
Abstract
The dysfunction of memory CD8+ T cell cannot be reverted by successful clearance of hepatitis C virus (HCV) after direct-acting antivirals (DAAs) therapy, increasing the risk of reinfection with HCV. Stem cell-like memory T cells (Tscm) with superior properties of long-lasting, self-renewing, and multipotency contribute to the maintenance of immune function. We investigated the impact of HCV infection on CD8+ Tscm, and their possible role in disease progression, by using DAA-naive HCV-infected and human immunodeficiency virus (HIV)/HCV-coinfected cohorts. The distribution of memory CD8+ T cell subsets and the level of T cell immune activation were determined by flow cytometry. Associations between CD8+ Tscm and other memory T cell subsets, HCV viral load, as well as the level of T cell immune activation were analyzed. We observed that the proportion of CD8+ Tscm increased in both HCV and HIV/HCV individuals. The proportion of CD8+ Tscm had positive and negative correlation with CD8+ Tcm (central memory T cells) and CD8+ Tem (effector memory T cell), respectively, representing the contribution of CD8+ Tscm in T cell homeostasis. In addition, higher frequency of CD8+ Tscm indicated lower HCV viral load and less T cell immune activation in HCV infection, which suggested that CD8+ Tscm is likely associated with effective control of HCV replication for protective immunity. Considering the characteristics of Tscm, our current findings provide implications for Tscm-based vaccine design and immunotherapy development to achieve HCV elimination.
Introduction
Approximately 58
Despite the high HCV clearance rates in most individuals (∼98%) achieved by interferon (IFN)-free direct-acting antivirals (DAAs) therapy, other negative effects including DAA resistance and partial restoration of adaptive immune functions after DAA-mediated HCV cure (especially the incomplete recovery of memory CD8+ T cell responses) can develop. These, in turn, affect the treatment outcomes and increase the risks for HCV reinfection and opportunistic infection after cure in HCV-infected and HIV/HCV-coinfected individuals (Barili et al, 2021; Hofmann et al, 2021). Therefore, continuing efforts and understanding are required to develop novel strategies to achieve HCV eradiation.
Adoptive cell therapies have shown promising efficacy against viral infections, leukemias, and solid tumors (Mao et al, 2022; Shi et al, 2022; Vishwasrao et al, 2022; Xu et al, 2019). Selection of transferred T cells with the characteristics of longevity and strong cytotoxicity is of high importance for clinical applications. Also, efficient and persistent memory CD8+ T cell responses are essential for long-term defense against viral infection or malignancies (Kaech et al, 2002; Restifo et al, 2012). The memory T cell compartment can be divided into three subsets according to a developmental hierarchy: stem cell-like memory T cells (Tscm), central memory T cells (Tcm), and effector memory T cells (Tem) (Gattinoni and Restifo, 2013; Gattinoni et al, 2017; Lugli et al, 2020).
Compared with Tcm and Tem, Tscm has superior capacity of long-lasting, quick emergence, self-renewing, and multipotency for generation of other memory T cells (Gattinoni et al, 2011; Lugli et al, 2013; Wang et al, 2021). Our previous study has demonstrated that Tscm is responsible for immune reconstitution in HIV-1 chronically infected patients (Lu et al, 2017). The CD8+ Tscm subset has more beneficial effects of antiviral or antitumor responses in the clinic, as well as the immune reconstitution after hematopoietic stem cell transplantation (Cieri et al, 2015; Ribeiro et al, 2014; Sabatino et al, 2016).
Of note, functional HCV-specific CD4+ and CD8+ Tscm can be elicited robustly by an adenoviral/modified vaccinia Ankara vector-based vaccination (Hartnell et al, 2020; Swadling et al, 2014). In addition, yellow fever-specific CD8+ Tscm cells can be stably maintained least >25 years postvaccination (Fuertes Marraco et al, 2015), whereas SARS-CoV-2-specific CD8+ Tscm in COVID-19 convalescent patients is sustained for 10 months (Jung et al, 2021). Therefore, CD8+ Tscm became a novel target for vaccine design against viral infection and the development of T cell-based immunotherapy for antiviral and antitumor therapy or immune reconstitution post-transplantation.
Understanding the changes of CD8+ Tscm compartment in HCV-infected and HIV/HCV-coinfected subjects and its correlation with virological and immunological parameters in disease may offer insights into further application of this subset in vaccine or immunotherapy. In this study, we demonstrated that the proportion of CD8+ Tscm increased in both HCV infection and HIV/HCV coinfection. Furthermore, the proportion of CD8+ Tscm is inversely correlated with HCV viral load or the level of T cell immune activation in HCV infection, indicating the important role of CD8+ Tscm in the impeding disease progression in HCV infection.
Materials and Methods
Study cohorts
Thirty-six healthy controls (HCs), 25 antiretroviral therapy (ART)-treated HIV-1-infected patients (HIV+), 28 untreated HCV chronically infected patients (HCV+), and 18 HIV virologically suppressed but HCV-untreated HIV/HCV-coinfected patients (HIV/HCV) were enrolled from four cohorts established in Beijing Youan Hospital. (1) HCs were anti-HIV, antihepatitis B virus (HBV), and anti-HCV antibody negative individuals, who were recruited from the men who have sex with men high-risk behavior group of HIV-1 infection screening cohort; (2) HIV+ patients with negative anti-HBV and anti-HCV antibodies were recruited from chronically HIV-infected ART-treatment cohort. The median HIV-1 infection duration was 4 years, and patients were treated for at least 2 years where plasma HIV-1 RNA was undetectable. HIV+ patients selected in this study were 96 weeks with ART. The ART regimen included AZT (zidovudine)/3TC (lamivudine)/NVP (nevirapine) (n = 12) or TDF (tenofovir)/3TC/EFV (efavirenz) (n = 13); (3) HCV+ patients with negative anti-HIV and anti-HBV antibodies were recruited from HCV-infected cohort. The median HCV infection duration was 3 years. HCV+ patients selected in this study were treatment-naive timepoint; (4) HIV/HCV patients with negative anti-HBV antibody were recruited from the HIV/HCV-coinfected cohort. Patients in this cohort had been diagnosed with HIV-1 before HCV infection. The median HIV-1 infection and treatment duration were 5 and 3 years, respectively. The ART regimen against HIV-1 included AZT/3TC/NVP (n = 8) or D4T (stavudine)/3TC/EFV (n = 10). The median HCV infection duration was 4 years. Patients selected from this group for this study were plasma HIV-1 RNA undetectable and HCV untreated. All subjects enrolled in this study had CD4+T cell count >200 cells/μL and detailed information is listed in Table 1.
Clinical Data for Study Subjects
—, data not available.
ALT, alanine aminotransferase; AST, aspartate aminotransferase; HCs, healthy controls; HCV, hepatitis C virus; HIV-1, human immunodeficiency virus type 1.
Peripheral blood mononuclear cells (PBMCs) from whole blood were isolated using a Ficoll–Hypaque gradient centrifugation method and stored in liquid nitrogen.
Ethics statement
This study and all the relevant experiments were approved by the Beijing Youan Hospital Research Ethics Committee (Approval No.: 2019-014), and written informed consent was obtained from each participant in accordance with the Declaration of Helsinki.
Immunophenotyping
Cryopreserved PBMCs were thawed in RPMI 1640 medium (Invitrogen, Carlsbad, CA, USA) and were washed with phosphate-buffered saline containing 1% bovine serum albumin. The cells were surface stained with live/dead fixable viability 510 (BD Biosciences, San Diego, CA, USA) for 15 min to exclude dead cells. Cells were then washed and surface stained with the following mouse antihuman monoclone antibodies at room temperature for 20 min in the dark: CD3-APC-Cy7 (Clone SK7), CD4- FITC (Clone SK3), CD8-Percp-Cy5.5 (Clone RPA-T8), CD45RA-PE-Cy7 (Clone HI100), CCR7-APC (Clone G043H7), CD27-PE (Clone O323), CD95-BV421 (Clone DX2), CD38-PE (Clone HIT2), and HLA-DR-APC (Clone TU36).
Relative isotype controls were also used. All antibodies and isotype controls were purchased from Biolegend (San Diego, CA, USA). At least 50,000 lymphocytes were acquired with BD FACS Canto II flow cytometer (BD Biosciences). Data were analyzed with Flowjo Software version 10 (Tree Star, Inc., Ashland, OR, USA). Tscm were defined as CD45RA+ CCR7+ CD27+ CD95+, Tcm were defined as CD45RA− CCR7+, and Tem were defined as CD45RA− CCR7− accordingly (Ribeiro et al, 2014).
Measurements of number of CD4+ T cells and plasma viral load
Routine CD4+ T TruCount (BD Biosciences), plasma HIV-1 viral load (Abbott Molecular, Inc., Des Plaines, IL, USA), and plasma HCV viral load (Roche Diagnostics, Mannheim, Germany) were measured according to the manufacturers' instructions. The detecting limitation of HIV-1 and HCV is 50 copies/mL and 15 IU/mL, respectively.
Statistical analysis
Data in figures are expressed as means ± standard deviation, or presented as median with range in Table 1. All statistical analyses were performed using GraphPad Prism software v5.0 (GraphPad Software, Inc., La Jolla, CA, USA). Comparisons between two groups were done using a Student's t test for data with normal distribution, and a nonparametric Mann–Whitney U-test was used with data that were not normally distributed. Correlations were determined by the Spearman rank correlation test. A p-value <0.05 represented significance.
Results
Elevated CD8+ Tscm proportion in HCV-infected and HIV/HCV-coinfected patients
To determine the role of CD8+ Tscm during HCV infection or HIV/HCV coinfection, we first analyzed the proportion of this compartment in untreated HCV+ and HIV virologically suppressed with viral load of <50 copies/mL but HCV-untreated HIV/HCV coinfected individuals. HCs and ART-experienced HIV+ patients with viral load of <50 copies/mL were used as controls. The gating strategy used for CD8+ Tscm analysis is shown in Figure 1A.
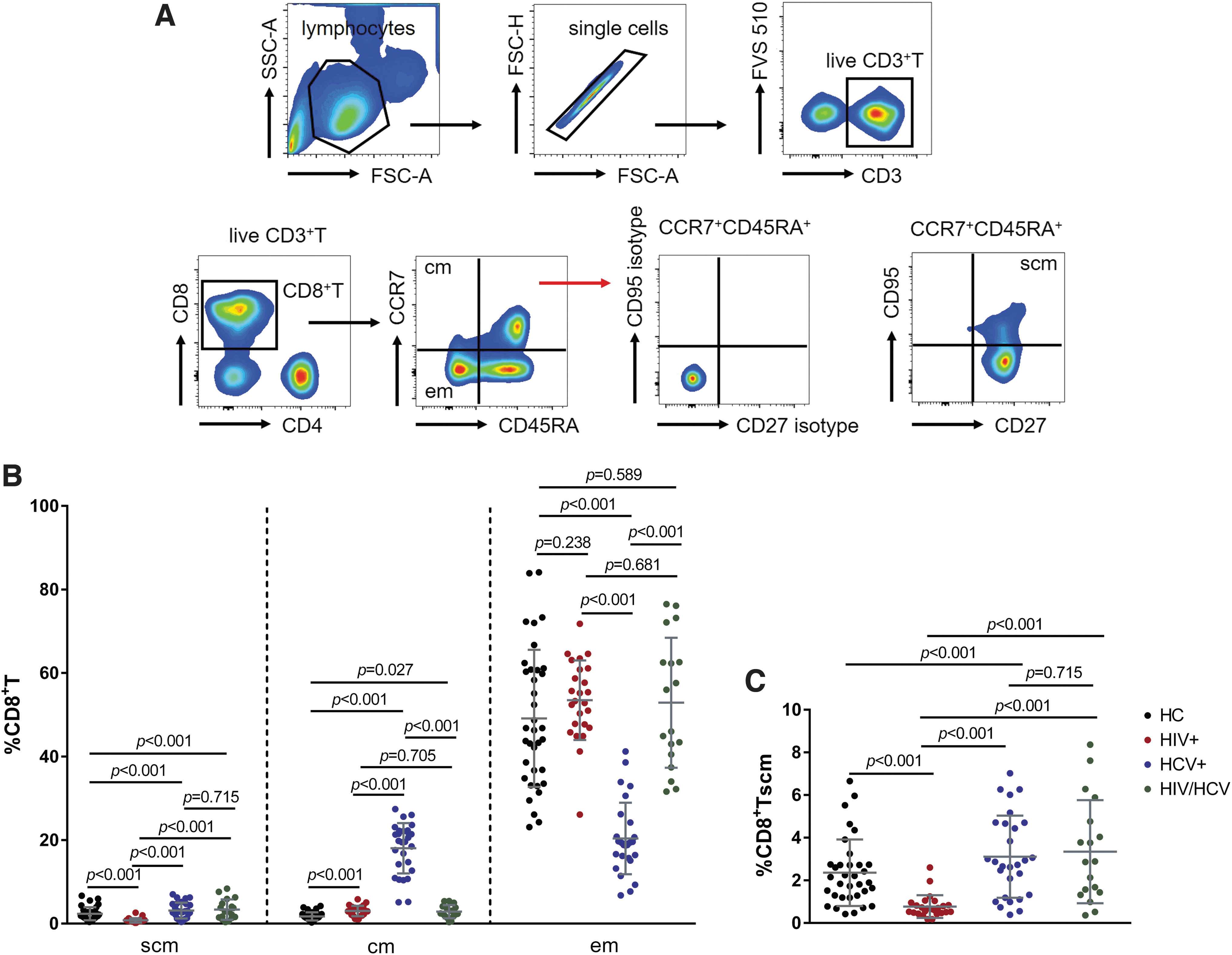
Distribution of memory CD8+T cells in HCs, HIV+, HCV+, and HIV/HCV.
Compared with HCs, the proportion of CD8+ Tscm in both HCV+ and HIV/HCV significantly increased (all p < 0.001), however, there was no difference between HCV+ and HIV/HCV (p = 0.715) (Fig. 1B, C). HIV+ had the lowest CD8+ Tscm frequency among four groups (all p < 0.001), whereas HCV+ had the highest CD8+ Tcm and the lowest CD8+ Tem frequencies among four groups (Fig. 1B).
CD8+ Tscm is involved in memory T cell maintenance during HCV infection
The frequency of CD8+ Tscm had a positive correlation with that of CD8+ Tcm in HIV+ (r = 0.640, p < 0.001) and HCV+ (r = 0.425, p = 0.024), and a similar correlation was found with HIV/HCV (r = 0.445, p = 0.086) (Fig. 2A). Conversely, negative correlation between the frequency of CD8+ Tscm and that of CD8+ Tem was observed in HCs (r = −0.630, p < 0.001), HIV+ (r = −0.538, p = 0.005), and HCV+ (r = −0.447, p = 0.016), and a similar trend in HIV/HCV (r = −0.418, p = 0.084) (Fig. 2B). These findings indicated that during the initial stage of memory T cell differentiation, Tscm may support the development of Tcm, which may subsequently differentiate into Tem in HCV infection.
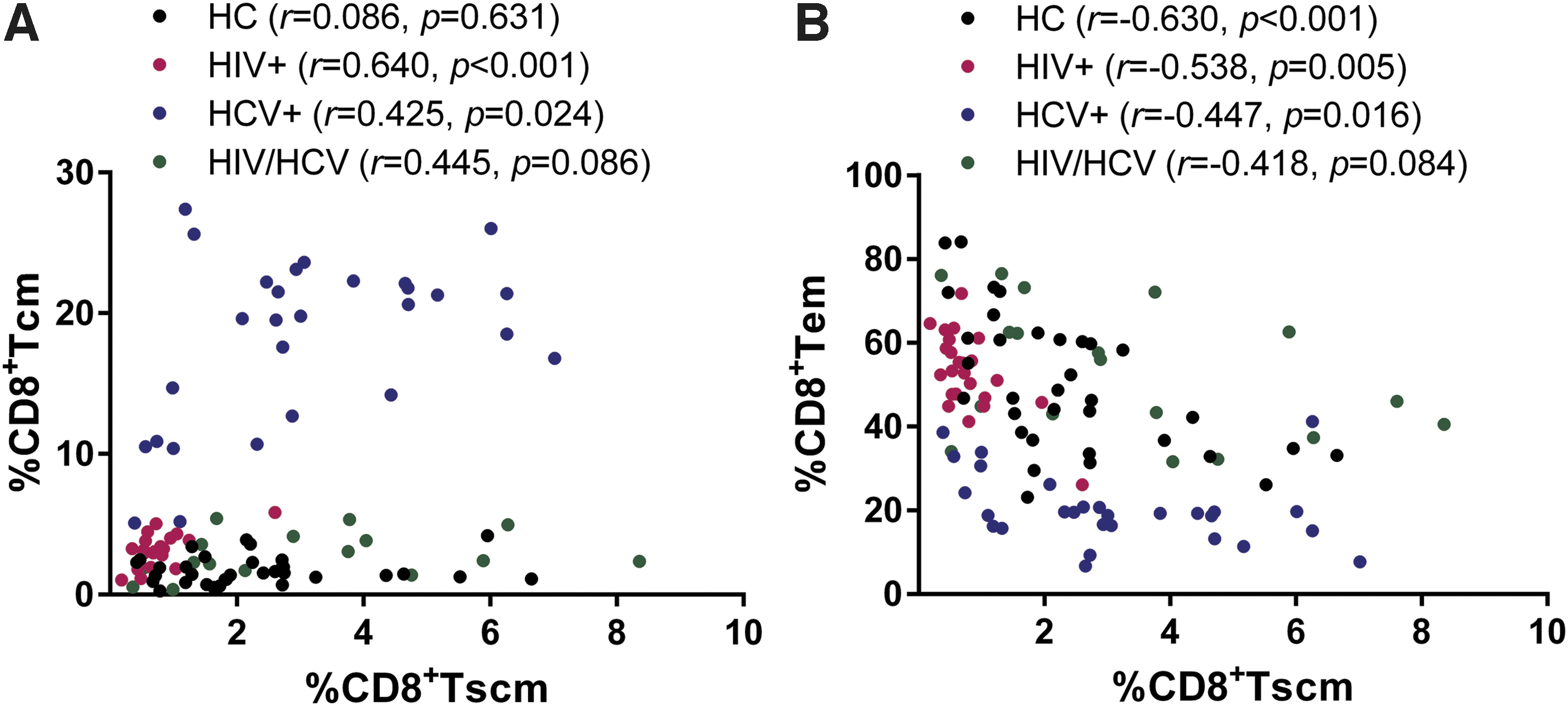
CD8+ Tscm frequency correlates with other memory CD8+ T cells. Correlation analysis between the frequency of CD8+ Tscm and that of CD8+ Tcm
Association between CD8+ Tscm and HCV clinical parameter
Next, we investigated the possible influence of CD8+ Tscm on the virological progression in HCV+ and HIV/HCV-infected patients. There was no significant difference in plasma HCV viral load between HCV+ (median: 5.33 log10IU/mL) and HIV/HCV (median: 5.92 log10IU/mL) (Table 1). Inverse relationships were observed between plasma HCV viral load and the proportion of CD8+ Tscm, CD8+ Tcm, respectively, in HCV+ (CD8+ Tscm: r = −0.563, p = 0.002; CD8+ Tcm: r = −0.401, p = 0.034), but not in HIV/HCV (CD8+ Tscm: r = 0.133, p = 0.599; CD8+ Tcm: r = −0.256, p = 0.338) (Fig. 3A, B). In addition, the proportion of CD8+ Tem was not related to HCV viral load in both HCV+ (r = 0.163, p = 0.407) and HIV/HCV (r = −0.280, p = 0.261) (Fig. 3C).

CD8+ Tscm is correlated with low HCV viral load in HCV monoinfection. Correlation analysis between plasma HCV viral load and the frequencies of CD8+ Tscm
Since immune activation is an independent predictor for HIV-1 disease progression, as well as a key factor to drive liver injury and fibrosis in HCV-infected patients (Huang et al, 2022), we consequently analyzed the possible connection between CD8+ Tscm and the level of immune activation. By analyzing the frequency of CD38 and HLA-DR coexpression on global CD8+ T cell surface by flow cytometry in HCV+ and HIV/HCV patient samples, we found that HCV+ had the highest level of CD38+ HLA-DR+ CD8+ T cell among the four groups (all p ≤ 0.001) (Fig. 4A).
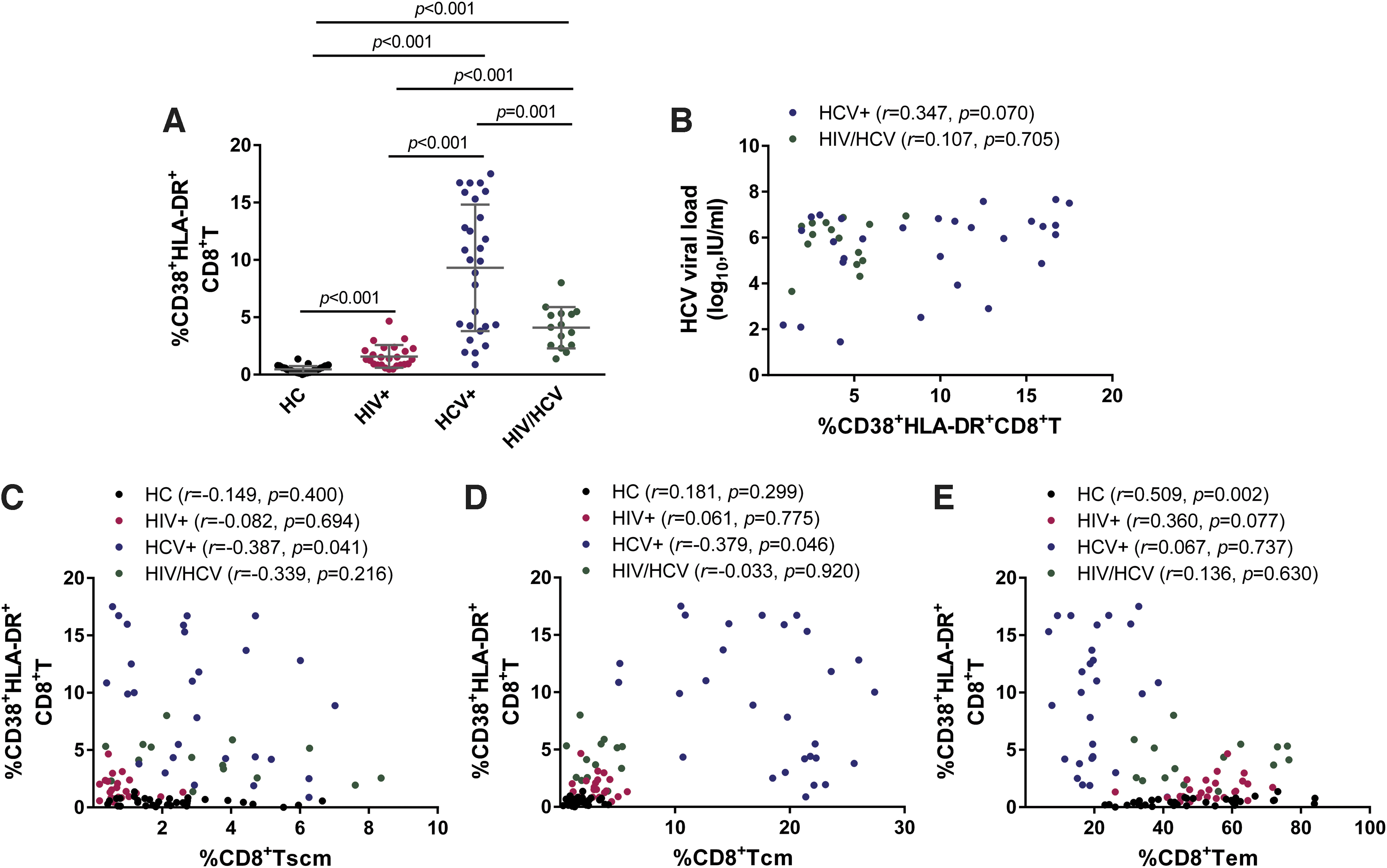
CD8+ Tscm is correlated with low level of T cell immune activation in HCV monoinfection.
Moreover, compared with HCs and HIV+, the frequency of CD38+ HLA-DR+ CD8+ T cell in HIV/HCV was increased (all p ≤ 0.001). Of note, the level of immune activation in HIV+ was higher than in HCs, although the virological suppression was achieved after successful ART in HIV+ individuals (p < 0.001). There was a trend of positive correlation between the frequency of CD38+ HLA-DR+ CD8+ T cell and plasma HCV viral load in HCV+ (r = 0.347, p = 0.070), but not in the HIV/HCV group (r = 0.107, p = 0.705) (Fig. 4B), indicating that HCV replication may drive the elevated level of immune activation in HCV-infected patients.
Correlation analysis revealed that within HCV+, the frequency of CD38+ HLA-DR+ CD8+ T cell was inversely related to that of CD8+ Tscm (r = −0.387, p = 0.041) (Fig. 4C), and CD8+ Tcm (r = −0.379, p = 0.046) (Fig. 4D). Similar relationship was not found when considering the same parameters in HCs, HIV+, and HIV/HCV. Besides, there was no association between the frequency of CD38+ HLA-DR+ CD8+ T cell and that of CD8+ Tem within all groups, with the exception of a positive correlation in HCs (r = 0.509, p = 0.002) (Fig. 4E).
We also analyzed the association between the memory CD8+ T cell compartments and the level of alanine aminotransferase (ALT) or aspartate aminotransferase (AST) in HCV+ and HIV/HCV groups. However, no significant correlation but a negative trend was observed between ALT or AST and the proportion of CD8+ Tscm, CD8+ Tcm, and CD8+ Tem in both HCV+ and HIV/HCV groups (data not shown).
Discussion
In this study, we observed the significant increase of the frequency of CD8+ Tscm in both HCV+ and HIV/HCV individuals. However, association between higher level of the CD8+ Tscm compartment and lower level of HCV RNA or that of T cell immune activation was only found in the HCV+ group.
The contribution of HCV-specific CD8+ T cells in the control of ongoing viral replication and consequent liver disease upon HCV infection has been extensively studied (Bowen and Walker, 2005; Hofmann et al, 2021; Shin et al, 2016). After successful HCV clearance, HCV-specific CD8+ T cells shift toward a memory phenotype with the ability to readily expand upon reinfection, making contribution to rapid and effective protection (Burchill et al, 2015; Osburn et al, 2010).
Persistent viral stimulation drives the dysfunction of memory CD8+ T cells through upregulation of coinhibitory molecules, metabolic derangements, proteostasis dysregulation, and chromatin silencing on CD8+ T cells (Barili et al, 2021). Although administration of DAAs results in a high rate of HCV cure and sustained virological responses, the memory CD8+ T cell dysfunction is irreversible in HCV and HIV/HCV patients, leading to the poor spontaneous control of HCV reinfection or other infections (Aregay et al, 2019; Bandera et al, 2021; Hensel et al, 2021; Perpinan et al, 2020).
CD8+ Tscm compartment has been identified as the precursor of other memory T cell subsets, supporting the long-time T cell memory in the postantigen phase and lifelong protection by yellow fever vaccination (Fuertes Marraco et al, 2015; Lugli et al, 2013). Studies defined Tscm with several markers including homing marker CD62L and CCR7, stem cell-like associated marker CD127, and memory-associated markers CD45RA, CD45RO, CD27, CD28, CD95, and CD122. Since the limitation of the eight-color FACS instrument used in this study, we chose CCR7, CD45RA, CD27, and CD95 for Tscm identification according to previous study (Ribeiro et al, 2014).
Consistent with the results found in HIV-1 (Ribeiro et al, 2014), we observed that the frequency of CD8+ Tscm subset was positively correlated with CD8+ Tcm and negatively correlated with CD8+ Tem in both HIV+ and HCV+ groups. Interestingly, the HIV/HCV group also had a similar trend (Fig. 2), indicating that the differentiation of CD8+ Tscm could be a key factor to maintain the homeostasis of memory T cell development in HIV, HCV, or HIV/HCV coinfection.
To date, the dynamics of CD8+ Tscm during HCV infection and the role of CD8+ Tscm on the control of HCV infection are not well analyzed. In HIV-1-infected individuals, the proportion and number of HIV-1-specific CD8+ Tscm were associated with number of CD4+ T cells and HIV-1 viral load on ART (Vigano et al, 2015). Low frequency of CD8+ Tscm and the lack of polyfunctional CD8+ Tscm were associated with severe disease progression in chronic chagasic patients (Mateus et al, 2015). CD19-CAR-modified CD8+ Tscm strategy showed enhanced polyfunctionality and long-lasting antitumor responses against human B cell malignancies (Sabatino et al, 2016).
These reports provide the evidence that CD8+ Tscm has the function of antiviral and antitumor activities. Thus, in our data, the negative correlation between CD8+ Tscm and HCV viral load observed in HCV-infected individuals suggests that CD8+ Tscm may have suppressive capacity against HCV replication (Fig. 3).
It is well known that HIV-1 and HCV infection induce T cell immune activation due to persistent viral exposure (Emmanuel et al, 2019; Huang et al, 2022; Vinikoor et al, 2013; Xia et al, 2018). We observed a negative correlation between the proportion of CD8+ Tscm and the level of CD8+ T cell immune activation in HCV infection (Fig. 4C), as well as a positive correlation trend between HCV viral load and the level of immune activation in HCV infection (Fig. 4B). Thus, we hypothesized that CD8+ Tscm inhibits T cell immune activation through controlling HCV replication.
Of note, similar association between CD8+ Tscm and HCV viral load or the immune activation was not found in HIV/HCV-coinfected patients, probably due to the influence of the administration of ART on HIV-1 treatment and immune suppression induced by HIV-1. Furthermore, the immune activation was not decreased to normal level in HIV+ patients, likely caused by residual virus replication despite ongoing ART (Lederman et al, 2013); these factors may impair the level of CD8+ Tscm. ALT and AST are two key indicators related to liver injury, but we observed a negative relationship trend between CD8+ Tscm and these two indicators. Whether these have any implications is unknown and should be verified due to insufficient patients available in our study.
This study had several limitations. First, the sample size is relatively small, thus additional patients should be recruited to confirm our results. Second, we did not analyze the relationship between CD8+ Tscm and the degree of liver fibrosis or cirrhosis, which is an important disease progressive parameter in HCV infection due to the lack of available documentation. Third, due to insufficient patient cells, we could only perform flow cytometry for the correlation analysis result between CD8+ Tscm and HCV viral load from this observational study, which cannot demonstrate directly the role of CD8+ Tscm in the control of HCV replication. The mechanism of HCV-specific CD8+ Tscm to HCV control should be further studied with new recruited patients and cell samples in the future.
Conclusions
In summary, our study identified the distribution of CD8+ Tscm in HCV infection. In this study we report that the elevated CD8+ Tscm proportion was associated with the differentiation of other CD8+ T memory subsets, as well as the level of HCV replication and immune activation in HCV infection. Considering the unique features of Tscm, our findings may have implications for CD8+ Tscm as a potent target for T cell-based vaccine design and immunotherapy for HCV cure.
Footnotes
Acknowledgments
We thank Wei Xia and Yali Liu for recruiting patients and blood sample and information collection and Yunxia Ji, Rui Wang, and Yanxiang Huang for cell counting and HIV or HCV viral load detection.
Authors' Contributions
X.L., B.B.S., T.Z., and Y.G. conceived the study; X.L., B.B.S., and W.W. performed experiments; X.L. and B.B.S. analyzed data; X.L., B.B.S., and A.K.L.C. wrote the article; and B.S., H.W., T.Z., and Y.G. revised the article. All authors read and approved the final article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by the Beijing Municipal Administration of Hospitals' Program (QML20181702 to X.L., DFL20191701 to T.Z.), National Natural Science Foundation of China (NSFC, 81501732 to X.L., 82072271 to T.Z.), Beijing Municipal Education Commission Science and Technology Project (KM202010025015 to X.L.), and Beijing Key Laboratory for HIV/AIDS Research (BZ0089). The funders had no role in study design, data collection and analysis, decision to publish, or in preparation of the article.