
Abstract
Infection caused by the Zika virus (ZIKV) can lead to serious neurological complications such as microcephaly in neonates. At present, no approved ZIKV vaccine is available, but few vaccine candidates are undergoing clinical trial. One major challenge faced is antibody-dependent enhancement (ADE) reaction that may provoke severe outcome in subsequent infection by ZIKV or other flaviviruses. Thus, more efforts should be dedicated to understanding ADE in designing a safe and effective vaccine to minimize the consequence of the potentially fatal infection's complications and to tackle potential ZIKV reemergence. This review discusses different types of ZIKV vaccine candidates that are currently underway in various stages of preclinical and clinical evaluations.
Introduction
Zika virus (ZIKV) is a single-stranded RNA arbovirus belonging to Flavivirus genus, which comprises various important human pathogens, including dengue virus (DENV), yellow fever virus (YFV), and Japanese encephalitis virus (JEV). ZIKV was first discovered from the Zika forest in Uganda in 1947 from the blood sample of a sentinel rhesus monkey and later from the suspension of Aedes africanus mosquito from the same forest (Dick et al, 1952), and subsequently isolated from A. africanus and A. luteocephalus in Africa (Lee and Moore, 1972) and A. aegypti in Malaysia (Marchette et al, 1969). ZIKV was initially believed to cause sporadic infection (MacNamara, 1954; Olson et al, 1981), and associated with mild symptoms in few outbreaks in Yap Island (2007) (Duffy et al, 2009), French Polynesia (2013) (Cao-Lormeau et al, 2014), Cooks Island, and Easter Island (2014) (Tognarelli et al, 2016).
Epidemic in year 2015 has placed ZIKV as one of the most feared pathogen, due to its rapid worldwide transmission in over 80 countries and its connection with neurological deficit cases reported among neonates (Campos et al, 2015), and Guillain-Barré syndrome in adults (Cao-Lormeau et al, 2016).
ZIKV enters human neurons and glial cells through binding to AXL, a member of the Tyro3-Axl-Mer receptor family member (Meertens et al, 2017; Nowakowski et al, 2016), and induces cell death (Gabriel et al, 2017; Tang et al, 2016). AXL null mice (Ifnar−/−/Axl−/−) demonstrates reduced degree of viral pathology upon infection due to reduced viral entry (Hastings et al, 2019). Besides AXL, ZIKV may enter cells through alternate portals as the inhibition of AXL alone is insufficient to prevent ZIKV infection (Wells et al, 2016). After entering cells, ZIKV C protein promotes ribosomal stress by disrupting tumor protein p53 and ribosomal protein L11 (Slomnicki et al, 2017). Besides, it alters subcellular localization of phosphorylated TANK-binding kinase 1 from centromeres to mitochondria, leading to impaired mitosis, supernumerary centrosomes, and aneuploidy (Onorati et al, 2016). This leads to cell death through activation of caspase pathway, as demonstrated by increased cleave caspase 3 expression following intracranial injection of ZIKV in early postnatal mice (Huang et al, 2016).
Furthermore, ZIKV infection in microglial cells promotes inflammatory responses through induction of various proinflammatory cytokines, including interleukin 6, tumor necrosis factor alpha, interleukin 1β, and monocyte chemotactic protein 1, causing neuroinflammation (Lum et al, 2017). Besides neuronal cells, many other cell types are permissive to ZIKV infection, including immune cells such as monocytes, macrophages, mast cells, and other fibroblasts and epithelial cells (Hamel et al, 2015; Ramos da Silva et al, 2019). Fc gamma receptor (FcγR)-bearing immune cells such as monocytes and macrophages serve as important intermediates that facilitate virus replication and spreading through a phenomenon called antibody-dependent enhancement (ADE). For instance, ZIKV infection in placental macrophages or Hofbauer cell results in viral replication and vertical transmission from mother to infant (Schwartz, 2017; Simoni et al, 2017).
The World Health Organization had in 2017 provided a target product profile, which lays out a list of preferred and minimal features for a ZIKV vaccine that is suitable for deployment in times of emergency and outbreak [52]. Numerous ZIKV vaccine platforms are currently in clinical development and preclinical assessment. Of these, a few of them are now at various phases of clinical trials.
DNA Vaccines
GLS-5700 (NCT02809443), a ZIKV DNA vaccine that is developed by Inovio Pharmaceuticals and GeneOne Life Science, had shown favorable results with well-tolerated safety and promising immunogenicity profile in clinical evaluations (Tebas et al, 2017). GLS-5700 possesses both sequences of precursor membrane (prM) and Envelope (Env) and hence expresses a consensus ZIKV-prM and E antigen. Immunization of GLS-5700 in mice and rhesus macaques induces enduring antigen-specific immunoglobulin G (IgG) and T cell responses, and complete protection against ZIKV-associated pathologies upon subsequent viral challenge. Furthermore, adoptive transfer of immune sera from immunized rhesus macaques to Ifnar1−/− mice conferred protection against ZIKV infection (Muthumani et al, 2016). GLS-5700 has been developed soon after ZIKV outbreak in the year 2015, and has advanced into clinical trial in year 2017. In an open-label study phase 1 clinical trial (ZIKA-001), GLS-5700 were delivered into patients through intradermal injection and electroporation at three different intervals within 12 weeks.
In general, all patients developed binding antibodies and neutralizing antibodies were detected in 62% of subjects utilizing Vero-cell assay. Neuronal-cell neutralization assay showed that 70% of the serum samples exhibit 90% inhibition of ZIKV infection and 95% showed 50% inhibition. Additionally, in comparison with the control mice, 92% of the mice that received sera from postvaccination patients through the intraperitoneal route are protected against infection with a lethal dose of ZIKV-PR209 strain (Tebas et al, 2017).
In the year 2016, the Vaccine Research Center at the National Institute of Allergy and Infectious Diseases (NIAID) constructed two DNA vaccines that are derivations of the French Polynesian isolate (strain H/PF/2013) (Dowd et al, 2016). Both vaccines, codenamed VRC5283 and VRC5288, have their prM signal sequence and part of the stem and transmembrane regions (ST-TM) replaced with the JEV analogous sequences, respectively. These respective modifications are believed to enhance the expression and secretion of subviral particles of VRC5283 and VRC5288. VRC5288 and VRC5283 are nearly identical, except for their E regions: VRC5288 has a chimeric E region that contains of a mixture of ZIKV sequences at the extracellular region and JEV sequences at the stem and transmembrane region, whereas VRC5283 E region is derived solely from ZIKV sequences (Dowd et al, 2016).
Both vaccines elicit strong sera neutralizing activities in mice and nonhuman primates (NHP) post-ZIKV challenge. Additionally, viremia was not detectable in 17 of 18 (94%) of nonhuman primates that were administered with two doses of vaccines (Dowd et al, 2016).
Subsequent clinical evaluations of VRC5283 and VRC5288 show high positive antibody responses and significant CD4+ and CD8+ T cell responses. VRC5283 elicits higher immunogenicity profiles than VRC5288 and has thus progressed to assessment in phase 2 of clinical trial (NCT03110770) (Gaudinski et al, 2018).
Grubor-Bauk and colleagues have synthesized three ZIKV DNA vaccines (pVAX-NS1, pVAX-tpaNS1, and pVAX-tpaNS1-IMX313P) that, in comparison with other prM and/or E-based DNA vaccines, contain the coding sequences for NS1. pVAX-NS1 encodes wild-type NS1, whereas pVAX-tpaNS1 secretes a fusion NS1 with a tissue plasminogen activator (TPA) leader sequence upstream of NS1 to facilitate secretion, and pVAX-tpaNS1-IMX313P expresses a heptameric NS1 by virtue of the presence of the oligomerization domain from the chicken complement inhibitor C4b-binding protein known as IMX313P, respectively. One of the ZIKV NS1 DNA vaccines, namely pVAX-tpaNS1-IMX313P, when administered intradermally into BALB/c and Ifnar−/− mice, induced robust cellular immunity characterized by enhanced CD4+ and CD8+ responses and protected mice from intravenous challenge with ZIKV.
The authors also noted that although increased humoral response was detected in mice following vaccination, the resultant anti-NS1 antibodies were not able to provide protection against viral replication in the absence of CD4+ and CD8+ responses (Grubor-Bauk et al, 2019). These results lend support to the hypothesis that immunodominant epitopes on nonstructural proteins and cellular immunity are key to elicit a protective anti-ZIKV immunity, as has been argued as the reason for the poor performance of Dengvaxia and underscore NS1 as an important target in the design and production of ZIKV vaccines.
RNA Vaccines
Several mRNA-based vaccine candidates have shown encouraging results against ZIKV. In contrast to other vaccine platforms, mRNA can be a safer strategy due to its noninfectious and nonintegrating nature, and thus the infectious risk and potential for mutation arising from insertion can be negated. Furthermore, mRNA can be readily modified to improve translation and stability. This versatility provides a possibility for rapid production and deployment (Pardi et al, 2018).
Richner et al (2017) synthesized a lipid nanoparticle encapsulated nucleoside-modified mRNA (mRNA–LNP) based on an Asian ZIKV strain isolated in 2007. This mRNA possesses a modified prME sequence that carries signal sequence from human IgE or JEV prM, and 5′ and 3′ untranslated regions (UTR) optimized to enhance translation and intracellular stability, as well as modification to the nucleoside to reduce the stimulation of innate immunity (Richner et al, 2017). Pardi et al (2017) designed a similar mRNA vaccine candidate that encodes the codon-optimized prME sequence from the French Polynesian ZIKV strain H/PF/2013 (Pardi et al, 2017). Both vaccines have been used on animals and were protective against ZIKV challenge (Pardi et al, 2017; Richner et al, 2017). Noteworthy, the mRNA-LNP by Richner et al (2017) was able to induce sterilizing immunity in mice and decrease ADE to DENV in cells or mice (Richner et al, 2017).
Other vaccine, mRNA-1325, has completed its clinical studies in 2019 (NCT03014089), but information on the preclinical and clinical efficacy are not available. Jagger et al (2019) employed the same method as that of Richner et al (2017) to generate an mRNA vaccine that was fully protective against vertical transmission in STAT2-knockin transgenic immunocompetent pregnant mice (Jagger et al, 2019). This vaccine, mRNA-1893, has recently entered into phase 1 clinical trial (NCT04064905).
Inactivated Viral Vaccines
In the year 2016, the Walter Reed Army Institute of Research (WRAIR) had developed a purified formalin-inactivated Zika virus vaccine (ZPIV) that was derived from the Puerto Rican ZIKV isolate (PRVABC59) (Larocca et al, 2016). Preclinical investigations of ZPIV using mouse and NHP models showed full protection to animals against challenges by both Puerto Rican and Brazilian ZIKV strains (Abbink et al, 2016; Larocca et al, 2016). ZPIV was assessed in three phase 1 clinical trials (NCT02963909, NCT02952833, and NCT02937233) as reported by Modjarrad et al (2018), and subjects were vaccinated with two doses of ZPIV adjuvanted with aluminum hydroxide between 4 weeks interval.
In brief, seroconversion occurred in 92% of participants with microneutralization titer ≥1:10. Infusion of purified IgG from vaccinated subjects into virus-naive mice resulted in partial to full prevention of viremia as compared with mice that were passively immunized with purified IgG from an individual without vaccination or placebo participants (Modjarrad et al, 2018). An optimized version of the vaccine ZPIV-SP, jointly developed by Sanofi Pasteur and WRAIR, exhibited 100% seroconversion rate and overall better immunogenicity than ZPIV in mice studies.
However, the efficacy of this vaccine in nonhuman primates and humans remains to be elucidated (Lecouturier et al, 2019). Similar vaccine platform has been adopted by Bharat Biotech (MR8766), Valneva Austria GmbH together with Emergent BioSolutions (VLA-1601), as well as Takeda Vaccines (TAK-426), and each of this vaccine candidate has entered into phase 1 clinical trial (CTRI/2017/05/008539, NCT03425149, NCT03343626, respectively). Preclinical investigations involving MR8766 and TAK-426 have shown promising capacity for inducing protective responses in vitro and in animals (Baldwin et al, 2018; Sumathy et al, 2017).
Live-Attenuated Viral Vaccines
Live-attenuated vaccine represents another platform, apart from mRNA-LNP, with which sterilizing immunity could be achieved. Shan et al created a live-attenuated vaccine bearing 10-nuleotide deletion in the 3′-UTR in the genome of the ZIKV strain Cambodian FSS13025 (10-del ZIKV) that conferred sterilizing immunity with a single dose in mice studies (Shan et al, 2017b). Additional investigation of a derivative of this vaccine carrying 20-nucleotide deletions in the 3′-UTR in mice demonstrated its efficacy in protecting mice against vertical transmission and testis damage as well preventing viremia in NHPs (Shan et al, 2017a).
Xie and colleagues used a similar approach to render ZIKV strain Cambodian FSS13025 defective in multiplication, resulting from deletion of 9 amino acids at the C-terminal of viral capsid. Application of this vaccine candidate on A129 mice completely abrogated viremia and infection-associated morbidity and mortality. Prevention of vertical transmission in pregnant mice was also noted after a single-dose vaccination (Xie et al, 2018). A chimeric live-attenuated ZIKV containing prME with sequence contribution from both ZIKV prME and DENV-2 has been described (Xie et al, 2017).
This vaccine candidate demonstrated high immunogenic capacity and provided protection against ZIKV and DENV challenge in mice (Xie et al, 2017). NIAID was involved in the development of a chimeric vaccine rZIKV/D4Δ30-713, which has had 30 nucleotides within its DENV-4 sequences deleted. This vaccine has advanced to clinical evaluation (NCT03611946) since year 2018, but reports of its efficacy is yet to be published.
A novel DNA-launched live-attenuated vaccine approach was elaborated by Zou et al (2018), which combines the features of DNA-based and live-attenuated vaccine (Zou et al, 2018). This method involves synthesis of a recombinant plasmid that carries the cDNA sequence of ZIKV strain Cambodian FSS13025 that has been attenuated through deletion of 20 nucleotides in the 3′-UTR. In this manner, production of live-attenuated virus takes place within cells, and protective immunity could thus be produced. When administered into mice, this vaccine imparted full protection to ZIKV disease and maternal-to-fetal transmission. In addition, neutralizing antibodies developed in fetal serum and shielded male mice from testis infection, injury, as well as oligospermia (Zou et al, 2018).
Recombinant Vaccines
Various recombinant viruses are under preclinical and clinical evaluations for their potential as viral vector vaccine. One prominent feature of viral vector vaccine lies in its ability to induce robust humoral immunity, and thus enduring protection can be sustained without multiple booster administrations (Lauer et al, 2017). Adenovirus has been the most common vector used in the construction of ZIKV viral vector vaccine. One recombinant rhesus adenovirus serotype 52 vector expressing ZIKV prM-Env (RhAd52-M-Env) had shown neutralizing antibodies inducing capacity and afforded full protection to ZIKV challenge in NHPs with a single administration (Abbink et al, 2016).
Continuing investigation of this vaccine in NHPs revealed sustained protection against ZIKV challenge 1 year postvaccination (Abbink et al, 2017). Adenoviral and vesicular stomatitis virus-based vectors have been constructed by other groups and showed potent immunogenicity and potential for protection against viremia and ZIKV-associated diseases in mice (Cox et al, 2018; Guo et al, 2018; Lopez-Camacho et al, 2018; Xu et al, 2018). A vaccinia-based viral vector with potency against ZIKV and Chikungunya (CHKV) was described by Prow et al (2018).
This vector was derived from the Sementis Copenhagen Vector (SCV) vaccine technology generated from deletion of the viral assembly protein gene D13L to nullify the infectivity of the vector (Eldi et al, 2017). Both ZIKV prME and CHKV structural protein C-E3-E2-6K-E1 were later incorporated into SCV and the resulting SCV-ZIKA/CHIK could protect against ZKV and CHKV viremia, ZIKV fetal placental transmission, CHKV-associated arthritis, as well as testes infection and pathology in mice (Prow et al, 2018). Themis Bioscience GmbH created a measles virus (MV)-based viral vector from the MV Schwarz vaccine strain (MVSchw) encoding ZIKV prME and tested this vaccine in a phase 1 clinical trial in Austria (NCT02996890) in the year 2016. This vaccine was evaluated in preclinical studies involving allogeneic pregnancy mice and demonstrated its capability to reduce ZIKV viral load in different organs and placenta, as well as the ability to protect fetal mice from ZIKV infection (Nurnberger et al, 2019). Themis Bioscience GmbH initiated another phase 1 clinical trial for another of its MV-based viral vector (MV-ZIKA-RSP) in 2019.
Subunit Vaccines
Virus-like particles (VLPs) comprised snippets of the virus in the form of structural proteins that can self-assemble to form a noninfectious particle that is devoid of the viral genome (Donaldson et al, 2018). A number of groups have developed VLPs for ZIKV infection and validated their immunogenic potentials in mice (Boigard et al, 2017; Espinosa et al, 2018; Garg et al, 2020; Garg et al, 2019; Garg et al, 2017). One study showed that immunization of mice with VLPs generated higher titers of neutralizing antibodies in comparison with DNA vaccines (Garg et al, 2017). Recently, Garg et al (2020) has successfully engineered a tetravalent vaccine that expresses the structural and capsid proteins of ZIKV, JEV, and West Nile virus (WNV) from Flavivirus genus of Flaviviridae family, and CHIKV from Alphavirus genus of Togaviridae family, with strong capability to induce neutralizing antibodies upon immunization of Balb/c mice (Garg et al, 2020).
ZIKV envelope (E) protein is the major target of neutralizing antibodies. Truncated N-terminal E subunit or EDIII domain has been shown to be immunogenic and protective against ZIKV infection when administered in the experimental animals (Amaral et al, 2020; Han et al, 2017; Liang et al, 2018; Yang et al, 2017). However, E protein contains a highly conserved fusion loop (FL) domain that is known to be the main source of cross-reactive antibodies for Flaviviruses that causes ADE. Many studies are ongoing to engineer the E subunit vaccine by disrupting FL epitope conformation but retaining the neutralizing epitope, to diminish the enhancement effect (Berneck et al, 2020; Dai et al, 2021). An association of subunit vaccine and ADE is as discussed below.
ADE in ZIKV Pathogenesis
ADE has been documented for many different viruses, such as coxsackievirus B, Ebola virus, influenza A virus, respiratory syncytial virus, coronavirus, human immunodeficiency virus, and flaviviruses, including DENV, YFV, JEV, WNV, Wesselsbron virus, and tick-borne encephalitis virus (Fagbami and Halstead, 1986; Gould and Buckley, 1989; Katzelnick et al, 2017; Kuzmina et al, 2018; Phillpotts et al, 1985; Sauter and Hober, 2009; Swanstrom et al, 2016; Tamura et al, 1994; Wan et al, 2020; Willey et al, 2011). Members of the genus Flavivirus demonstrate a significant degree of protein sequence homology. DENV for example, shares 55.6% of amino acid sequence identity with ZIKV, and high level of sequence similarities have been reported between ZIKV and WNV (56.1%), as well as JEV (57.0%) (Chang et al, 2017).
Thus, antigenic cross-reactivity is a common occurrence among the members of Flavivirus (Calisher et al, 1989; De Madrid and Porterfield, 1974). Given the close phylogenetic relatedness between ZIKV and other flaviviruses (Kuno et al, 1998), ADE carries important implication for disease pathogenesis as it may portend the onset of severe diseases.
ADE arises due to suboptimal antibody responses against viral infections, which paradoxically facilitates viral invasion, thus amplifying viral replication and leading to exacerbation of diseases (Tirado and Yoon, 2003). A widely accepted model of the underlying mechanism of this phenomenon involves the interaction of fragment crystallizable region (Fc region) of the virus–antibody complex with the Fc-receptor (mainly FcγR)-bearing cells of the immune system, such as macrophages and monocytes (Fig. 1). In this scenario, the FcγR-bearing cells recognize and attach to the Fc-portion of the subneutralizing or non-neutralizing antibodies bound to viral particles, resulting in enhanced virus uptake and titer in the cells (Flipse et al, 2013).
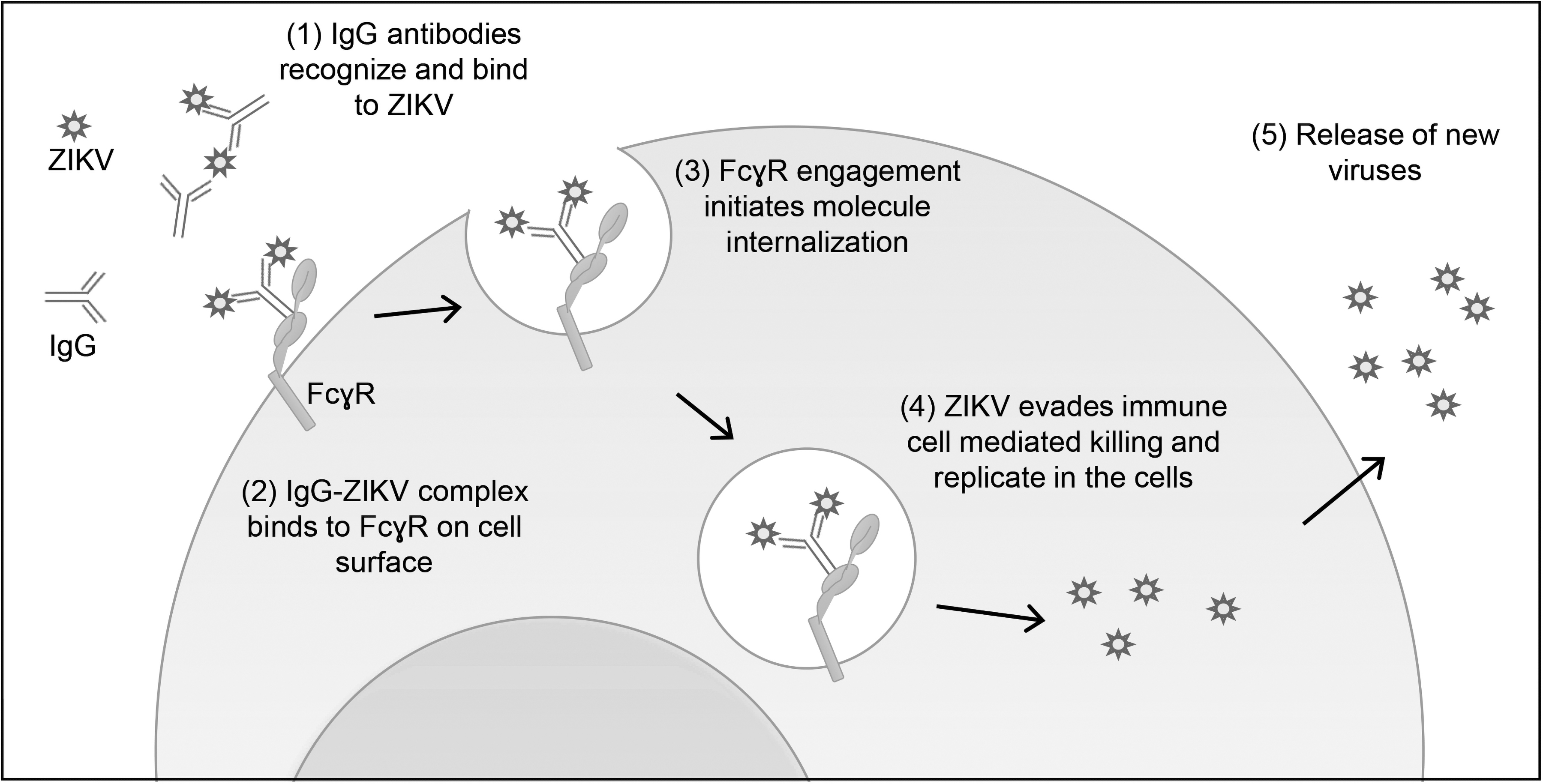
An illustration of ADE and its implication for ZIKV pathogenesis. ADE occurs when suboptimal antibody responses fail to eliminate viral infections, and result in amplification of viral replication. In this phenomenon, antibody first binds to ZIKV but unable to fully neutralize the virus. The antibody–virus complex binds to FcγR-bearing cells of the immune system such as macrophages and monocytes. FcγR engagement initiates internalization of the antibody–virus complex. Virus immune evasion strategy enables its survival and replication in the cells, leading to generation of new viruses, which exacerbates the disease. ADE, antibody-dependent enhancement; FcγR, fragment crystallizable region gamma receptor; ZIKV, Zika virus.
Majority of in vitro experiments employing FcγR-bearing cell lines have found a positive relation between pre-existing antiflavivirus immunity, especially that of DENV, with enhanced ZIKV infection (Bardina et al, 2017; Castanha et al, 2017; Charles and Christofferson, 2016; Dejnirattisai et al, 2016; Kam et al, 2017; McCracken et al, 2017; Pantoja et al, 2017; Paul et al, 2016; Priyamvada et al, 2016; Stettler et al, 2016; Swanstrom et al, 2016). WNV-convalescent plasma also strongly enhanced ZIKV infection (Bardina et al, 2017). FcγR-blocking antibody reverses the placenta damage caused by ZIKV infection, suggesting that the ADE is critical in exaggerating the disease outcome. Although several instances of cross-neutralizing activity of DENV antibodies against ZIKV infection were noted in a few investigations (Kam et al, 2017; Swanstrom et al, 2016), in most cases, cross-reactive antibodies were associated with significant enhancement of ZIKV infection (Bardina et al, 2017; Castanha et al, 2017; Charles and Christofferson, 2016; Dejnirattisai et al, 2016; Paul et al, 2016; Priyamvada et al, 2016; Stettler et al, 2016; Swanstrom et al, 2016).
Recently, a fatal case of DENV infection has been attributed to ADE effect provoked by anti-ZIKV antibodies generated from a prior ZIKV infection (Bonheur et al, 2021). It is worrisome as both tetravalent live-attenuated DENV vaccine (Dengvaxia) and a physical tetravalent DENV mixture (TV-DENV) produces antibodies that vigorously promote ADE of ZIKV infection (Shukla et al, 2020). Due to similarity of the DENV and ZIKV, it is plausible that ZIKV vaccine could trigger a similar consequence in the immunized individuals.
Although general recognition exists in the literature concerning the in vitro cross-reactive antibody enhancement of ZIKV infection, in vivo studies using predominantly mouse models and a few involving NHP have yielded mixed results that do not provide empirical support to this observation. Clinical manifestations of worsened disease outcome in the form of fever, viremia, higher viral loads in the spinal cord and testes, as well as elevated mortality were detected in infection studies using Stat2−/− mice that were administered with DENV- or WNV-convalescent plasma (Bardina et al, 2017). Cross-protection to ZIKV was seen previously in mice that received high concentration of DENV-immune plasma with correlated increase in survival and reduced clinical symptoms, and similar dose-dependent response was also reported in mice pretreated with WNV-immune plasma. This suggests that antibody dose may play a role in shaping the overall response during heterologous and homologous flavivirus infection (McCracken et al, 2017).
On the contrary, studies using NHP models have been unable to reproduce the results obtained with murine models. Enhancement of disease was not observed in one study of rhesus macaques using two cohorts consisting of animals that were 2.8 years post-DENV infection (Pantoja et al, 2017). Similarly, a separate investigation in which rhesus macaques with previous infections (≥420 days) of DENV2, DENV4, or YFV demonstrated no significant increase in viral titers, levels of neutralizing antibody, as well as profile of cell-mediated immunity as compared with flavivirus-native control group (McCracken et al, 2017).
Shim et al (2019) has produced early evidence for homotypic enhancement of ZIKV infection (Shim et al, 2019). It was reported that inoculation of ZIKV-immune plasma to Ifnar1−/−-immunocompromised mice provides homotypic protection to the animals against ZIKV reinfection in a dose-dependent manner, where low concentration of infused plasma induced ADE of infection characterized by increased mortality. On the other hand, significant improvement in survival rate was observed when mice received higher concentration of plasma. In addition, infusion of ZIKV-immune plasma into Ifnar1−/− pregnant mice reduced the body weight of fetal mice and increased fetal demise (Shim et al, 2019), suggesting a possible link between homotypic ZIKV ADE of infection with severe clinical course, at least in adult and pregnant mice.
Viral Elements Associated with ADE
Flaviviruses contain a single polyprotein-encoding RNA genome which, following translation and processing by the viral and cellular proteases, produces three structural proteins (capsid protein [C], envelope protein [E], and precursor membrane protein [M]), as well as seven nonstructural proteins (NS1, NS2A, NS2B, NS3, NS4A, NS4B, and NS5) (Chin et al, 2007; Dai et al, 2016; Luca et al, 2012). The viral genome is enclosed within a capsid. The E and M proteins are embedded into a bilayer lipid membrane to form an icosahedral outer shell that encircles the nucleocapsid (Therkelsen et al, 2018). Structurally conserved across flaviviruses, the E protein is involved in viral invasion by mediating attachment to host cell as well as subsequent virus–host membrane fusion (Modis et al, 2004). The E protein comprises three domains (domain DI, DII, and DIII) (Modis et al, 2004), which are the major antigenic sites that give rise to neutralizing antibodies with different breadth and potency.
Potently neutralizing antibodies primarily target epitopes found on DI, II, III, as well as other confirmational epitopes on the quaternary structure of E proteins. Antibodies to DIII largely exhibit high neutralizing capacity against WNV, DENV, and ZIKV and are associated with reduced Flavivirus pathogenesis as well as viral-induced mortality in mice (Beasley and Barrett, 2002; Gromowski and Barrett, 2007; Oliphant et al, 2005; Sanchez et al, 2005; Shrestha et al, 2010; Stettler et al, 2016). DIII-specific neutralizing antibodies isolated from ZIKV-infected individuals have also been shown to be able to confer cross-humoral protection against ZIKV and/or DENV infections in mice and NHPs (Keeffe et al, 2018; Kim et al, 2021; Magnani et al, 2017; Niu et al, 2019; Robbiani et al, 2017; Zhao et al, 2020). Such antibodies, however, appear to occur less frequently in humans (Gallichotte et al, 2019). Other potent neutralizing antibodies to flaviviruses recognize the complex quaternary epitopes comprising E multimers.
The E-dimers are highly conserved among flaviviruses and play a role in the binding of prM for immature virions during viral assembly and maturation (Rey et al, 2018). Antibodies directed against a region within the E-dimer known as the E-dimer epitope (EDE) produced by DENV-infected patients, for example, are capable of neutralizing all four DENV serotypes and are effective in preventing lethal ZIKV infection, in addition to reducing viral burden as well as fetal death when administered in mice (Barba-Spaeth et al, 2016; Dejnirattisai et al, 2015; Fernandez et al, 2017). DENV-specific antibody against the EDE also demonstrates protective effect toward viral escape in rhesus monkeys, suggesting its resilience against the emergence of immune escape viral mutations (Abbink et al, 2018). Reactive antibodies recognizing the connecting region between DI and DIII (DI/DIII-linker), on the other hand, can also cross-neutralize DENV and ZIKV in vivo (Dussupt et al, 2020), and inhibit ADE in vitro or may promote the development of immune escape variants (Wang et al, 2017).
Poorly neutralizing and infection-augmenting antibodies have been reported to bind predominantly to the prM as well as the fusion-loop epitope (FLE) within DII. DENV infection in humans generates cross-reactive prM-antibodies, which predispose to ADE in a FcγR-dependent manner (Beltramello et al, 2010; Dejnirattisai et al, 2010). Moreover, prM antibodies can interact with immature virions during DENV infection and promote invasion of immature viral particles into cells bearing FcγR, potentially leading to disease pathogenesis (Rodenhuis-Zybert et al, 2010). FLE-specific antibodies derived from DENV- or ZIKV-infected patients tend to be nonprotective and have been observed to enhance ZIKV infections in cell cultures as well as experiments using murine models (Barba-Spaeth et al, 2016; Bardina et al, 2017; Brown et al, 2019; Dejnirattisai et al, 2015; Stettler et al, 2016), but inconsistent results have been obtained from macaque studies (George et al, 2017; McCracken et al, 2017; Pantoja et al, 2017).
More recent epidemiological studies involving two cohorts from Nicaragua and Brazil, on the other hand, showed that preexisting DENV immunity correlates with decreased risk of symptomatic ZIKV and suggested that prior DENV may provide protection to ZIKV infection (Gordon et al, 2019; Rodriguez-Barraquer et al, 2019).
Learning from Field Experience
One crucial goal of ZIKV vaccine is to elicit cross-neutralizing sterilizing immunity with minimum adverse effects such as ADE. However, epitope conservation between ZIKV and other flaviviruses, particularly DENV, represents a key concern for the development of a ZIKV vaccine that is safe for use in humans. As ZIKV cocirculates with DENV and share a common vector, cautionary steps must be taken in the design of a vaccine for ZIKV and DENV and its eventual deployment. The development of a YFV live-attenuated vaccine (YF17D) in the 1930s and its subsequent global roll-out has lent credence to the feasibility of a DENV vaccine (Monath, 2005). Dengvaxia, manufactured by Sanofi Pasteur, is a chimeric multivalent live-attenuated vaccine constructed using the nonstructural genes of YF17D and the prM and E genes of each of the DENV serotypes, and represents the first dengue vaccine that is approved for use in dengue prevention.
Although large-scale clinical studies to examine the efficacy and long-term safety of Dengvaxia involving more than 30,000 children 2–16 years of age 2 years postvaccination in several endemic Asian as well as Latin American countries have demonstrated an efficacy of 60.3% against symptomatic DENV infection across all individuals, the rates were lower in vaccinated children younger than 9 years of age compared with the 9–16-year-old group, at 44.6% and 65.6%, respectively.
Extended efficacy trials in the years that followed using anti-NS1 IgG ELISA as a baseline to differentiate the antibodies between participants previously exposed to DENV and vaccinees showed that Dengvaxia conferred only 39% protection against virologically confirmed dengue in seronegative children (Hadinegoro et al, 2015; Sridhar et al, 2018; Villar et al, 2015). In light of these findings, it has been argued that inadequate T cell responses might be responsible for this lower-than-expected protective efficacy of Dengvaxia, as Dengvaxia does not carry nonstructural proteins belonging to DENV, which previous studies have shown to contain immunodominant epitopes for CD4+ and CD8+ T cells.
In addition, imbalance in immunity resulting from the combination of four different DENV serotypes might have skewed the response in favor of one serotype over another (Halstead, 2012). This likely stems from the competition in the differential replicative ability among different DENV serotypes, and if exist, the presence of immunodominant epitopes may also interfere with the magnitude of cellular immunity that ensues, a suggestion supported by the observation in rhesus monkeys administered with live-attenuated, bivalent and/or tetravalent DENV vaccines (Guy et al, 2009). Concordantly, a subsequent serological analysis in a group of DENV-exposed individuals who were vaccinated with Dengvaxia induces antibodies predominantly toward DENV-4, suggesting the occurrence of interference and that Dengvaxia elicits a response that closely mirrors a monotypic DENV infection (Henein et al, 2017). On the other hand, the finding that children between 2 and 5 years of age showed a higher rates of hospitalization 3 years postvaccination raised a red flag about the possibility of ADE in Dengvaxia recipients (Sridhar et al, 2018).
Recently, Shukla et al (2020) found that antibodies generated by Dengvaxia, while able to cross-neutralize ZIKV, offered no protection to mice against lethal DENV challenge and promoted the development of ADE of both DENV and ZIKV infection.
Conclusions
Despite having made significant strides in developing an efficacious ZIKV vaccine, there still exists a number of challenges and obstacles to be overcome before licensure and eventual deployment of a ZIKV vaccine can be realized. Dwindling number of ZIKV infections in much of the world in recent years has made clinical trials difficult to conduct and this has become a major setback to pushing through the development as well as testing the efficacy of potential ZIKV vaccines. One probable solution to the shortage of human subjects, although highly controversial, is controlled human infection studies. Ethical and legal challenges need to be addressed and importantly, cooperation with relevant authorities on all fronts are required in the drawing as well as execution of a blueprint for such studies. Lastly, as ZIKV and other flaviviruses share the same vector, cotransmission and cocirculation between flaviviruses in a given environment may occur, and thus more efforts should be dedicated to understanding ADE as well as its implication on the safety and efficacy of a potential ZIKV vaccine.
Footnotes
Acknowledgment
The authors thank Dr. Kim Kee Tan for project management.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This project was supported by Malaysia Ministry of Higher Education Long-Term Research Grant Scheme (LRGS MRUN Phase 1: LRGS/MRUN/F1/01/2018) and University of Malaya (GPF004A-2020).