
Abstract
Antimicrobial peptides are proteins that have been found to be an important factor in the natural immune response to a variety of pathogens. Respiratory syncytial virus (RSV) is a respiratory pathogen with the capability to cause serious upper and lower respiratory infections in infants and children and is a major viral cause of infant mortality. There is currently no functional vaccine for the virus, as recent efforts have been hindered by the virus's low immunogenicity, its ability to effectively mutate, and underlying instabilities of potential vaccines. Previous studies have shown that antimicrobial peptides may affect viral replication and spread of RSV. Our study evaluates the susceptibility of chimeric strains of RSV that express different fusion (F) and attachment (G) proteins to susceptibilities to inactivation by LL-37 and human beta-defensins (hBDs) hBD-1, hBD-3, and hBD-4. We show that LL-37 and hBD-3 result in dose-dependent, strain-independent inactivation of RSV, whereas treatment with either hBD-1 or hBD-4 appears more variable between strains. This suggests a potential role of the viral structural proteins in mitigating the inhibitory effects of the peptides. This study provides the first evidence of the sensitivity of RSV to several hBDs and indicates a role of LL-37 and beta-defensins in both limiting establishment of natural RSV infections and in the therapeutic treatment of severe RSV disease.
Introduction
Antimicrobial peptides (AMPs) are small peptides secreted by a wide range of organisms, including animal cells, plants, fungi, and bacteria, as well as viruses (Jenssen et al, 2006; Yeaman and Yount, 2003). They were first discovered during a 1939 experiment where secreted molecules from Gram-positive bacteria were shown to be capable of destroying living cells of other Gram-positive species, as well as limiting the growth and spread of these species in culture media (Dubos, 1939). Since their initial discovery, they have been found to play an important role in the immune response of organisms to defend against infections through direct antimicrobial activity (Anonymous, 2013). The structures of AMPs can be categorized as a loop, extended, α-helical, or β-sheet peptides and as a whole are categorized as amphiphilic (Hancock and Sahl, 2006; Seo et al, 2012; Zasloff, 2002).
Recently, AMPs have been used as a therapeutic option to replace common antibiotics, such as penicillin, because they have a low susceptibility to acquisition of resistance (Zasloff, 2002). AMPs have previously been shown to inhibit the replication of HIV-1 through gene expression suppression as well as displaying antiproliferative and antitumor activity against four different bladder cancer cell lines (Suttmann et al, 2008; Wachinger et al, 1998). However, it remains unclear what role many known mammalian AMPs play in regulating respiratory viruses.
Human respiratory syncytial virus (hRSV) is an enveloped, negative-sense RNA (-ssRNA) virus classified into the Pneumovirus family (Borchers et al, 2013; Bennett et al, 2014). RSV is responsible for both upper and lower respiratory tract infections with young infants and the elderly being most at risk for severe disease (Hall et al, 2001; Walsh and Falsey, 2012). RSV is the most abundant cause of sudden-onset respiratory tract infections, especially in young children and is predominantly found in infants that are <6 months of age (Cattoir et al, 2019; Hall et al, 2001). During infection, RSV utilizes two surface proteins, G and F, for attachment and entry into cells. RSV glycoprotein G mediates the initial attachment of the virus to cells, whereas the RSV fusion peptide (F) is responsible for fusion of the viral envelope to cell membranes. Due to their roles in the viral attachment and entry processes, both RSV F and G proteins are responsible for eliciting the majority of neutralizing antibodies during active infections (Collins and Melero, 2011; Levine et al, 1987). In addition, we have previously shown that RSV F and G are the primary viral proteins that regulate virion stability, making them attractive targets for vaccine and therapeutic design.
Human cathelicidin or LL-37 is an alpha-helical, amphipathic AMP secreted predominately by epithelial cells. It has been previously shown to inactivate RSV both in vitro and in vivo using a variety of culture models (Currie et al, 2013; Gwyer Findlay et al, 2013; Sørensen et al, 1997). In addition to LL-37, other human AMPs have been identified, which inactivate RSV.
Human beta-defensins (hBDs) are small, cysteine-rich AMPs that are known to be active against a wide array of pathogens. Due to the presence of numerous disulfide bonds, hBDs are morphologically distinct and structurally stable molecules that have been shown to activate host innate immune cells (Brice and Diamond, 2019). While human beta-defensin 2 (hBD-2) has been shown to reduce RSV infection, it remains unclear what impacts hBDs 1, 3, and 4 have on the regulation of RSV despite being secreted into the respiratory epithelium (Pazgier et al, 2006). It has been hypothesized that LL-37 and human respiratory hBDs inactivate RSV through direct damage to the viral envelope as well as potentially triggering other antiviral activities (Bahar and Ren, 2013). However, all studies to date have only used the common laboratory strain of RSV, A2, in their inactivation assays (Currie et al, 2013, 2016; Harcourt et al, 2016).
In this study, we will use a panel of three recombinant RSV strains that differ only in the F and G protein expression to evaluate what roles, if any, these surface proteins which govern virus stability have on their susceptibility to inactivation by human AMPs. In this experiment, we analyze the susceptibility of multiple RSV strains to LL-37 and demonstrate inactivation of RSV by hBD-1, hBD-3, and hBD-4. This study highlights an important feature of the host innate immune events during RSV infection and may represent a novel direction in development of therapeutics to treat RSV.
Materials and Methods
Cells, viruses, and AMPs
Panel strains used in this study of recombinant RSV viruses expressing monomeric Katushka 2 (mKate2), a far-red fluorescent reporter, were constructed and recovered previously using an RSV A2-based reverse genetics system (Stobart et al, 2016). The strains used in this study were A2, A2-line19F, and A2-12-21GF (Fig. 1). The RSV strains were propagated in Hep2 cells (ATCC CCL-23) in minimal essential medium (MEM) supplemented with 10% fetal bovine serum with a penicillin, streptomycin, and amphotericin B antibiotic mixture (Corning). The AMPs used in this study include LL-37 (AnaSpec, Inc.), hBD-1 (AnaSpec, Inc.), hBD-3 (AnaSpec, Inc.), and hBD-4 (PeproTech, Inc.). These peptides were prepared according to the manufacturer's recommendations for resuspension in stock solutions, which were further diluted for use in neutralization and replication curve analyses.
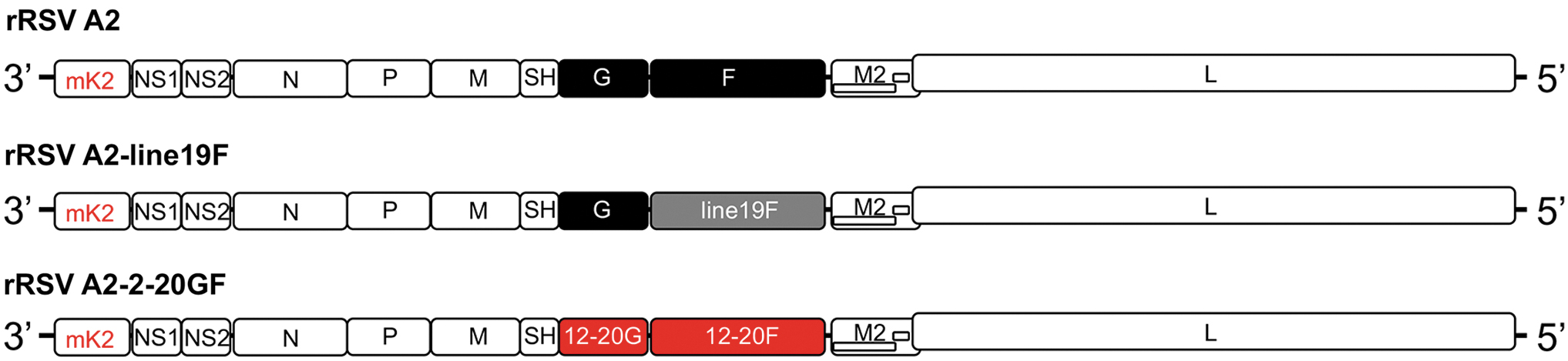
Recombinant RSV strains evaluated for sensitivity to AMPs. Recombinant strains A2, A2-line19F, and A2-2-20GF were all synthesized using an RSV A2-based reverse genetics system previously described (Hotard et al, 2012). Strain A2-line19F differs from A2 in the expression of a heterologous fusion (F) peptide (in gray) from strain Line19. Strain A2-12-20GF differs from A2 in the expression of heterologous glycoprotein (G) and fusion (F) peptides (in red) from strain 12–20. All three recombinant viruses also express a red fluorescent mK2 reporter protein. AMPs, antimicrobial peptides; mK2, monomeric Katushka 2; RSV, respiratory syncytial virus.
The experiments and protocols described herein were given approval by the Butler University Institution Biosafety Committee.
Neutralization and titering assays
A neutralization assay for antimicrobial activity was performed by mixing serial dilutions of AMP preparations of LL-37 (cathelicidin), hBD-1, hBD-3, or hBD-4 with diluted RSV strains, A2, A2-line19F, and A2-12-21GF, for 1 h at 37°C before infection of confluent monolayers of HEp2 cells in 96-well plates. Virus titers were determined by quantifying virus-infected red foci (due to expression of the mKate2 reporter) visualized using a Texas Red filter. The percent inhibition of no treatment was calculated for each AMP dosage by dividing the number of detected foci in the treatment well by the number of detected foci in the untreated well. Determination of the inhibitory concentration for a 50% reduction (IC50) for each treatment was calculated by using a line of best fit to the average inhibition data collected to identify the concentration, which would correlate with 50% reduction in detected virus titer.
Bacterial inactivation experiments
Tryptic soy broth cultures were inoculated with equal volumes of Escherichia coli and either AMP (experimental condition), solvent (positive control for growth), or a 0.02% sodium azide solution (negative control for growth). At time points, the optical density (OD) of the solution was measured at 600 nm.
Replication curve analysis
Confluent monolayers of HEp2 cells were infected at a multiplicity of infection of 0.01 in complete MEM containing 25 μg/μL of AMP treatment or a mock treatment control. At times of 0, 1, 3, and 5 days postinfection (d.p.i.), the virus titer was determined through counting of mKate2 fluorescence as described above.
Results
Prior studies have shown that the AMP, LL-37, was capable of inhibiting replication of RSV A2 in both HEp2 and in air–liquid interface cultures as well as in vivo (Harcourt et al, 2016). LL-37 consists of a helical molecule with mostly hydrophobic and charged/hydrophilic faces, which is believed to disrupt viral envelope (membrane) integrity (Fig. 2A). However, due in part to known differences in stability between RSV strains mediated by the RSV F and G structural proteins, it remains unclear whether there are RSV strain-specific differences in susceptibility to LL-37-mediated inactivation. To address this question, an initial stock solution of LL-37 was prepared and evaluated for inhibition against a culture of E. coli (Fig. 2B), which has previously been shown to be sensitive to LL-37 in low concentrations (Mount et al, 2007). After 4 h of incubation, E. coli treated with LL-37 showed a significant reduction (p = 0.026; t-test) in OD compared with an untreated culture control.

Inactivation and inhibition of Escherichia coli and recombinant RSV strains by human LL-37 (cathelicidin).
To test for sensitivity of RSV to LL-37-mediated inactivation, three different RSV strains were mixed with serial dilutions of LL-37: A2, A2-line19F, or A2-2-20GF (Fig. 2C). All three strains were generated using an RSV A2 reverse genetics platform but differ in the expression of heterologous F and/or G proteins. While subtle differences were observed between strains, all three viruses exhibited a dose-dependent level of inhibition with no significant differences observed between strains. A Dunnett's test of the varying experimental doses versus the control revealed statistical significance in reduction of virus at the 25 and 12.5 μg/mL (p < 0.05). The LL-37 inhibitory concentrations associated with 50% reductions in infectious virus (IC50) for RSV strains, A2, A2-line19F, and A2-2-20GF, were 14.5, 15.8, and 15.1 μg/mL, respectively, with the average determined to be 15.1 ± 0.7 μg/mL.
It has been previously reported that the primary mechanism for LL-37 to inactivate RSV is through disruption of the viral envelope integrity (Ahmed et al, 2019). It has also been hypothesized that LL-37, may also bind to the viral fusion F protein, as part of its inactivation process. To determine what role, if any, differences in attachment and fusion proteins may have on susceptibility to inactivation, RSV replication was tracked over 5 days in the presence or absence of an inhibitory dosage of 25 μg/mL of LL-37 (Fig. 3). For all three strains, there was a significant reduction in replication titers between the treatment and control conditions (analysis of variance [ANOVA], p = 0.012). No significant differences were observed in the reductions between strains. LL-37 was more effective at reducing viral titers earlier during infection rather than late in infection with an average 23-fold reduction of detected titers at 1 d.p.i. compared with sevenfold and fourfold reductions at days 3 and 5 p.i., respectively.
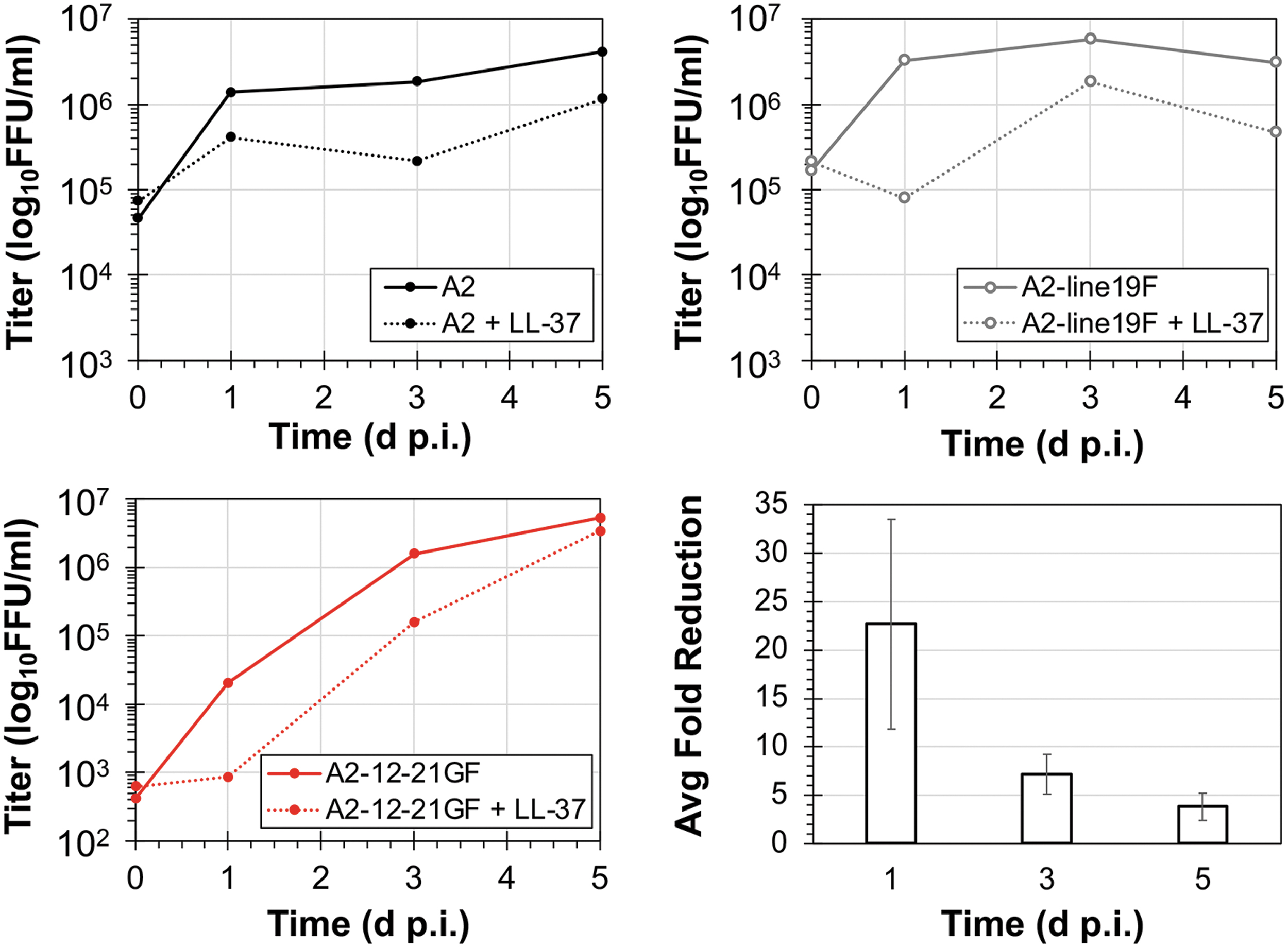
Inhibition of RSV replication by human LL-37 (cathelicidin). HEp2 cells were infected with an MOI of 0.01 in complete MEM cell culture media containing either 25 μg/mL of human LL-37 (dashed lines) or mock treatment (solid lines). Cells and supernatant were harvested at 0, 1, 3, and 5 d.p.i. and subsequently quantified by mKate2 expression on HEp2 cells. The average titer (n = 2) is reported as FFU/mL. The average fold reduction compared with mock treatment for the three RSV strains at 1, 3, and 5 d.p.i. is shown at the bottom right. d.p.i., days postinfection; FFU/mL, fluorescent focus units per mL; MEM, minimal essential medium; MOI, multiplicity of infection.
While hBD-2 has been shown to inhibit RSV, it remains unknown whether other identified beta-defensins in humans are capable of inactivating RSV. hBDs 1, 3, and 4 are structurally distinct from LL-37, however, they share a mixture of mostly hydrophobic and positively charged regions on their surfaces (Fig. 4A). Unlike hBD-1, and hBD-3, however, hBD-4 exhibits significantly more negatively charged, acidic residues on its peptide surface. When mixed with serial dilutions of the beta-defensins, all three viruses exhibit a dose-dependent reduction in virus. The sensitivity of the viruses to inactivation differs between peptide treatment. hBD-1 treatment resulted in an average IC50 of 75.5 ± 30.8 μg/mL, with markedly higher variability between strains. In contrast, hBD-3 treatment showed a similar IC50 (14.0 ± 0.9 μg/mL) and higher level of consistency between strains to that of LL-37. Lastly, treatment with hBD-4 resulted in an IC50 of 29.5 ± 9.3 μg/mL, with a greater level of variability between strains than LL-37 and hBD-3 treatment.

Inhibition of RSV replication by hBDs.
Discussion
Human AMPs play critical roles in the innate immune defenses against a wide array of pathogens, including fungi, bacteria, and viruses (Bahar and Ren, 2013; Hsieh and Hartshorn, 2016; Jenssen et al, 2006). In this study, we evaluated (1) whether LL-37 and a panel of hBDs could inhibit RSV and (2) if any observed inhibitions were dependent upon the viral structural proteins F and G. Past studies evaluating the susceptibility of RSV to inactivation by AMPs have been limited to a single strain, A2. We found that all four peptides evaluated in this study (LL-37, hBD-1, hBD-3, and hBD-4) are capable of inhibiting RSV in a dose-dependent manner with variable levels of sensitivity and consistency between RSV strains.
Consistent with prior studies, LL-37 was effective at lower concentrations (Table 1) and showed a high level of consistency between RSV strains that contain heterologous F and/or G structural proteins. Furthermore, we show that greatest impact of LL-37 on viral replication occurs earlier rather than later during infection in vitro. We did not test postinfection application of LL-37 against RSV. Previous work with LL-37 and RSV strain A2 demonstrated significantly less potency when administered as little as 2 h postinfection suggesting that either pretreatment or concurrent treatment with infection is necessary for optimal inactivation (Currie et al, 2013). Studies evaluating the in vivo concentrations of LL-37 have shown that neutrophil granules can contain concentrations as high as 40 μM and other studies have reported physiologic concentrations as high as 15 μM suggesting a potential role of in vivo expression of LL-37, and potentially other AMPs, in regulating the persistence of RSV infection.
IC50 for Inactivation of Recombinant Respiratory Syncytial Virus Strains with LL-37 and Human Beta-Defensins
hBD-3, human beta-defensin 3; RSV, respiratory syncytial virus; SD, standard deviation.
In addition, while the potency of these peptides toward RSV is mostly in the low micromolar range, chemical modifications to these molecules and long-term application may provide additional avenues for therapeutic treatment (Schaller-Bals et al, 2002). These data support the hypothesis that this AMP is targeting the viral membrane envelope structure, which would be most sensitive before attachment and entry of the virus and suggests that structural gene expression likely plays a limited role in the inactivation function of LL-37. Similar to LL-37, we show for the first time that treatment with hBD-3 results in both dose-dependent and strain-independent inactivation of RSV. The IC50 and variability for LL-37 (15.1 ± 0.7 μg/mL) and hBD-3 (14.0 ± 0.9 μg/mL) were very similar despite distinct tertiary structures. Other groups have suggested that, like LL-37, hBDs target membrane integrity. The consistency between strains observed in the data for hBD-3 would be consistent with this hypothesis. However, treatment with either hBD-1 or hBD-4 resulted in far greater variability between strains.
We previously reported that strain A2-line19F exhibits significantly greater thermal stability than strain A2. When treated with either hBD-1 or hBD-4, the IC50 for A2-line19F appears nearly twice as high as that of strain A2. These data collectively suggest that mechanisms of inactivation beyond disruption of membrane integrity may occur and should require additional study.
To date, there remains no commercially available vaccine for RSV and only a limited and costly prophylaxis therapeutic option (palivizumab), despite a significant clinical burden of disease (Shi et al, 2017). Several studies have demonstrated roles for both LL-37 and hBDs in both inactivation and immunoregulation against a wide array of viruses, including Influenza, Dengue, Zika, and SARS-CoV-2 (Ahmed et al, 2019; Elnagdy and AlKhazindar, 2020; Hsieh and Hartshorn, 2016). Studies in HIV have shown evidence for differences in pathogenicity of HIV associated with distinct human polymorphisms in AMP genes (Baroncelli et al, 2008; Braida et al, 2004; Milanese et al, 2006). These studies may support a role of AMP polymorphisms in susceptibility to RSV infection and the potential for advanced disease. Future retrospective studies may aim to evaluate whether correlations between specific polymorphisms in AMPs can predict RSV infection health outcomes.
This study provides the first evidence of the broad neutralizing capacity of RSV with hBDs and demonstrates that LL-37 is capable of inactivating RSV in a strain-independent manner. The sensitivity of RSV to inactivation by both LL-37 and hBDs provides evidence for a potential therapeutic application for either administration of purified AMPs or treatments, which promote increased expression of these peptides in the respiratory mucosa. In addition, these data may suggest that differences in natural expression of these peptides in respiratory mucosa may affect susceptibilities to RSV infection in high-risk populations. Lastly, these data support further studies into the impacts of LL-37 and hBD secretion on RSV susceptibility and infection.
Footnotes
Acknowledgments
The authors would like to thank Sean Berthrong and other members of the Stobart laboratory for their support and meaningful discussions regarding both the direction of these studies and the writing of this article.
Authors' Contributions
K.N.L., A.T.J., N.A.J., C.E.H., K.R.C., S.R.K., and E.R.H. were all directly involved in experiment planning, data collection, analysis, and writing. M.P.T. and C.C.S. were involved in experiment planning, data analysis, and writing.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by an Indiana Academy of Science (IAS) Senior Research Grant, funding from the Butler University Holcomb Awards Committee (HAC), the Butler University Department of Biological Sciences, and the Butler Summer Institute (BSI).