic hepatitis B virus (HBV) infection has progressively increased in recent years (Spyrou et al, 2020). Dendritic cells (DCs), as the most powerful antigen-presenting cells that internalize antigens and present antigen-derived peptides to T cells, play a central role in the activation of innate and adaptive immune responses against invading HBV (Chen and Tian, 2019; Meng et al, 2019; Nomura et al, 2018). Therefore, we have considered that the recovery of DC function suppressed by HBV could be a useful immunological therapy to eliminate HBV infection (Ciupe, 2018).
Studies have shown that loading virus-related antigens with lentiviral vectors and immunizing the body can effectively induce antigen-specific immune responses (Annoni et al, 2019; Magrin et al, 2019; Tora et al, 2020). Hepatitis B virus core antigen (HBcAg), an immunostimulatory protein, can induce strong antigen-specific Cytotoxic T Lymphocyte (CTL) responses in HBV-infected patients (Akbar et al, 2012). Therefore, using HBcAg as a target antigen to promote DC function is of great significance in the treatment of chronic HBV infection.
Autophagy, as a fundamental intracellular process, is responsible for the lysosomal degradation of certain microorganisms (viruses, bacteria, fungi, and protists) (Lennemann and Coyne, 2015; Zang et al, 2016) and damaged cell organelles and proteins (Yu et al, 2018). Appropriate levels of autophagy can facilitate the recycling of nutrients, balance cellular metabolism, and promote cell functions. In recent years, autophagic activities have been considered a necessary part of their ability to activate DCs (Ghislat and Lawrence, 2018). Optimizing antigen processing and antigen presentation in DCs by autophagic machinery can significantly orchestrate T cells and then enhance immune responses to inhibit HBV infection (Valdor et al, 2014).
Mammalian target of rapamycin (mTOR), a critical regulator of various cell signaling pathways and nutrient and hormonal signal processes, maintains the balance between cell apoptosis and autophagy in response to cellular stress (Wang and Le, 2018). mTOR inhibition could activate overall protein degradation by the ubiquitin proteasome system and autophagy process (Wang et al, 2019). Previous reports have indicated that mTOR is activated by the PI3K/Akt signaling pathway and that the PI3K/Akt/mTOR signaling pathway is critically involved in orchestrating the metabolic demands necessary for DC activation (Wang et al, 2019; Yue et al, 2019).
In this study, we investigated the effects of lentiviral vector carrying ubiquitinated HBcAg (LV-UbHBcAg) on DC function and autophagy activation. Moreover, the PI3K/Akt/mTOR signaling pathway may mediate the autophagy progress induced by LV-UbHBcAg. Our finding provides a further understanding of the impact of LV-UbHBcAg on DC function and may ultimately benefit the development of new DC-based immunomodulatory strategies for chronic hepatitis B (CHB).
Materials and Methods
Animals, reagents, and antibodies
C57BL/6 (B6) mice (6–8 weeks old, weighing 20–25 g) were purchased from Chinese Academy of Sciences (Shanghai, China). The study was approved by the Shanghai Sixth People's Hospital Ethics Committee. The HBcAg and ubiquitinated HBcAg (UbHBcAg) genes were amplified by polymerase chain reaction (PCR). The primers used were as follows: HBcAg forward: 5′-AACCCCGGTCCGGCTAGCGCCACCATGGACATTGACCCGTATAAAG-3′ and reverse: 5′-GCCAGATCCGCCGGATCCACATTGAGATTCCCGAGATTGA-3′, and UbHBcAg forward: 5′-AACCCCGGTCCGGCTAGCGCCACCATGCAGATCTTCGTGAAGACCC-3′ and reverse: 5′-GCCAGATCCGCCGGATCCACATTGAGATTCCCGAGATTGA-3′ (Dai et al, 2016). Antibodies against Bim, Bax, PI3K, and p-PI3K were obtained from Cell Signaling Technology (Boston, MA). Antibodies against Akt, p-Akt, mTOR, p-mTOR, P70S6K, p-P70S6K, HBcAg, LC3, Beclin-1, p62 and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) were purchased from Abcam (Cambridge, UK), and fluorescent antibodies were obtained from BD Biosciences (San Diego, CA). Proteasome inhibitor (MG132), chloroquine (CQ), 3-methyladenine (3-MA), and rapamycin (Rapa) were purchased from Sigma–Aldrich (St. Louis,MO).
Generation of bone marrow-derived DCs and cell culture
Mouse bone marrow-derived DCs (DCs) were harvested from C57BL/6 mice and generated according to the protocol described by Ma et al (2018). These DCs were seeded in complete RPMI 1640 medium at a density of 2 × 106 cells/mL with the addition of 10% fetal bovine serum (FBS; Gibco, USA), 10 ng/mL murine IL-4 (mIL-4; PeproTech, Rocky Hill, NJ), and 20 ng/mL murine granulocyte-macrophage colony-stimulating factor (mGM-CSF; PeproTech); mGM-CSF, mIL-4, and fresh RPMI 1640 were supplemented on alternate days. DC2.4 cells were purchased from the American Type Culture Collection (Manassas, VA). All the above cells were cultured at 37°C with 5% CO2 in RPMI 1640 or Dulbecco's modification of eagle's medium dulbeo (DMEM) supplemented with 10% FBS, 100 U/mL penicillin, and 100 mg/mL streptomycin (Li et al, 2019).
Transfection of DCs
DCs were generated as described above, and the 2nd day of isolation and culture of marrow-derived DCs was defined as the first experimental day (day 1). On day 5, immature DCs were seeded in a 6-well plate (2 × 106/well) or a 24-well plate (2 × 105/well) and then cultured at 37°C with 5% CO2. LV-UbHBcAg, LV-HBcAg, and LV were transfected into DCs at an multiplicity of infection (MOI) of 40, and phosphate buffered solution (PBS) was used as a control. After 24 h, the supernatant was harvested, and fresh RPMI 1640 containing 10% FBS was added. The transfected cells were cultured for an additional 3 days and analyzed for green fluorescent protein expression with a Coulter Epics XL flow cytometer (Beckman, Miami, FL).
DC immunophenotyping and cytokine production
On day 8, the supernatant was collected, and the levels of different secreted cytokines (IL-12p70 and TNF-α) were measured using ELISA kits (R&D Co., Ltd., Minneapolis, MN) according to the manufacturer's instructions. For each assay, all samples were analyzed in three duplicates. On day 9, mature DCs loaded with LV-UbHBcAg, LV-HBcAg, and LV were harvested and DCs treated with lipopolysaccharide (LPS; 1 μg/mL, 12 h; Sigma-Aldrich) were used as a control group. These cells were incubated in cold buffer and subsequently stained for 30 min with the following allophycocyanin (APC)- or Phycoerythrin-labeled monoclonal antibodies: CD11c, CD80, CD83, CD86, and major histocompatibility complex (MHC)-II (BD Biosciences). The stained cells were analyzed by flow cytometry.
Western blotting
The above DCs were collected in the presence/absence of Rapa, 3-MA, CQ, and MG132. The expression of autophagy-related proteins, PI3K/Akt/mTOR pathway-related proteins, and apoptosis proteins was determined by Western blot according to standard protocols. The above corresponding primary and secondary antibodies were used. The signals were detected with an ECL assay kit (Amersham). The Western blots shown are representative of at least three independent experiments.
Transmission electron microscopy
DCs in the LV-UbHBcAg and Rapa groups were treated with the indicated concentrations of triptolide for 24 h and collected by trypsinization. Cells were fixed in 2.5% phosphate-buffered glutaraldehyde at 4°C overnight and postfixed in 1% phosphate-buffered osmium tetroxide for 1.5 h at room temperature. After being embedded and double stained with uranyl acetate/lead citrate, the sections were analyzed under an H-7650 transmission electron microscope (Hitachi) at 60 kV.
Assessment of apoptosis
DCs in each group were washed twice with PBS, and 7-amino-actinomycin D and Annexin V-APC staining (eBioscience) were performed according to the manufacturer's instructions. Triple-stained positive cells among the whole cell population were detected by flow cytometry.
Fluorescence microscopy analysis
DC2.4 cells were placed in a 12-well plate (NEST, China) and later loaded with LV-UbHBcAg at an MOI of 40 for 72 h. Then the cells were washed with PBS and fixed with 4% paraformaldehyde for 20 min at room temperature. Thereafter, the cells were permeabilized with 0.1% Triton X-100, preincubated with 5% Albumin from bovine serum, and washed thrice with PBS. The cells were incubated overnight at 4°C with 1:500 rabbit p62 antibody and secondary antibodies (DyLight 549; Novus Biologicals) in the dark for 1 h. Finally, the cell nuclei were stained with 4′,6-diamidino-2-phenylindole dihydrochloride (Beyotime, Nanjing, China) for 5 min followed by three washes with PBS. Representative images were captured using a BX63 automatic fluorescence microscope (Olympus, Japan).
Real-time quantitative PCR analysis of mTOR
RNA was extracted from DCs by TRIzol (Invitrogen) according to a standard procedure. Real-time PCR was performed using SYBR Green I (Eurogentec, Seraing, Belgium) and Multiscribe Reverse Transcriptase (Applied Biosystems, Foster City, CA) and subsequently monitored using an ABI Prism 7900 Sequence Detection System (Applied Biosystems). Amplification of GAPDH transcription was performed as a control. The mTOR primers were previously reported (Liu et al, 2019). Primer sequences for each complementary DNA were designed using either Primer Express Software (Applied Biosystems) or Primer depot (
Statistical analysis
All values in the text and figures were obtained from three independent experiments. Data are presented as the mean ± standard deviation. GraphPad Prism V7.0 (San Diego, CA) was used to perform statistical analyses. Differences between two groups were analyzed using Student's t test, and the differences between two or more groups were analyzed using one-way analysis of variance. Differences were considered significant at p < 0.05.
Results
Autophagy is notably induced in DCs loaded with LV-UbHBcAg
To investigate the induction of autophagy by LV-UbHBcAg, we first examined the protein levels of autophagy-related membrane structures, such as LC3, Beclin-1, and p62. Reduced expression of p62 and increased levels of LC3 and Beclin-1 usually indicate enhanced autophagic flux. As shown in Figure 1A and B, Western blot analysis revealed noticeably increased levels of LC3 and Beclin-1 in the LV-UbHBcAg group compared with the normal contrast (NC) and LV groups. Moreover, decreased expression of p62 was observed. Then, to further verify autophagy induction by LV-UbHBcAg, transmission electron microscopy analysis was performed to assess the formation of autophagosomes.
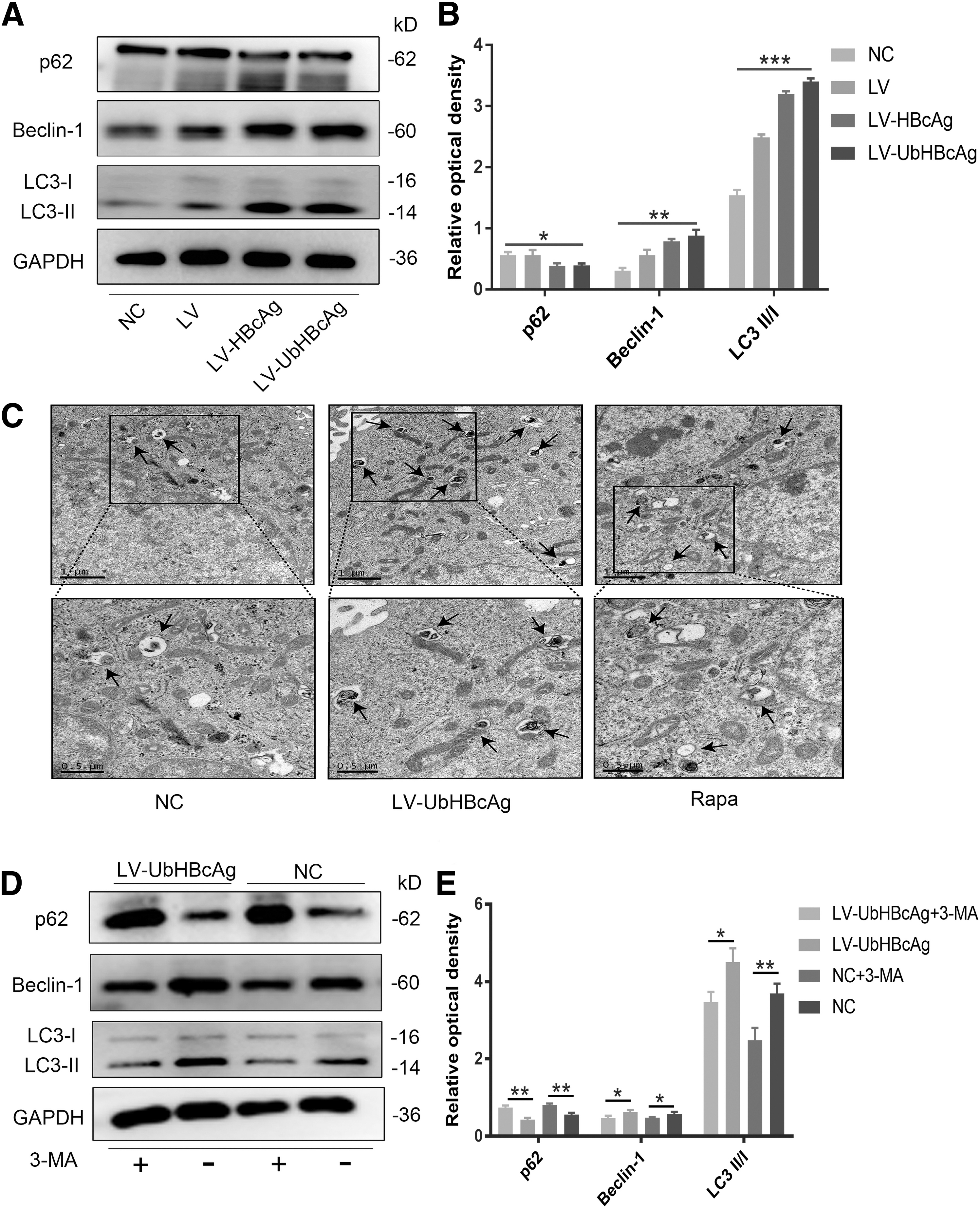
Activation of autophagic flux in DCs loaded with LV-UbHBcAg. DCs were transduced with LV, LV-HBcAg, and LV-UbHBcAg for 72 h.
As shown in Figure 1C, compared with the NC group, many characteristic double-membrane autophagic vesicles that contained engulfed organelles were observed in the LV-UbHBcAg group, which showed even more vesicles than the positive control group (Rapa). In this study, to explore whether autophagy activation can be inhibited by the autophagy inhibitor 3-MA, DCs were treated with 3-MA (10 μM) for 24 h. As shown in Figure 1D and E, 3-MA significantly reduced the protein expression of LC3 and Beclin-1, which are required for autophagic flux induction.
The effects of LV-UbHBcAg on DC maturation and functional activation
Autophagy is recognized as an important factor in DC maturation and function (Sugawara et al, 2020). To investigate whether LV-UbHBcAg plays a role in DC maturation, we detected the levels of costimulatory molecules and the secretion of cytokines. In this study, LPS treatment was included as a positive control, and the NC group was the negative control. First, flow cytometry analysis showed that both LV-UbHBcAg and LV-HBcAg significantly elevated the expression of costimulatory molecules (CD80, CD80, and CD86) compared with the NC group (Fig. 2A, B). Similarly, the expression of MHC class II (MHC-II) in the LV-UbHBcAg group was significantly higher than that in the NC group (Fig. 2C, D). Moreover, we found that the effects of LV-UbHBcAg to promote DC maturation were stronger compared with LV-HBcAg.
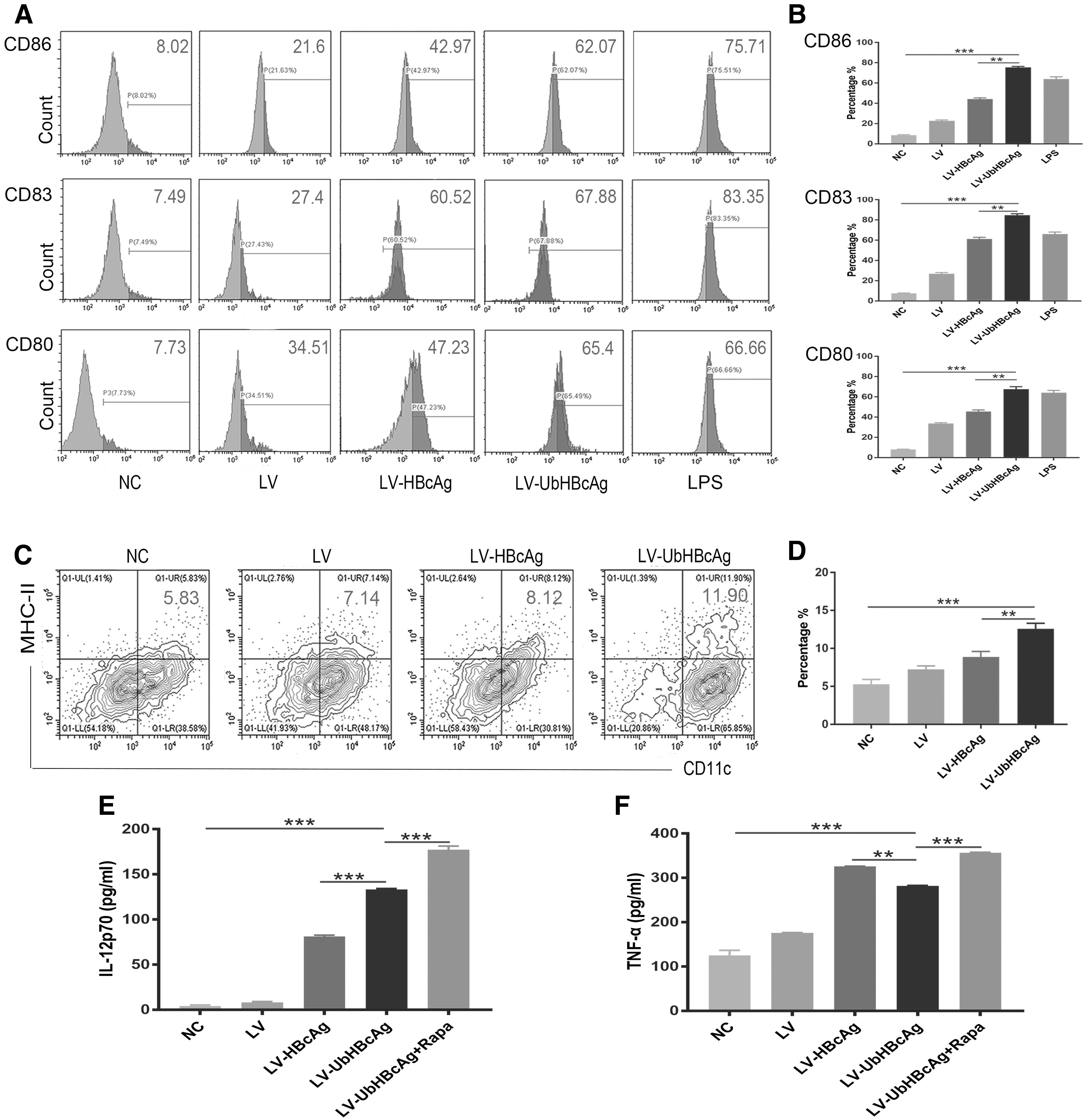
LV-UbHBcAg promoted DC maturation and immune function.
DCs, as immune regulatory cells, participate in cellular immune responses by secreting different types of cytokines, affecting the process and outcome of adaptive immune responses. First, we discussed the effects of LV-UbHBcAg on cytokine secretion in DCs, and the levels of cytokines (IL-12p70 and TNF-α) in the cell culture supernatant were detected by ELISA. The results showed that LV-UbHBcAg-loaded DCs expressed significantly higher amounts of IL-12p70 and TNF-α than the NC group of DCs (p < 0.05; Fig. 2E, F). In addition, exposure of DCs to rapamycin (an autophagy inducer) in the presence of LV-UbHBcAg resulted in the highest levels of IL-12p70 and TNF-α.
The overall results indicated that LV-UbHBcAg can promote DC maturation, and with the enhancement of autophagy, LV-UbHBcAg-loaded DCs have a stronger effect on DC functional activation. It is worth noting that all the above results indicate the effects of LV-UbHBcAg are stronger than those of LV-HBcAg. The possible reason for this result is that with the participation of the ubiquitin molecule (Ub), HBcAg can be rapidly degraded by the ubiquitin–proteasome system (UPS); however, it is unclear how the autophagic program is involved. Next, we will continue to examine the role of autophagy.
Autophagy mediates the degradation of UbHBcAg
Eukaryotic cells mainly rely on two protein degradation mechanisms, the autophagy–lysosome system and the UPS, to complete the metabolism of intracellular proteins (Singh et al, 2021). Among them, the UPS mainly removes soluble small molecule proteins, while the autophagy–lysosome system is responsible for polymers and damaged organelles. P62, known as a classical receptor of autophagy, serves as a link between LC3 and ubiquitinated proteins in the autophagic machinery (Zaffagnini et al, 2018). In the process of autophagy–lysosome degradation, p62 is degraded by hydrolytic enzymes along with the degradation of ubiquitinated proteins. In this study, we assume that a reduced protein level of p62 can reflect the degree of degraded proteins.
To explore the role of autophagy in UbHBcAg degradation, we next analyzed the expression of HBcAg, LC3, and p62 in DCs loaded with LV-UbHBcAg. As shown in Figure 3A and B, treatment with the proteasome inhibitor MG132 slightly increased the expression of p62, while apparently upregulating the level of LC3 and HBcAg in DCs, which indicates that MG132 may not strongly affect the levels of p62, but can reduce HBcAg degradation. Interestingly, in Figure 3C and D, CQ treatment noticeably increased the expression of HBcAg in DCs, suggesting that when the autophagic process was inhibited, the degradation of HBcAg was suppressed as well.
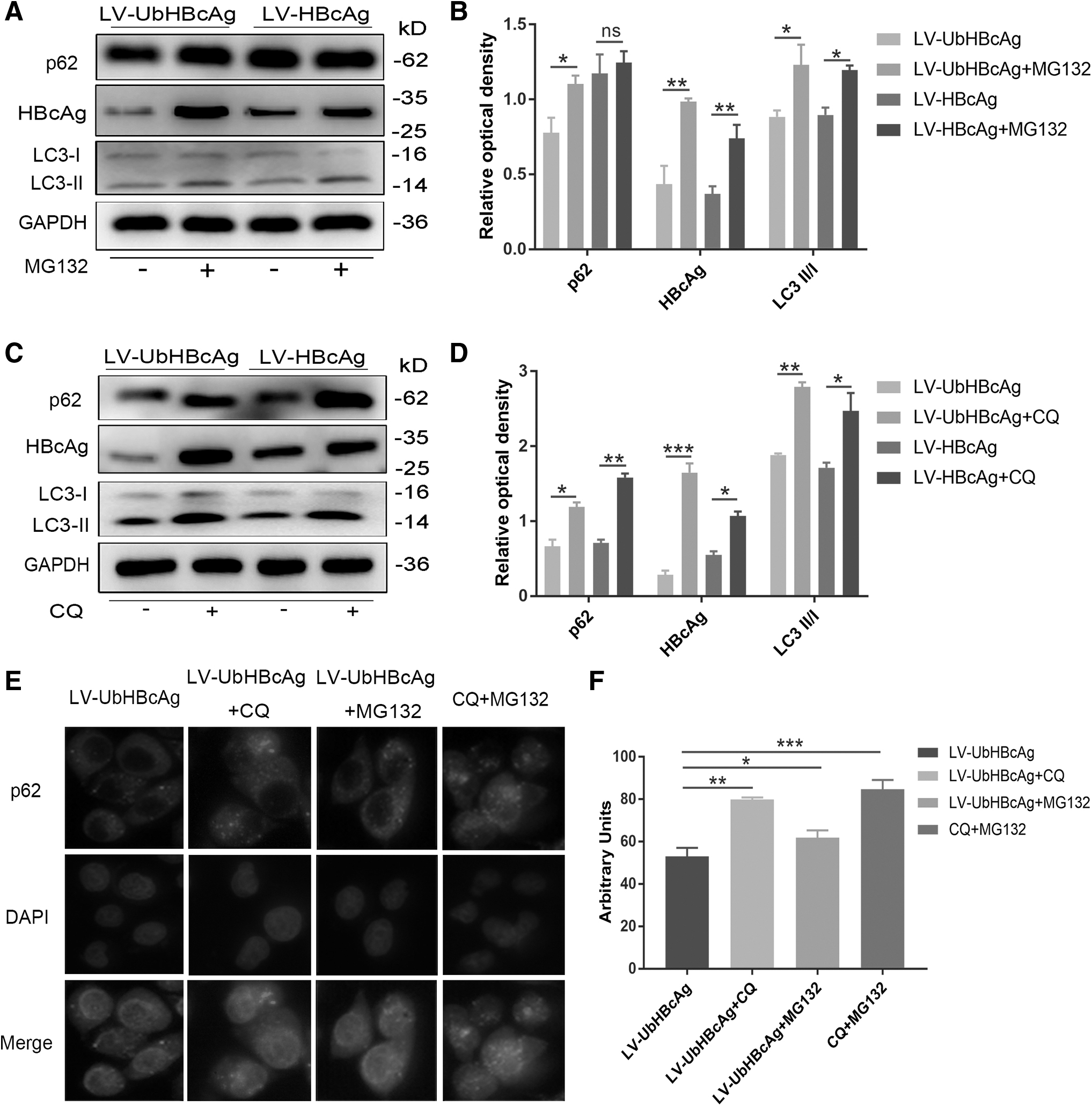
Autophagy mediated the degradation of UbHBcAg.
In addition, the reduction in HBcAg in the LV-UbHBcAg group was greater than that in the LV-HBcAg group. More importantly, the level of p62 and HBcAg proteins has a consistent change trend. To further investigate the role of p62 in UbHBcAg degradation, we next analyzed the intracellular staining of p62 in DCs loaded with LV-UbHBcAg. LV-UbHBcAg was used to transfect DC2.4 cells (immortalized cells morphologically and functionally similar to bone marrow-derived dendritic cells, which allowed the cells to stably express UbHBcAg. In Figure 3E and F, immunofluorescence images showed that the presence of CQ or MG132 resulted in a significant accumulation of p62-positive punctate structures in DCs. With the combined effects of MG132 and CQ, the expression of p62 in the cytoplasm was higher than that in untreated cells.
The overall results collectively suggest that both the UPS and autophagy could mediate the degradation of UbHBcAg and that p62 may serve as an important factor in the degradation process. Indeed, we cannot clearly explain the specific mechanism of UPS combined with autophagy to mediate the degradation of ubiquitinated proteins, which is an issue that needs more research.
Upregulated autophagy affects DC apoptosis
Given that the role of autophagy in cell survival and death is controversial (Feng et al, 2018), we further explored the effect of the autophagic process on cell apoptosis. As shown in Figure 4A and B, Western blot analysis identified significantly increased expression of the proapoptotic proteins (Bim and Bax) in the LV-UbHBcAg group compared with the NC group, which maintained a level similar to that of the LV-HBcAg group.

Upregulated autophagy contributed to DC apoptosis.
To further explore the effect of autophagy on cell survival, we next examined the percentages of DCs undergoing early apoptosis by flow cytometry analysis. Figure 4C and D show that the early apoptotic cell ratio was significantly decreased in LV-UbHBcAg-treated DCs compared with LV-treated DCs. Unfortunately, we found that the proportion of apoptotic cells in the above two groups was higher than that in the NC group. We speculate that this is because lentiviruses are cytotoxic and cause cell apoptosis. We next investigated whether the promotion of autophagy was associated with the modulation of apoptosis in DCs loaded with LV-UbHBcAg. With 3-MA treatment, the number of apoptotic cells in the LV-UbHBcAg group was significantly reduced. The above results indicate that the lentivirus can cause cell apoptosis; moreover, the upregulated autophagic process induced by LV-UbHBcAg also causes the death of DCs.
The PI3K/Akt/mTOR signaling pathway is involved in autophagy induced by LV-UbHBcAg
Previous studies have indicated that the PI3K/Akt/mTOR signaling pathway is essential for the regulation of autophagy (Qiao et al, 2020; Varshney and Saini, 2018). To further determine the potential mechanism of autophagy induced by LV-UbHBcAg, we next investigated the protein levels of PI3K/Akt/mTOR signaling pathway-related proteins. Among them, mTOR, as a central regulator of cell growth, can negatively regulate autophagy activity. As demonstrated in Figure 5A and B, when DCs were treated with LV-UbHBcAg, the expression of total and phosphorylated PI3K, Akt, and mTOR proteins significantly decreased compared with the NC group. In addition, P70S6K, as a downstream target of mTOR, did not show a decline, while phosphorylated P70S6K was clearly downregulated.
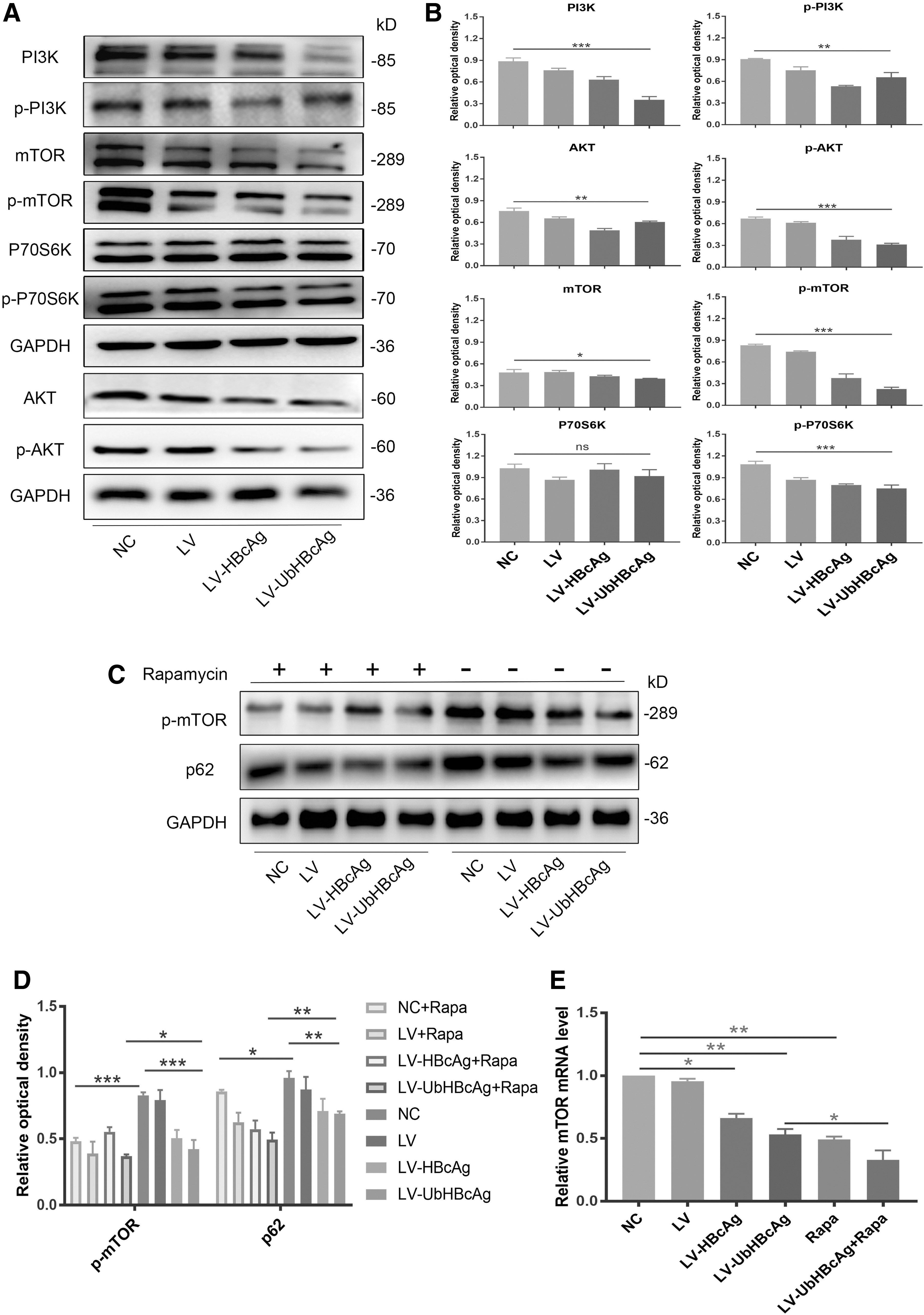
Autophagy induced by LV-UbHBcAg was relevant to the inhibition of the PI3K/Akt/mTOR signaling pathway.
In view of the regulatory effects of mTOR on autophagy, after treatment with the mTOR-specific inhibitor rapamycin, we evaluated the protein level of phosphorylated mTOR in DCs. In agreement with other works, rapamycin treatment resulted in downregulated levels of phosphorylated mTOR and p62 (Fig. 5C, D). In parallel with the protein analysis, the real-time quantitative PCR data also showed decreased expression of mTOR mRNA in DCs loaded with LV-UbHBcAg (Fig. 5E), which means that LV-UbHBcAg can inhibit the expression of mTOR. Thus, our results suggested that LV-UbHBcAg can significantly modulate mTOR protein expression and the PI3K/Akt/mTOR signaling pathway may be involved in the progression of autophagy activated by LV-UbHBcAg.
Discussion
DCs are essential for the initiation of antigen-specific immune responses through antigen presentation, which is enhanced by autophagy activation (Ghislat et al, 2018; Ho et al, 2021; Zang et al, 2016). We and others have previously shown that autophagy significantly affects T cell and DC functions, with important physiological and pathological implications in the immune response (Eisenbarth, 2019; Ma et al, 2018). Indeed, we have previously indicated that LV-UbHBcAg stimulates robust HBV-specific CTL immune responses (Dai et al, 2015; Ma et al, 2018; Tan et al, 2017). The purpose of this study was to investigate whether LV-UbHBcAg-treated DCs may undergo a proautophagic program. We report that LV-UbHBcAg can induce autophagy by inhibiting the PI3K/Akt/mTOR signaling pathway in DCs. Furthermore, under the effects of autophagy, LV-UbHBcAg is beneficial for DC maturation and functional activation, which promote strong immune responses.
A growing number of studies have demonstrated that autophagy is central to the development of an efficient balanced immune response (Boya, 2021). The cellular autophagic machinery breaks down damaged proteins and organelles by sequestering or directing cargo to the lysosome (Deretic and Kroemer, 2022). This same machinery is required for DCs to fight invading pathogens and to shape cellular immunity through antigen processing, costimulatory molecule expression, and cytokine secretion. Autophagy is a very complex process that has been associated with DC functions (Wculek et al, 2019).
Prior studies show that autophagy is involved in several biological processes, including DC maturation, cytokine production, migration, responses to Toll-like receptor stimulation, and T-cell activation (Parekh et al, 2017; Qiao et al, 2020). Moreover, autophagy-related proteins can control the homeostasis and function of antigen cross-presentation in DCs. In our study, we found that LV-UbHBcAg promoted autophagy in DCs, and the formation of autophagosomes was enhanced. Furthermore, we observed a significant increase in the levels of several cytokines, such as TNF-α and IL-12p70, which contributed to DC function promotion. In addition, with the participation of Ub, the effect of LV-UbHBcAg on DC activation was stronger compared with LV-HBcAg.
Autophagy and UPS are considered two distinct and interacting proteolytic systems, and p62 is considered an integration center for autophagy and UPS (Lee et al, 2020; Xiong et al, 2018). In selective autophagy, cytoplasmic components are selected and tagged before being transported into an autophagosome by means of selective autophagy receptors such as p62. Selected components are tagged, often with ubiquitin, to facilitate recognition by autophagy receptors. On the other hand, the expression of p62 is also markedly increased during conditions in which autophagy substrates accumulate. Therefore, p62 is considered an important component in delivery and clearance of ubiquitinated proteins (Mattern et al, 2019; Wang et al, 2019). Given the networked relationship between p62, autophagy, and UPS, we next assessed the levels of p62 and HBcAg in LV-UbHBcAg groups. MG132 and CQ are inhibitors of the UPS and autophagy, respectively.
In this study, the immunofluorescence images showed that the presence of CQ or/and MG132 resulted in an accumulation of p62-positive punctate structures in DCs. Notably, we observed that both compounds increased the protein levels of p62 and HBcAg, suggesting that the degradation of p62 and HBcAg reduced, and change of HBcAg is probably associated with p62 levels. These results suggested that p62 may play an important role in UbHBcAg degradation; in addition, both the UPS and autophagy pathways have an important effect on the degradation of UbHBcAg. However, its underlying molecular mechanism needs to be further explored.
Previous studies have indicated that there exists a strong and complex relationship between autophagy and apoptosis under different conditions (Zheng et al, 2019). Autophagic and apoptotic modes of cell death can coexist in response to exoteric stimuli or may be cause-and-effect incidents (Feng et al, 2018; Goodall et al, 2016). Therefore, we examined the level of apoptosis in DCs treated with LV-UbHBcAg. Western blot analysis showed that the levels of apoptotic proteins were increased compared with those in the NC group. A similar pattern was also observed for the percentage of apoptotic DCs; the apoptotic cells in the LV-UbHBcAg groups were higher than those in the NC group. With 3-MA treatment, the number of apoptotic cells in the LV-UbHBcAg group was significantly reduced. The overall results indicate that the promotion of autophagy was associated with apoptosis in DCs loaded with LV-UbHBcAg.
Finally, we would like to mention that one of the most important proteins, mTOR (mTORC1), as a central regulator of cell growth, plays a key role in coordinating the balance between cell growth and catabolism in response to nutritional status and various stress signals (Sethna et al, 2021). An increasing number of studies have reported that mTOR can negatively regulate autophagy activity, and the PI3K/Akt/mTOR signaling pathway plays an important role in regulating cell autophagy and survival (Deng et al, 2021; Zheng et al, 2019). In this study, we showed that LV-UbHBcAg treatment resulted in a reduction in PI3K/Akt/mTOR signaling pathway-related proteins, which are relevant to activation of the autophagy process. In addition, rapamycin treatment significantly downregulated the level of phosphorylated mTOR in DCs. Thus, these results dedicate that induction of the autophagic process in DCs is strictly related to mTOR.
Conclusions
Taken together, the recovery of DC function suppressed by HBV infection can be an important part of the development of immunological therapies aiming to achieve a functional cure for chronic HBV infection. Our results showed an important role of LV-UbHBcAg in DC activation and suggested that LV-UbHBcAg could stimulate DC immune function and enhance autophagy process, which may potentially be used as a therapeutic strategy to combat persistent HBV infection. Notably, the precise mechanism underlying the interplay between autophagy and UPS in ubiquitinated protein degradation is not yet fully understood, and this issue requires further investigation. In addition, our study reports the beneficial effects of LV-UbHBcAg at the cellular level, but there is still a lack of evidence at the animal level. Therefore, in future research, we will focus on the immunomodulatory effects of LV-UbHBcAg on DCs in vivo.
Footnotes
Authors' Contributions
R.H. drafted and finalized the article. Z.T. was responsible for revising and critical reading. We thank J.C., W.H., X.C., and Q.T. for valuable discussions. We are grateful to Y.Y. and G.Z. for their assistance.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by grants from the National Natural Science Foundation of China (Grant Nos. 81702050 and 82100631).